Characterizing the Influence of a Heterotrophic Bicosoecid Flagellate Pseudobodo sp. on the Dinoflagellate Gambierdiscus balechii

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
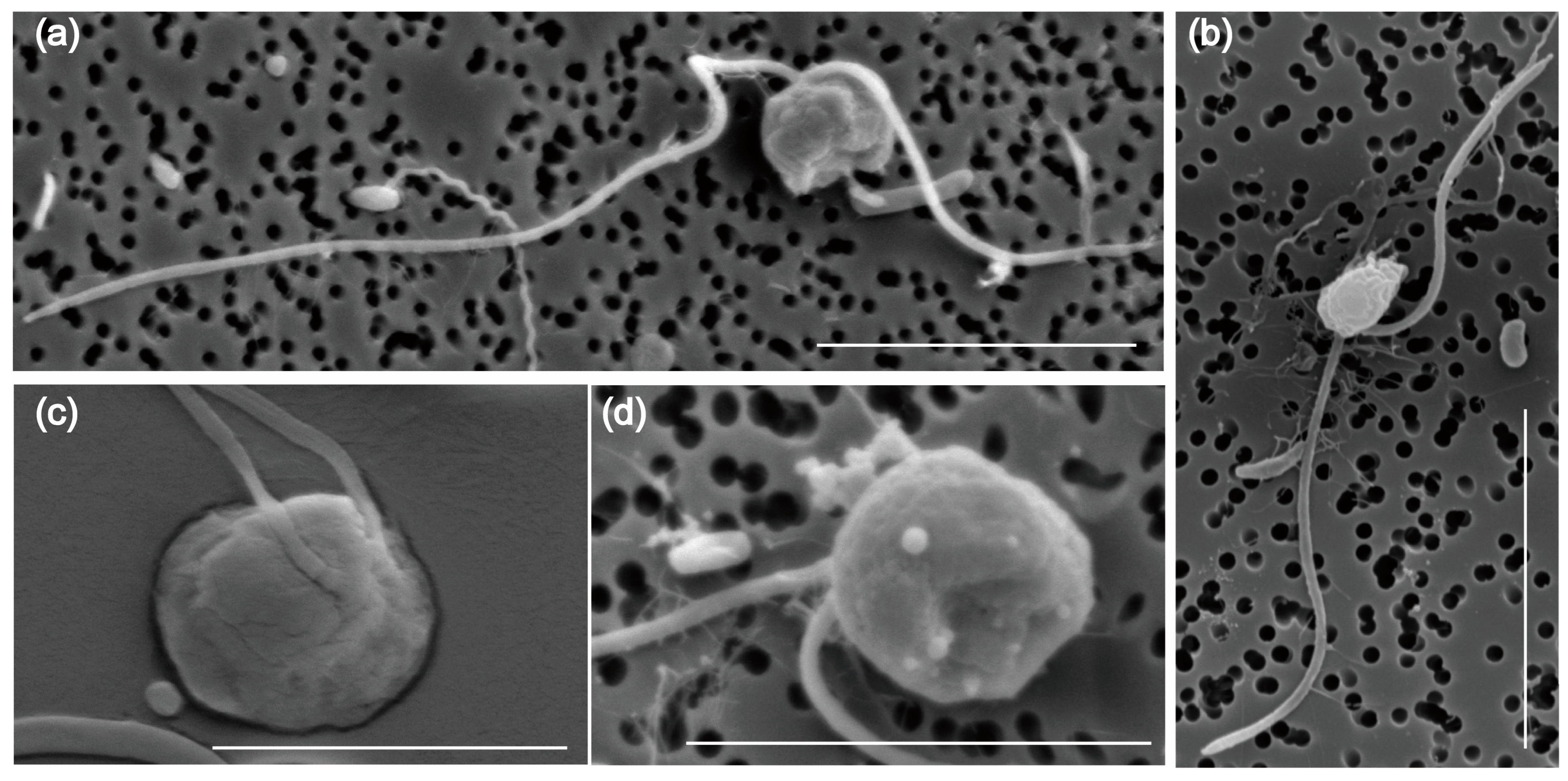
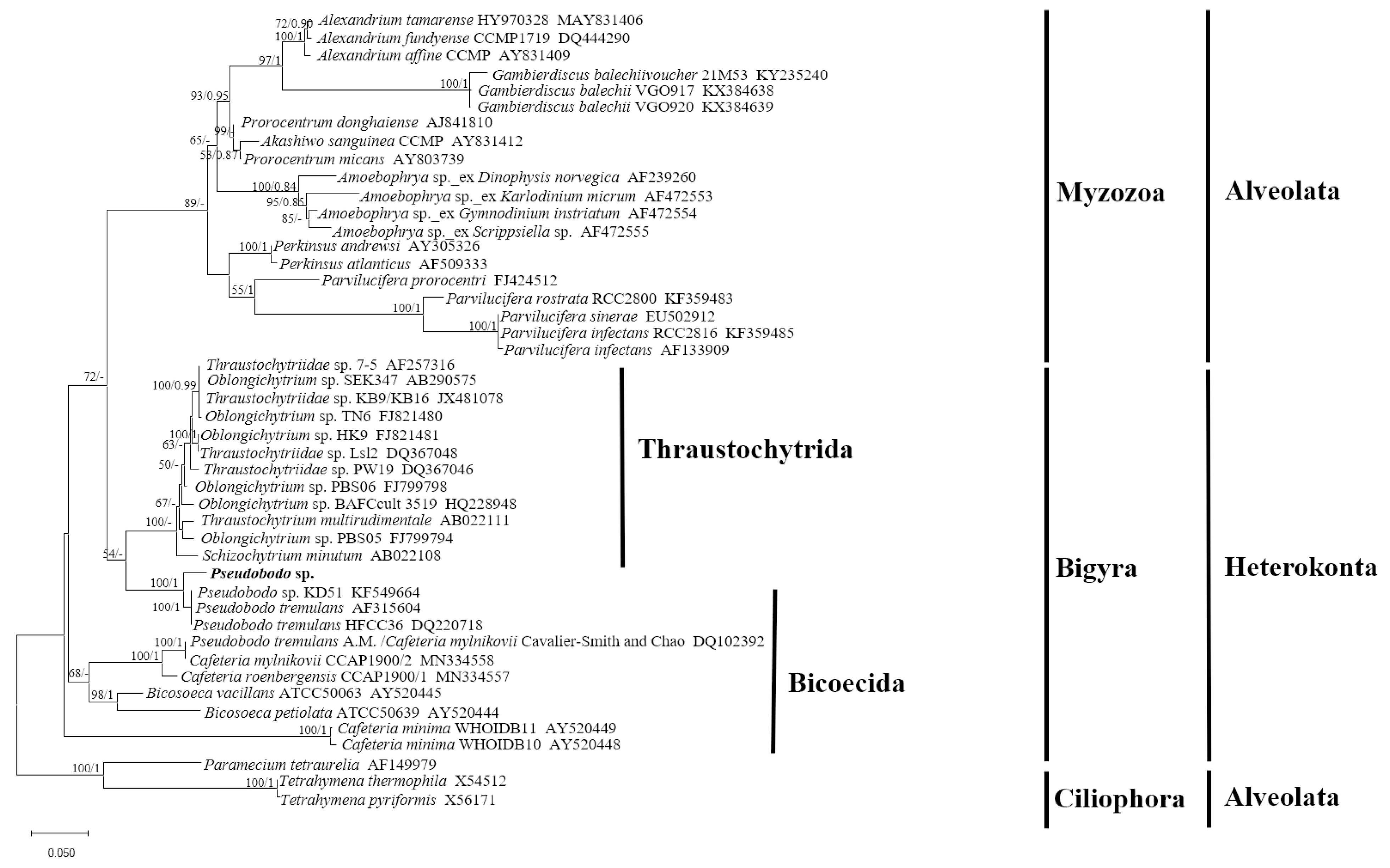
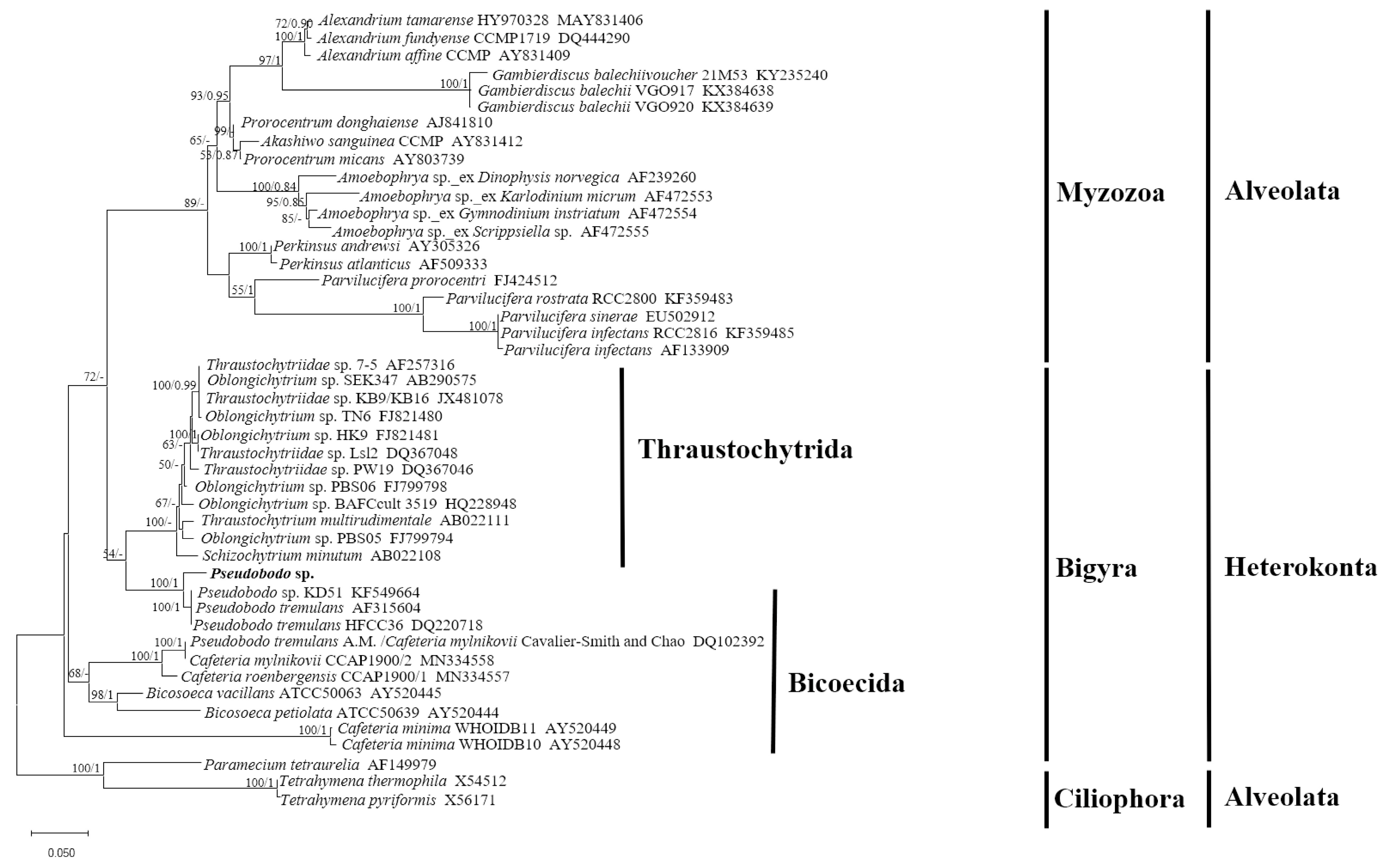
2.1. Morphological Characteristics and Phylogenetic Analysis of Pseudobodo sp.
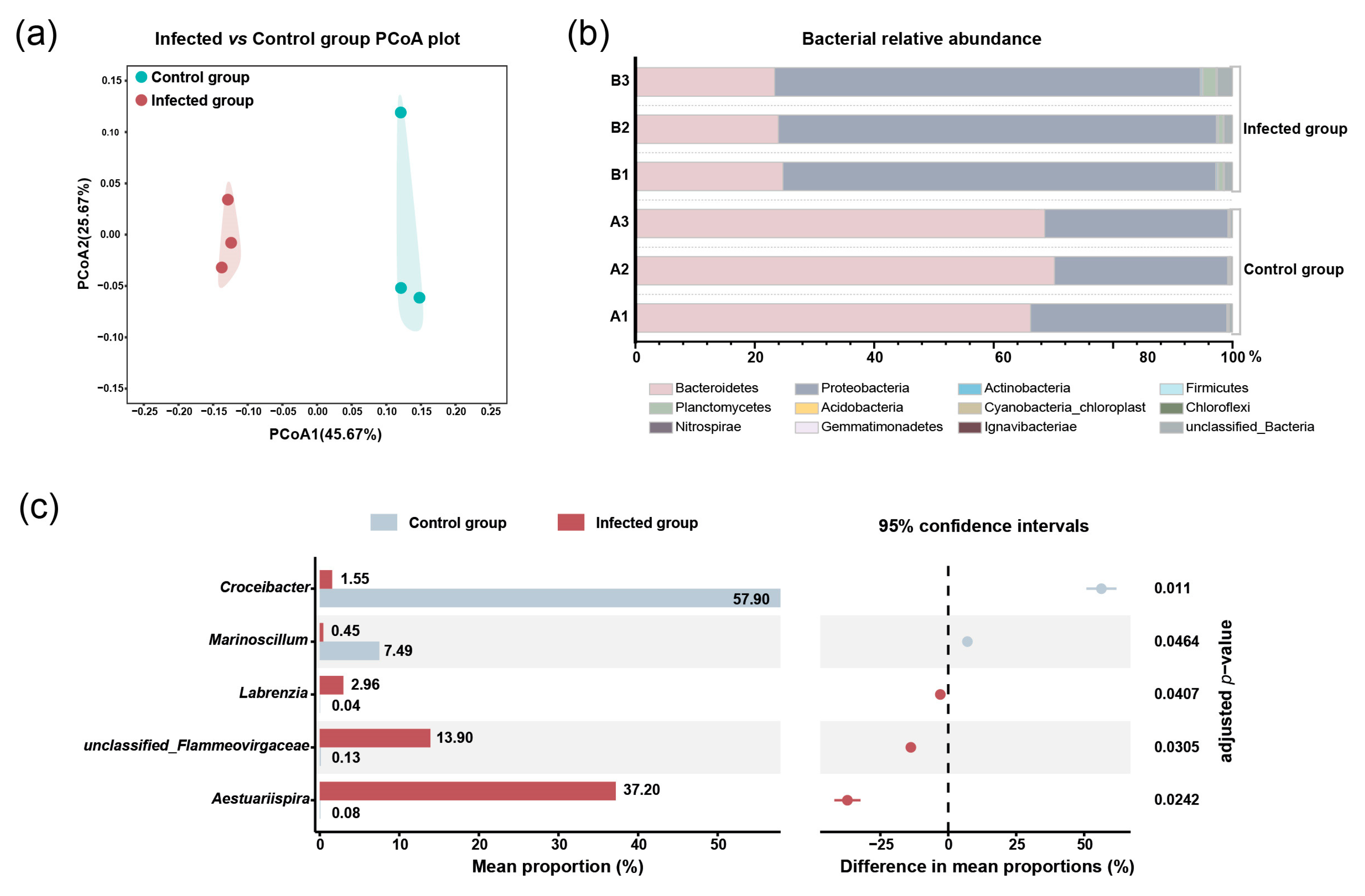
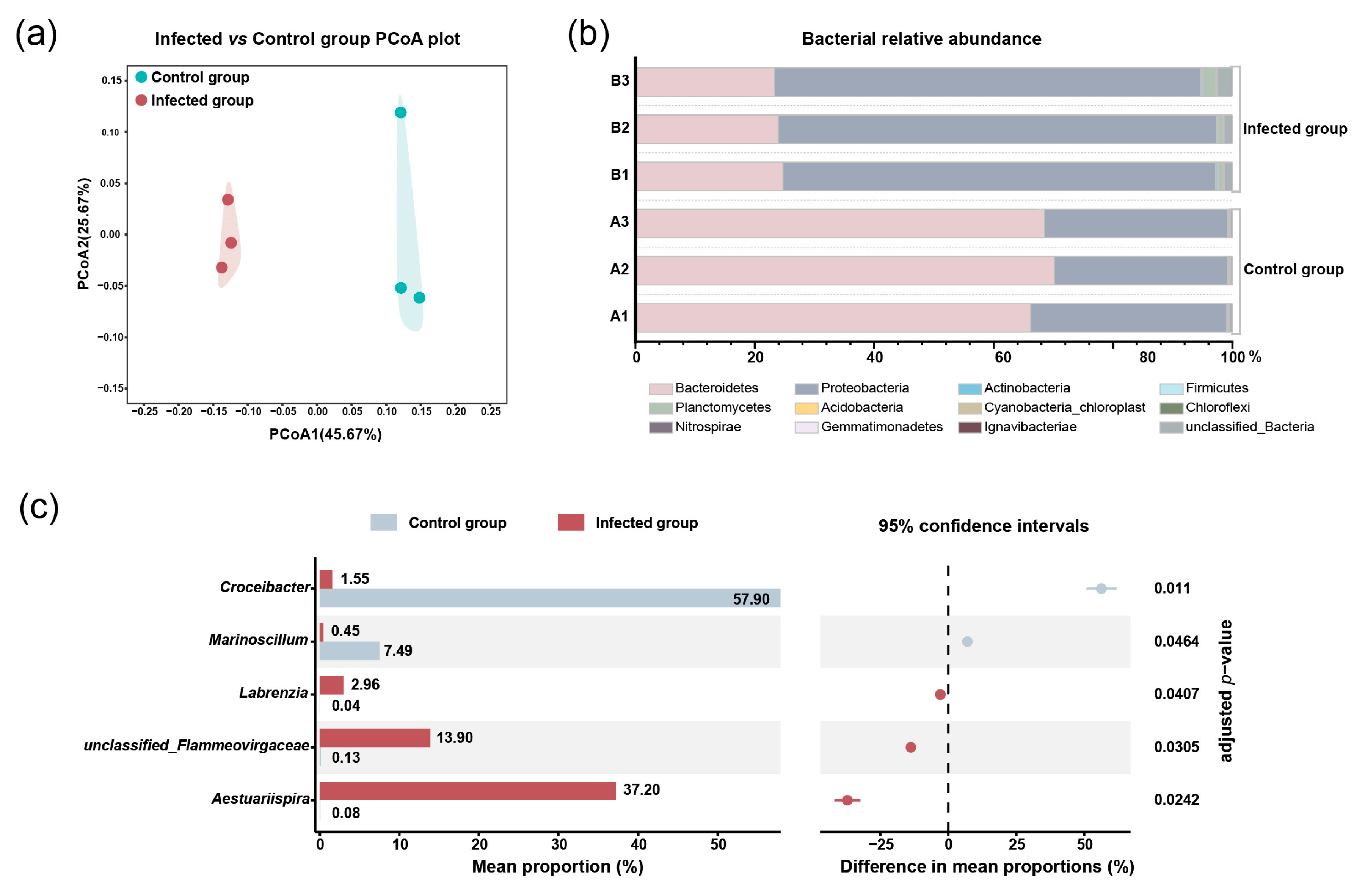
2.2. The Impacts of Pseudobodo sp. on Bacterial Community of Algal Culture
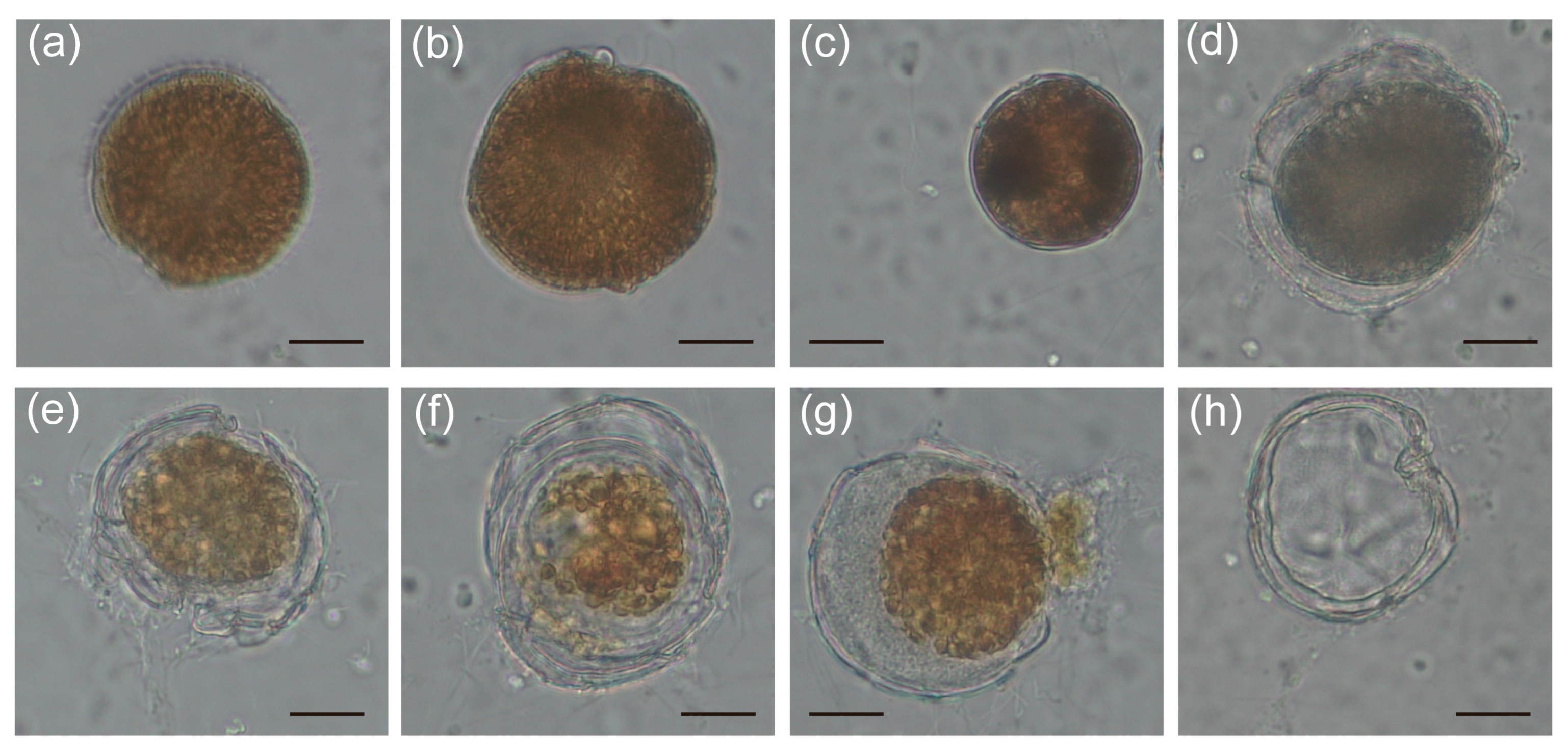
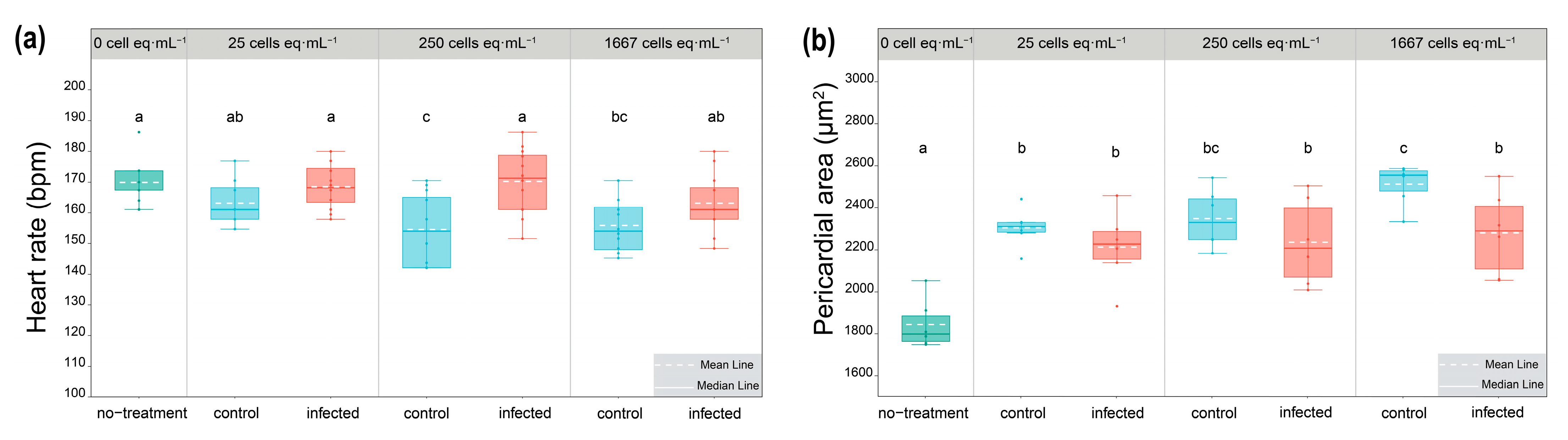
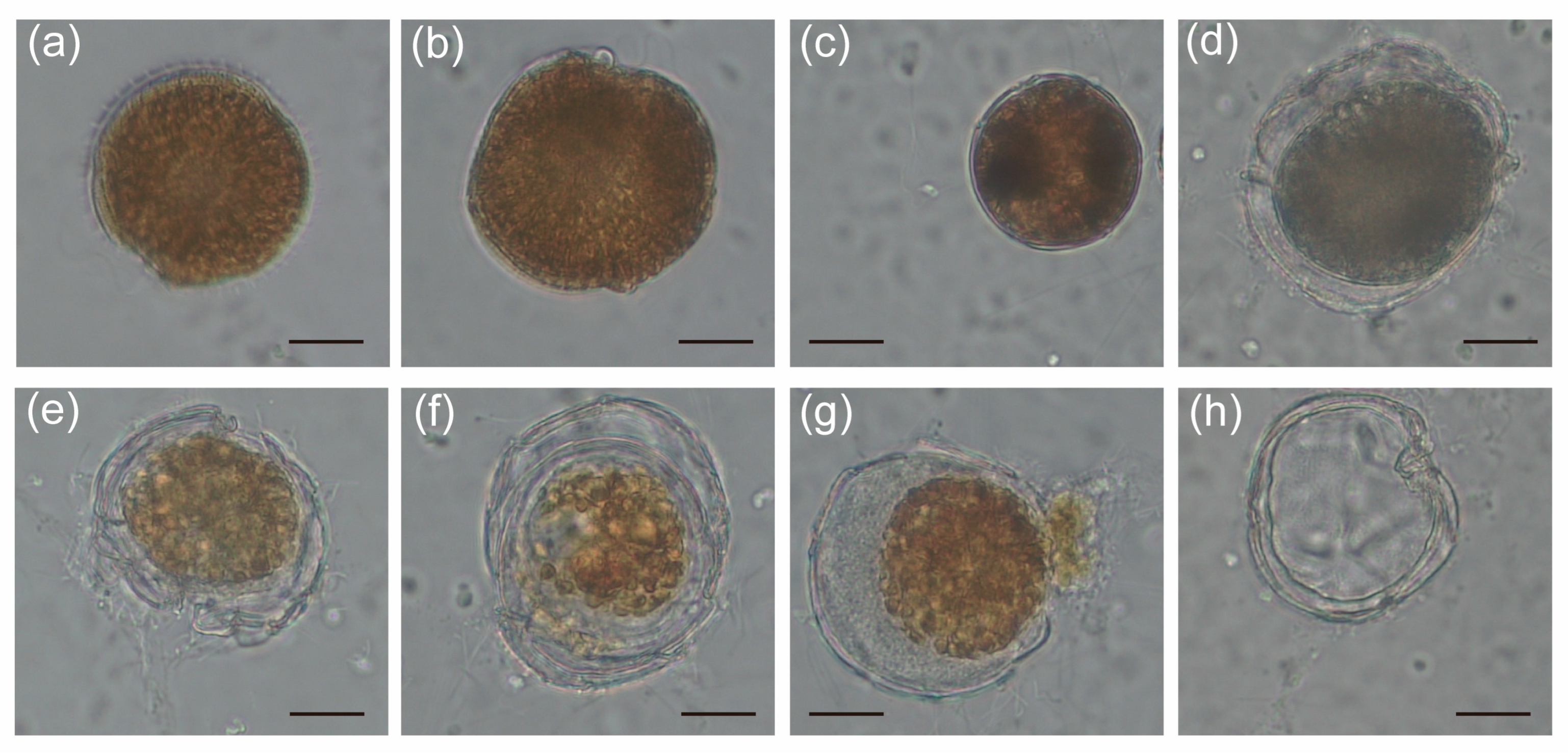
2.3. The Alteration of Morphology and Toxicity of G. balechii 1123M1M10 by Infection
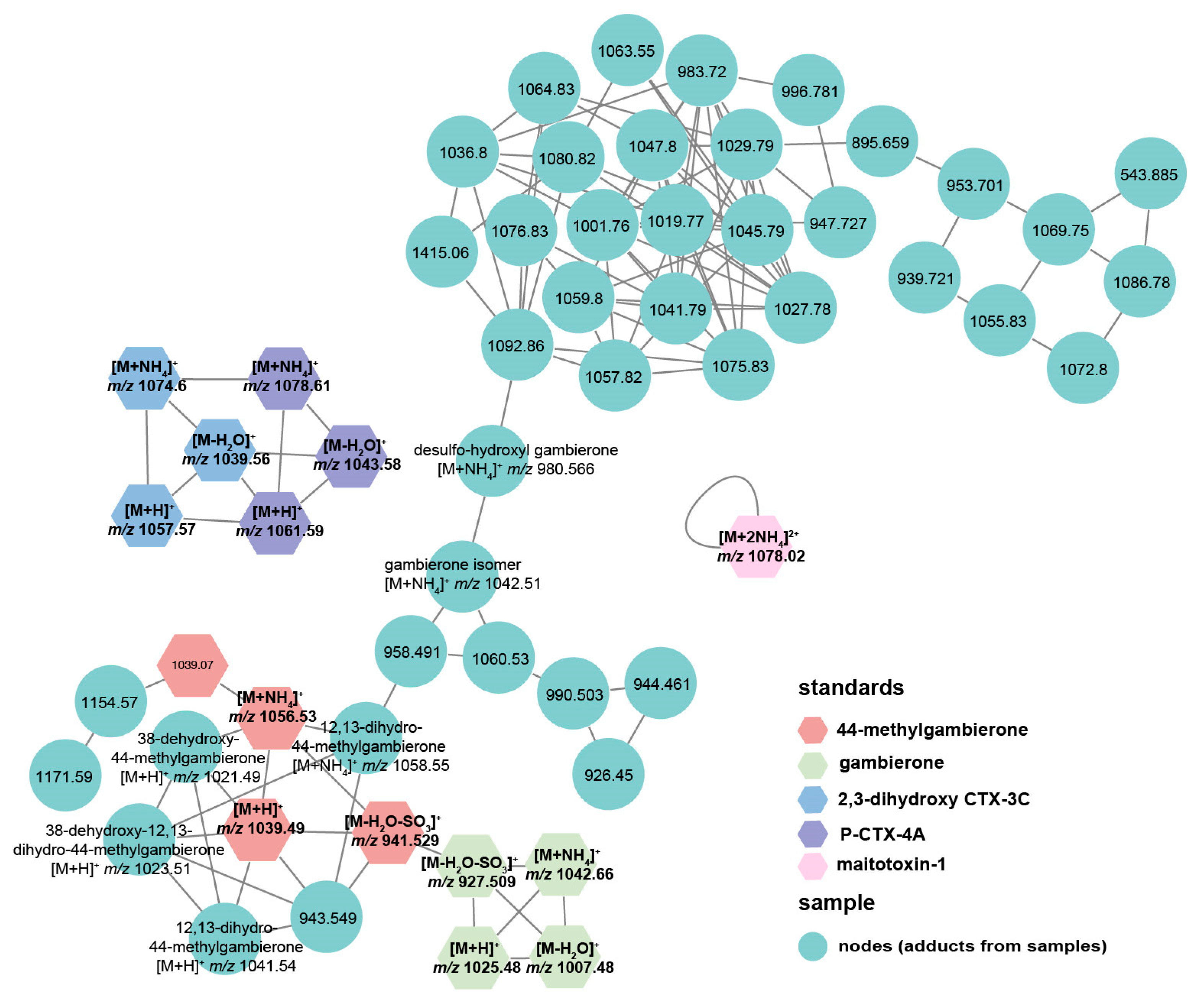
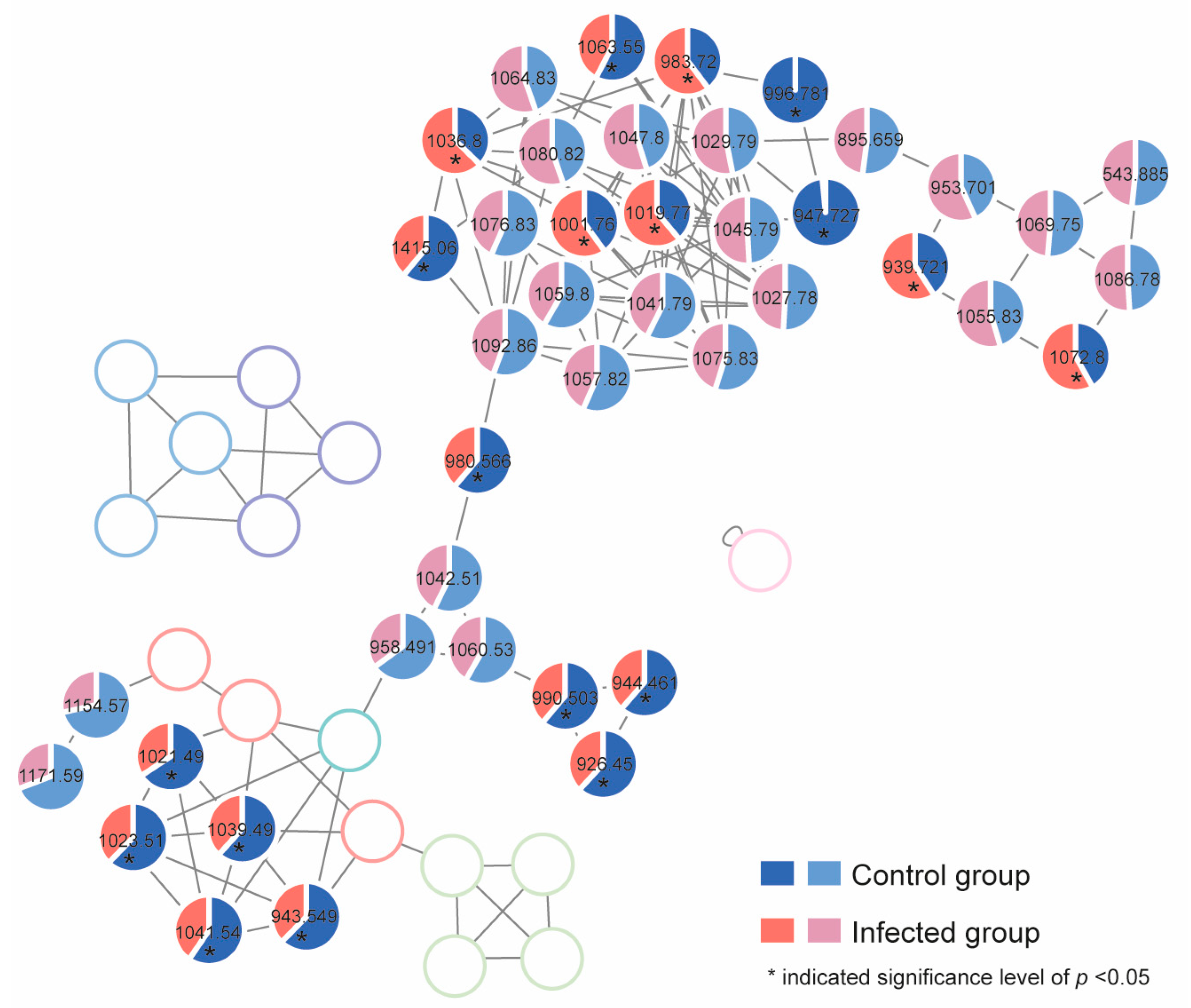
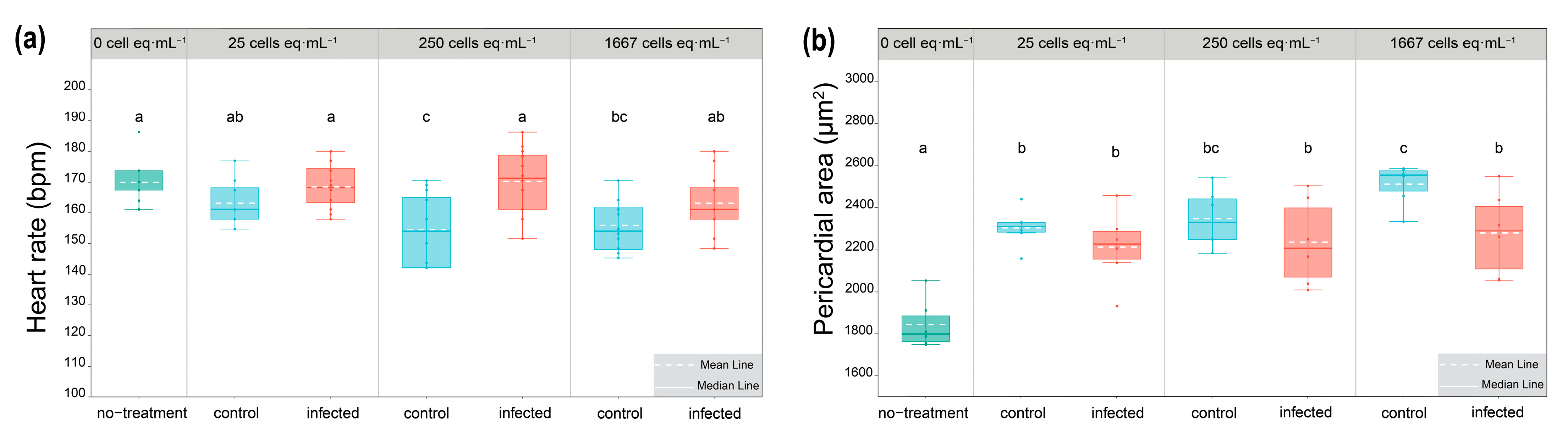
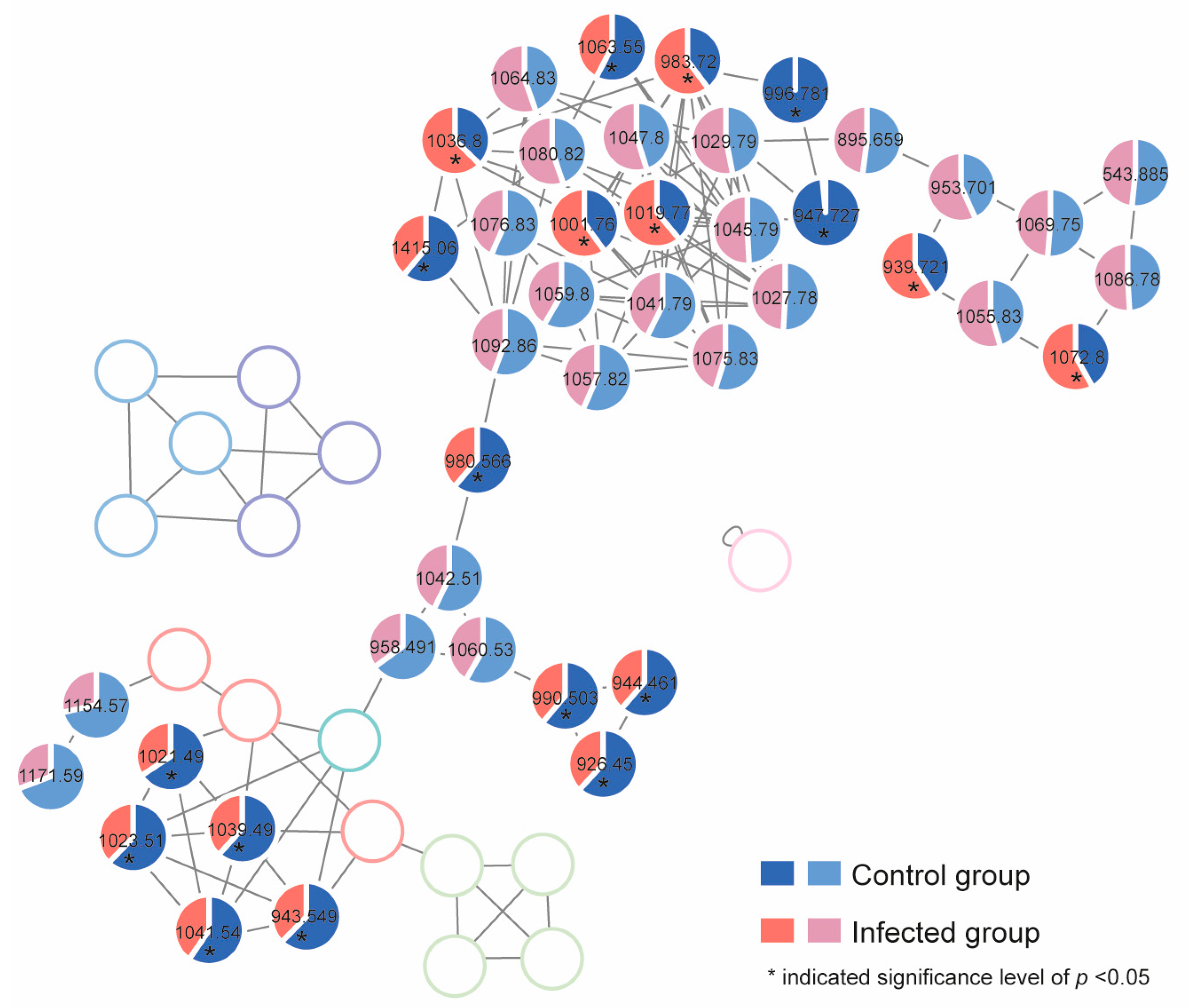
2.4. Response of Algal Toxin Production to Pseudobodo sp. Infection
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Gambierdiscus Balechii 1123M1M10 and Pseudobodo sp. Cultivation
5.2. Co-Culture Experiments
5.3. Light Microscopy and Electron Microscopy
5.4. DNA Extraction, PCR Amplification, and Sequencing for Pseudobodo sp.
5.5. Pseudobodo sp. SSU rRNA Gene Alignment and Phylogenetic Analyses
5.6. High-Throughput Sequencing of Microbiome 16S rRNA Gene Sequencing
5.7. Assessment of Cardiotoxicity of Algal Extracts in Zebrafish
5.8. Sample Collection and Treatment for HPLC-MS/MS Analysis
5.9. Toxin Standards
5.10. Instrumental Analysis
5.11. Untargeted Toxin Profiling and Relative Quantification
5.12. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez, F. Diversity, and classification of dinoflagellates. In Dinoflagellates: Classification, Evolution, Physiology and Ecological Significance; Rao, D.V.S., Ed.; Nova Science Publishers, Inc.: Puerto de Santa María, Spain, 2020; pp. 1–38. ISBN 978-1-53617-888-3. [Google Scholar]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Roth, P.B.; Twiner, M.J.; Mikulski, C.M.; Barnhorst, A.B.; Doucette, G.J. Comparative analysis of two algicidal bacteria active against the red tide dinoflagellate Karenia brevis. Harmful Algae 2008, 7, 682–691. [Google Scholar] [CrossRef]
- Fu, L.; An, X.; Li, D.; Zhou, L.; Tian, Y.; Zheng, T. Isolation and alga-inhibiting characterization of Vibrio sp. BS02 against Alexandrium tamarense. World J. Microbiol. Biotechnol. 2011, 27, 2949–2956. [Google Scholar] [CrossRef]
- Zheng, T.L.; Su, J.Q.; Maskaoui, K.; Yu, Z.M.; Hu, Z.; Xu, J.S.; Hong, H.S. Microbial modulation in the biomass and toxin production of a red-tide causing alga. Mar. Pollut. Bull. 2005, 51, 1018–1025. [Google Scholar] [CrossRef]
- Molina-Grima, E.; García-Camacho, F.; Acién-Fernández, F.G.; Sánchez-Mirón, A.; Plouviez, M.; Shene, C.; Chisti, Y. Pathogens and predators impacting commercial production of microalgae and cyanobacteria. Biotechnol. Adv. 2022, 55, 107884. [Google Scholar] [CrossRef]
- Miller, J.J.; Delwiche, C.F.; Coats, D.W. Ultrastructure of Amoebophrya sp. and its changes during the course of infection. Protist 2012, 163, 720–745. [Google Scholar] [CrossRef]
- Reñé, A.; Alacid, E.; Ferrera, I.; Garcés, E. Evolutionary trends of Perkinsozoa (Alveolata) characters based on observations of two new genera of parasitoids of dinoflagellates, Dinovorax gen. nov. and Snorkelia gen. nov. Front. Microbiol. 2017, 8, 1594. [Google Scholar] [CrossRef]
- Garcés, E.; Alacid, E.; Reñé, A.; Petrou, K.; Simó, R. Host-released dimethylsulphide activates the dinoflagellate parasitoid Parvilucifera sinerae. ISME J. 2013, 7, 1065–1068. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Y.; Wu, J.; Yin, Q.; Wang, P.; Zhu, J.; Chan, L.L.; Wu, B. Characterization of new gambierones produced by Gambierdiscus balechii 1123M1M10. Mar. Drugs 2023, 21, 3. [Google Scholar] [CrossRef]
- Chateau-Degat, M.L.; Chinain, M.; Cerf, N.; Gingras, S.; Hubert, B.; Dewailly, É. Seawater temperature, Gambierdiscus spp. variability and incidence of ciguatera poisoning in French Polynesia. Harmful Algae 2005, 4, 1053–1062. [Google Scholar] [CrossRef]
- Watari, T.; Tachibana, T.; Okada, A.; Nishikawa, K.; Otsuki, K.; Nagai, N.; Abe, H.; Nakano, Y.; Takagi, S.; Amano, Y. A review of food poisoning caused by local food in Japan. J. Gen. Fam. Med. 2021, 22, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fleming, L.E.; Fernandez, M.; Bienfang, P.; Schrank, K.; Dickey, R.; Bottein, M.Y.; Backer, L.; Ayyar, R.; Weisman, R.; et al. Ciguatera fish poisoning: Treatment, prevention and management. Mar. Drugs 2008, 6, 456–479. [Google Scholar] [CrossRef] [PubMed]
- Soliño, L.; Costa, P.R. Global impact of ciguatoxins and ciguatera fish poisoning on fish, fisheries and consumers. Environ. Res. 2020, 182, 109111. [Google Scholar] [CrossRef]
- Kodama, M.; Doucette, G.J.; Green, D.H. Relationships between Bacteria and Harmful Algae. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 243–255. [Google Scholar]
- Sakami, T.; Nakahara, H.; Chinain, M.; Ishida, Y. Effects of epiphytic bacteria on the growth of the toxic dinoflagellate Gambierdiscus toxicus (Dinophyceae). J. Exp. Mar. Bio. Ecol. 1999, 233, 231–246. [Google Scholar] [CrossRef]
- Wu, Z.; Lee, W.H.; Liu, Z.; Lin, S.; Lam, P.K.S. Microbiome associated with Gambierdiscus balechii cultures under different toxicity conditions. Front. Mar. Sci. 2022, 9, 760553. [Google Scholar] [CrossRef]
- Jirsová, D.; Füssy, Z.; Richtová, J.; Gruber, A.; Oborník, M. Morphology, ultrastructure, and mitochondrial genome of the marine non-photosynthetic bicosoecid cafileria marina gen. Et sp. nov. Microorganisms 2019, 7, 240. [Google Scholar] [CrossRef]
- Arndt, H.; Dietrich, D.; Auer, B.; Cleven, E.; Gräfenhan, T.; Weitere, M. Functional diversity of heterotrophic flagellates in aquatic ecosystems. In The Flagellates: Unity, Diversity and Evolution; Taylor & Francis: London, UK, 2000; pp. 240–268. [Google Scholar]
- Karpov, S. Ultrastructure of the aloricate bicosoecid Pseudobodo tremulans, with revision of the order Bicosoecida. Protistology 2000, 1, 101–109. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. 2022. Available online: http://www.algaebase.org (accessed on 6 March 2022).
- Grießmann, K. Über marine Flagellaten. J. Chem. Inf. Model. 1913, 53, 1689–1699. [Google Scholar]
- Parslow, J.; Doucette, G.; Taylor, F.; Harrison, P. Feeding by the zooflagellate Pseudobodo sp. on the picoplanktonic prasinomonad Micromonas pusilla. Mar. Ecol. Prog. Ser. 1986, 29, 237–246. [Google Scholar] [CrossRef]
- Chen, Z.; Lei, X.; Zhang, B.; Yang, L.; Zhang, H.; Zhang, J.; Li, Y.; Zheng, W.; Tian, Y.; Liu, J.; et al. First report of Pseudobodo sp., a new pathogen for a potential energy-producing algae: Chlorella vulgaris cultures. PLoS ONE 2014, 9, e89517. [Google Scholar] [CrossRef]
- Cavalier-Smith, T.; Chao, E.E.Y. Phylogeny and megasystematics of phagotrophic heterokonts (kingdom Chromista). J. Mol. Evol. 2006, 62, 388–420. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a ladder-shaped polyether from the dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef]
- Yon, T.; Sibat, M.; Robert, E.; Lhaute, K.; Holland, W.C.; Litaker, R.W.; Bertrand, S.; Hess, P.; Réveillon, D. Sulfo-gambierones, two new analogs of gambierone produced by Gambierdiscus excentricus. Mar. Drugs 2021, 19, 657. [Google Scholar] [CrossRef]
- Li, W.; Yan, R.; Yu, Y.; Shi, Z.; Mándi, A.; Shen, L.; Kurtán, T.; Wu, J. Determination of the absolute configuration of super-carbon-chain compounds by a combined chemical, spectroscopic, and computational approach: Gibbosols A and B. Angew. Chem. 2020, 132, 13128–13136. [Google Scholar] [CrossRef]
- Fenchel, T. Ecology of heterotrophic microflagellates. I. some important forms and their functional morphology. Mar. Ecol. Prog. Ser. 1982, 8, 211–223. [Google Scholar] [CrossRef]
- Mayali, X.; Doucette, G.J. Microbial community interactions and population dynamics of an algicidal bacterium active against Karenia brevis (Dinophyceae). Harmful Algae 2002, 1, 277–293. [Google Scholar] [CrossRef]
- Bolch, C.J.S.; Subramanian, T.A.; Green, D.H. The toxic dinoflagellate Gymnodinium catenatum (Dinophyceae) requires marine bacteria for growth. J. Phycol. 2011, 47, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Coyne, K.J.; Wang, Y.; Johnson, G. Algicidal Bacteria: A review of current knowledge and applications to control Harmful Algal Blooms. Front. Microbiol. 2022, 13, 871177. [Google Scholar] [CrossRef] [PubMed]
- Hold, G.L.; Smith, E.A.; RappÃ, M.S.; Maas, E.W.; Moore, E.R.; Stroempl, C.; Stephen, J.R.; Prosser, J.I.; Birkbeck, T.H.; Gallacher, S. Characterisation of bacterial communities associated with toxic and non-toxic dinoflagellates: Alexandrium spp. and Scrippsiella trochoidea. FEMS Microbiol. Ecol. 2006, 37, 161–173. [Google Scholar] [CrossRef]
- Pan, X.; Raaijmakers, J.M.; Carrión, V.J. Importance of bacteroidetes in host–microbe interactions and ecosystem functioning. Trends Microbiol. 2023, 31, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Sapp, M.; Schwaderer, A.S.; Wiltshire, K.H.; Hoppe, H.G.; Gerdts, G.; Wichels, A. Species-specific bacterial communities in the phycosphere of microalgae? Microb. Ecol. 2007, 53, 683–699. [Google Scholar] [CrossRef] [PubMed]
- Curson, A.R.J.; Liu, J.; Bermejo Martínez, A.; Green, R.T.; Chan, Y.; Carrión, O.; Williams, B.T.; Zhang, S.H.; Yang, G.P.; Bulman Page, P.C.; et al. Dimethylsulfoniopropionate biosynthesis in marine bacteria and identification of the key gene in this process. Nat. Microbiol. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.M.; Jones, G.B.; Deseo, M.A.; Shepherd, K.M.; Kiene, R.P.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Effects of environmental factors on dimethylated sulfur compounds and their potential role in the antioxidant system of the coral holobiont. Limnol. Oceanogr. 2014, 59, 758–768. [Google Scholar] [CrossRef]
- Zhong, H.; Sun, H.; Liu, R.; Zhan, Y.; Huang, X.; Ju, F.; Zhang, X. Comparative genomic analysis of Labrenzia aggregate (Alphaproteobacteria) strains isolated from the Mariana trench: Insights into the metabolic potentials and biogeochemical functions. Front. Microbiol. 2021, 12, 770370. [Google Scholar] [CrossRef]
- Davies, M.S.; Hawkins, S.J. Mucus from marine molluscs. Adv. Mar. Biol. 1998, 34, 1–71. [Google Scholar]
- Fraga, S.; Rodríguez, F.; Riobó, P.; Bravo, I. Gambierdiscus balechii sp. nov. (Dinophyceae), a new benthic toxic dinoflagellate from the Celebes Sea (SW Pacific Ocean). Harmful Algae 2016, 58, 93–105. [Google Scholar] [CrossRef]
- Wu, Z.; Luo, H.; Yu, L.; Lee, W.H.; Li, L.; Mak, Y.L.; Lin, S.; Lam, P.K.S. Characterizing ciguatoxin (CTX)- and non-CTX-producing strains of Gambierdiscus balechii using comparative transcriptomics. Sci. Total Environ. 2020, 717, 137184. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, C.J.S.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genom. 2015, 16, 1–10. [Google Scholar] [CrossRef]
- Gallimore, A.R.; Spencer, J.B. Stereochemical uniformity in marine polyether ladders—Implications for the biosynthesis and structure of maitotoxin. Angew. Chem.—Int. Ed. 2006, 45, 4406–4413. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Mak, Y.L.; Lu, C.K.; Mei, H.H.; Wu, J.J.; Lee, W.H.; Chan, L.L.; Lim, P.T.; Mustapa, N.I.; Lim, H.C.; et al. Taxonomic assignment of the benthic toxigenic dinoflagellate Gambierdiscus sp. type 6 as Gambierdiscus balechii (Dinophyceae), including its distribution and ciguatoxicity. Harmful Algae 2017, 67, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Joo, J.H.; Park, B.S.; Kim, J.H.; Kim, J.H.; Han, M.S. Relationship between dissolved organic carbon and bacterial community in the coastal waters of Incheon, Korea. Oceanol. Hydrobiol. Stud. 2017, 46, 50–61. [Google Scholar] [CrossRef]
- Massana, R.; Camp, J.; Icmcsic, M.; Marı, P. Description, host-specificity, and strain selectivity of the dinoflagellate parasite Parvilucifera sinerae. Protist 2008, 159, 563–578. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Huang, M.; Jiao, J.; Wang, J.; Xia, Z.; Zhang, Y. Characterization of acrylamide-induced oxidative stress and cardiovascular toxicity in zebrafish embryos. J. Hazard. Mater. 2018, 347, 451–460. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Ma, Y.; Wu, J.; Wang, P.; Wang, Y.; Wang, A.; Yin, Q.; Ma, H.; Chan, L.L.; Wu, B. Characterizing the Influence of a Heterotrophic Bicosoecid Flagellate Pseudobodo sp. on the Dinoflagellate Gambierdiscus balechii. Toxins 2023, 15, 657. https://doi.org/10.3390/toxins15110657
Liu X, Ma Y, Wu J, Wang P, Wang Y, Wang A, Yin Q, Ma H, Chan LL, Wu B. Characterizing the Influence of a Heterotrophic Bicosoecid Flagellate Pseudobodo sp. on the Dinoflagellate Gambierdiscus balechii. Toxins. 2023; 15(11):657. https://doi.org/10.3390/toxins15110657
Chicago/Turabian StyleLiu, Xiaowan, Yihan Ma, Jiajun Wu, Pengbin Wang, Yinuo Wang, Anli Wang, Qizhao Yin, Haiying Ma, Leo Lai Chan, and Bin Wu. 2023. "Characterizing the Influence of a Heterotrophic Bicosoecid Flagellate Pseudobodo sp. on the Dinoflagellate Gambierdiscus balechii" Toxins 15, no. 11: 657. https://doi.org/10.3390/toxins15110657
APA StyleLiu, X., Ma, Y., Wu, J., Wang, P., Wang, Y., Wang, A., Yin, Q., Ma, H., Chan, L. L., & Wu, B. (2023). Characterizing the Influence of a Heterotrophic Bicosoecid Flagellate Pseudobodo sp. on the Dinoflagellate Gambierdiscus balechii. Toxins, 15(11), 657. https://doi.org/10.3390/toxins15110657