Phytotoxicity and Phytotoxic Substances in Calamus tenuis Roxb.

 ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Results
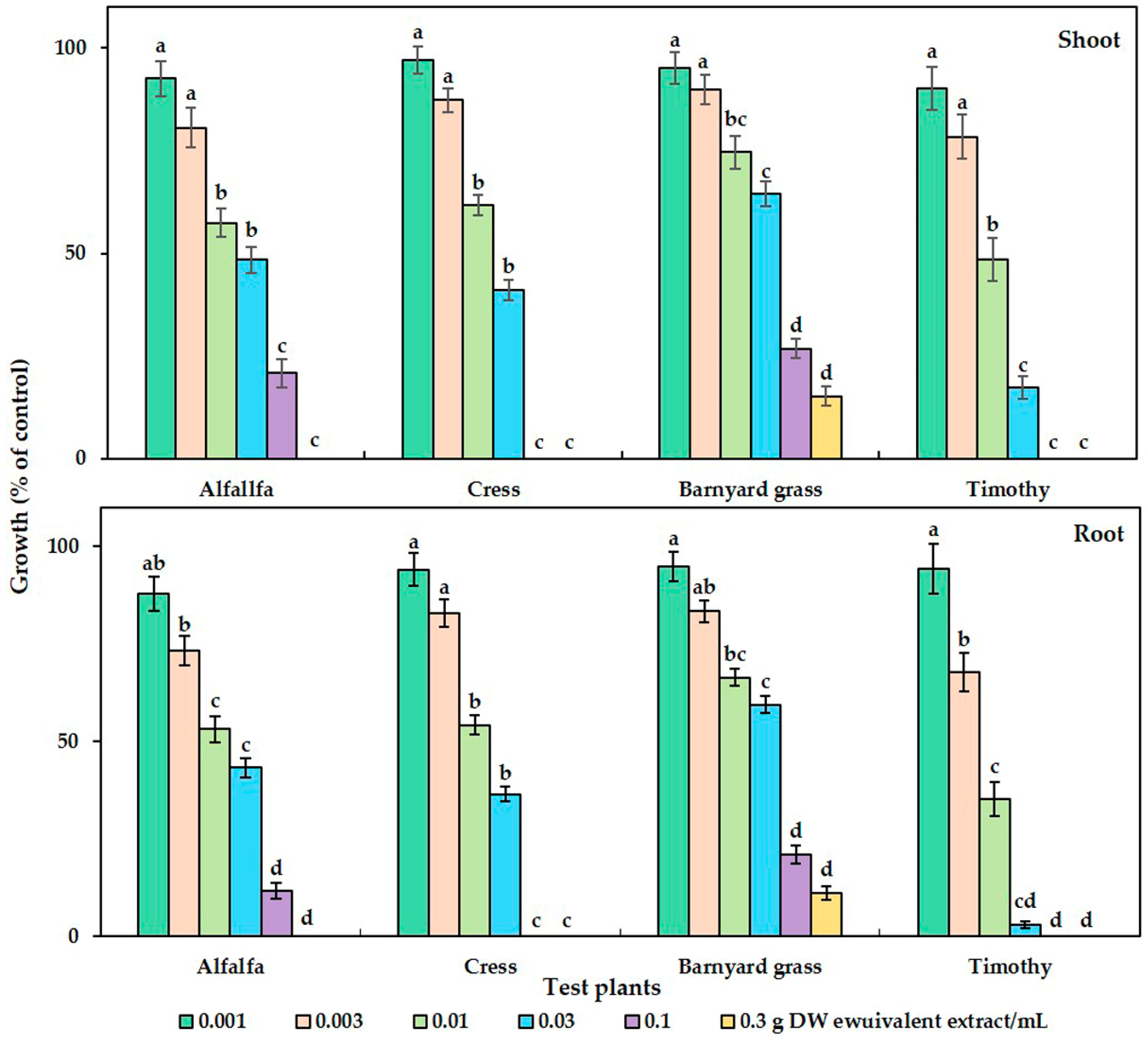
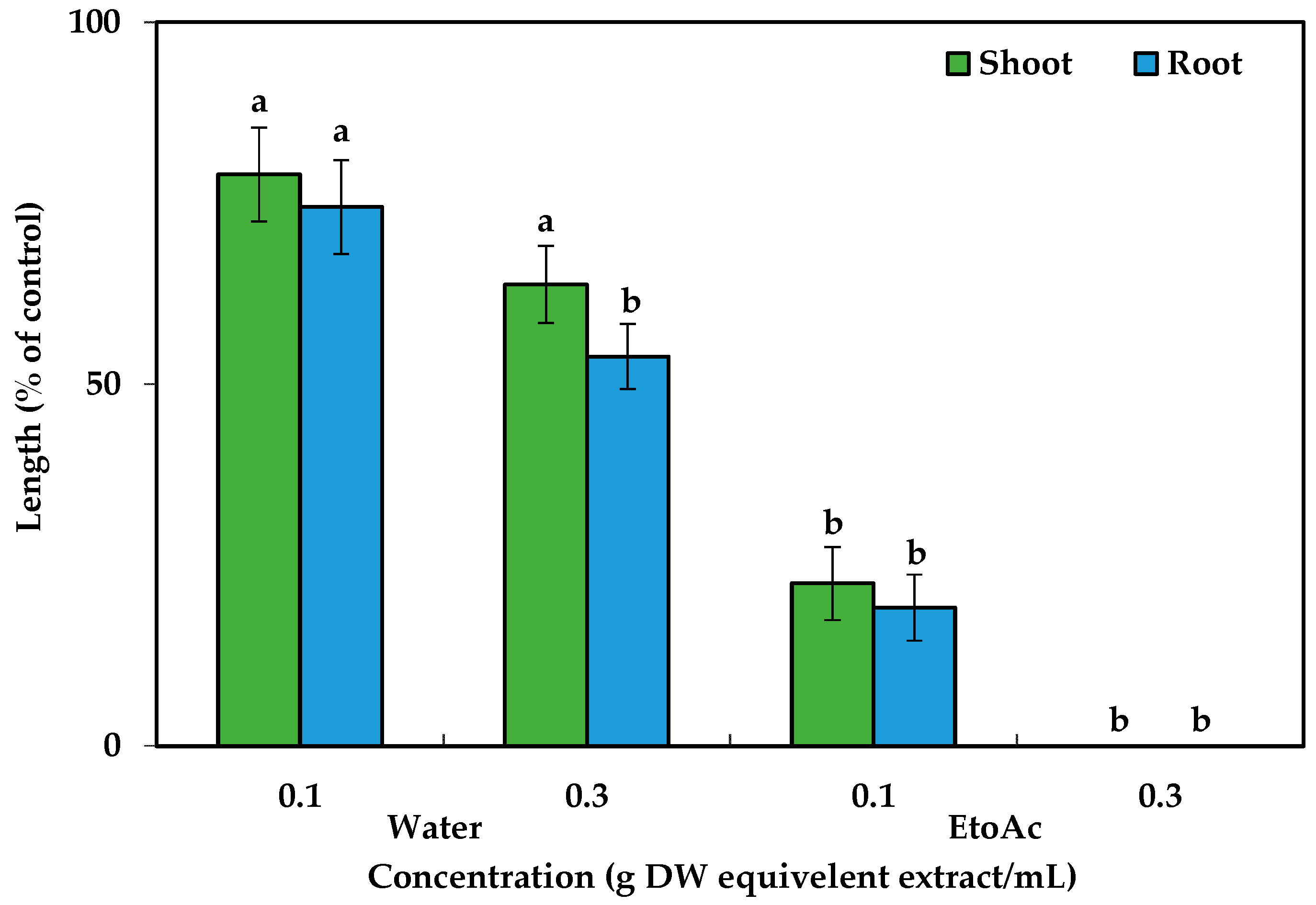
2.1. Phytotoxicity of the C. tenuis Extract
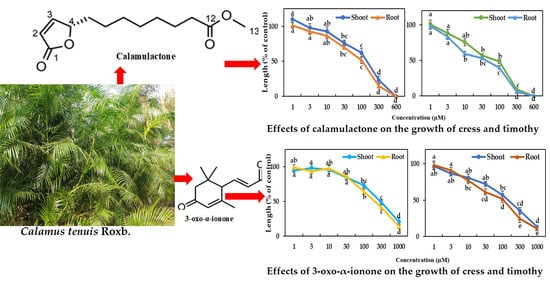
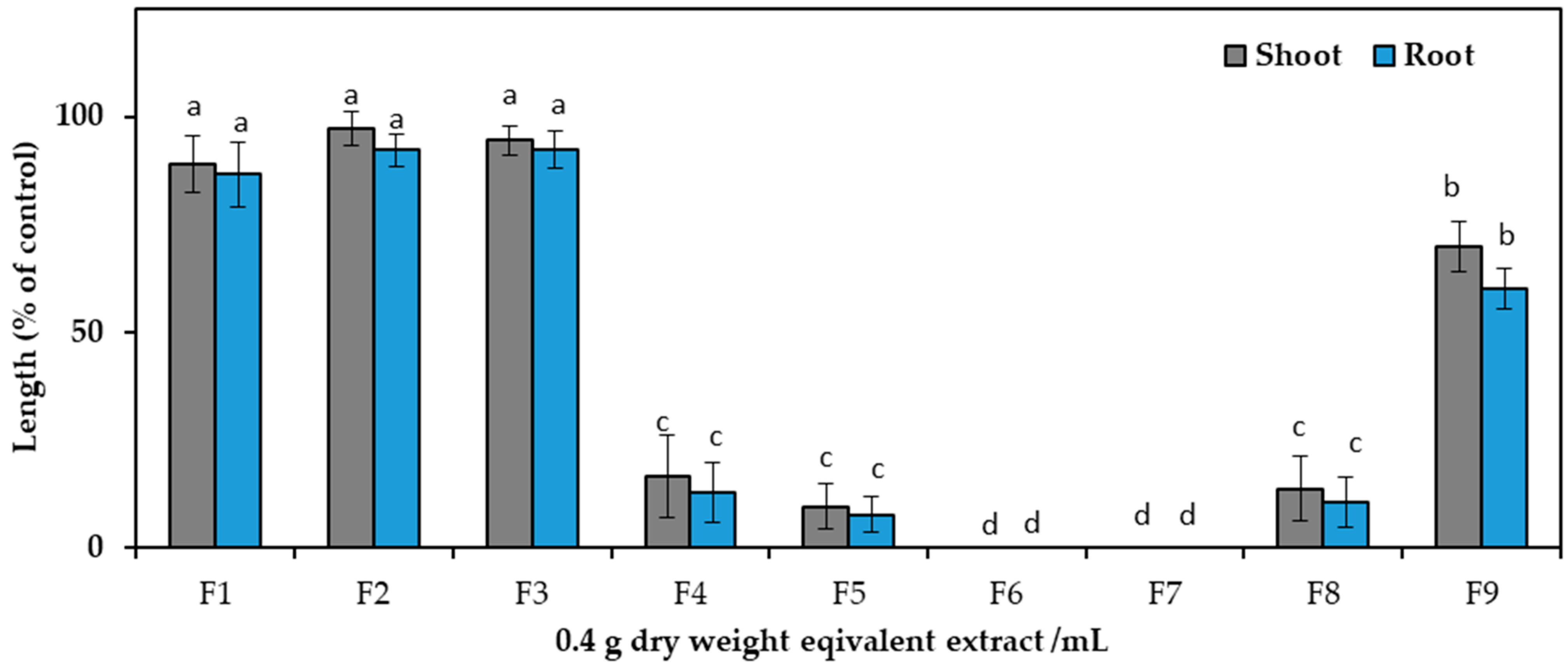
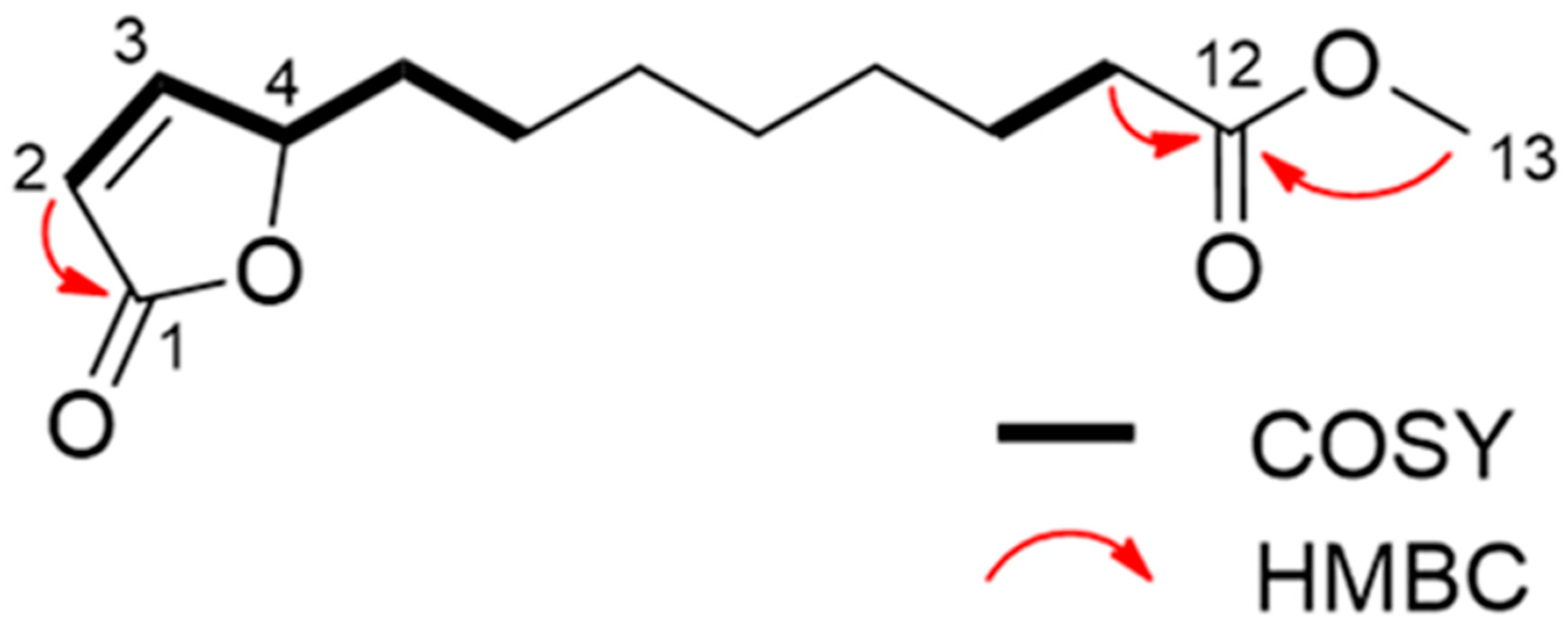
2.2. Isolation and Characterization of the Active Substances
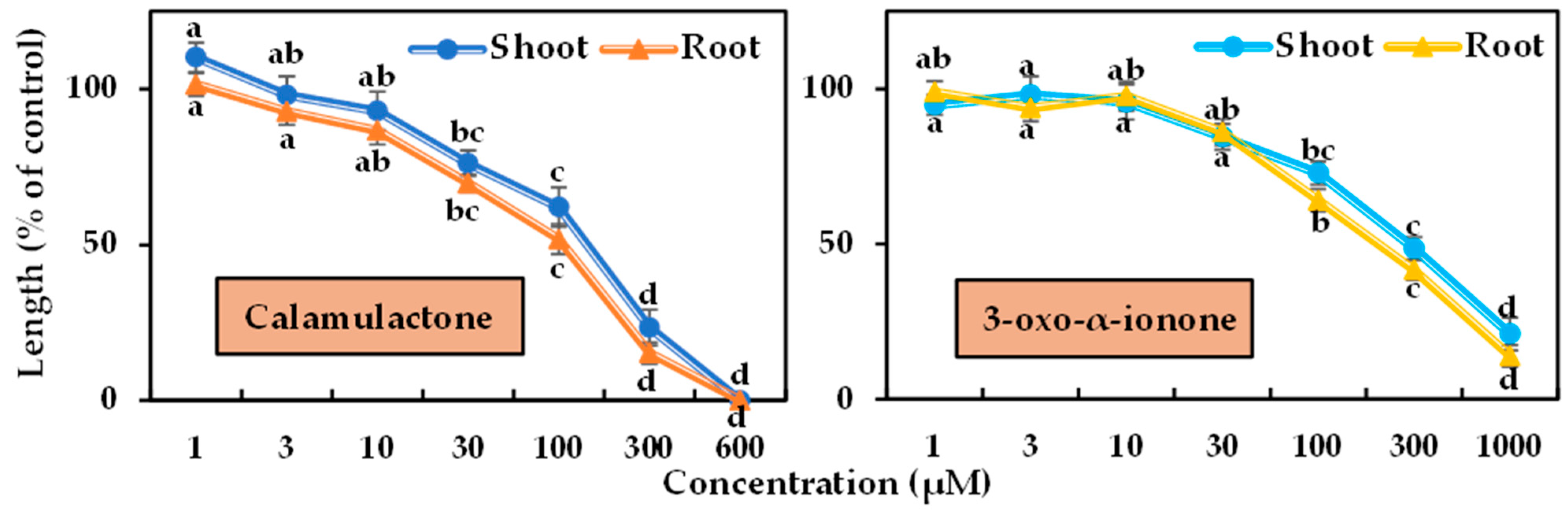
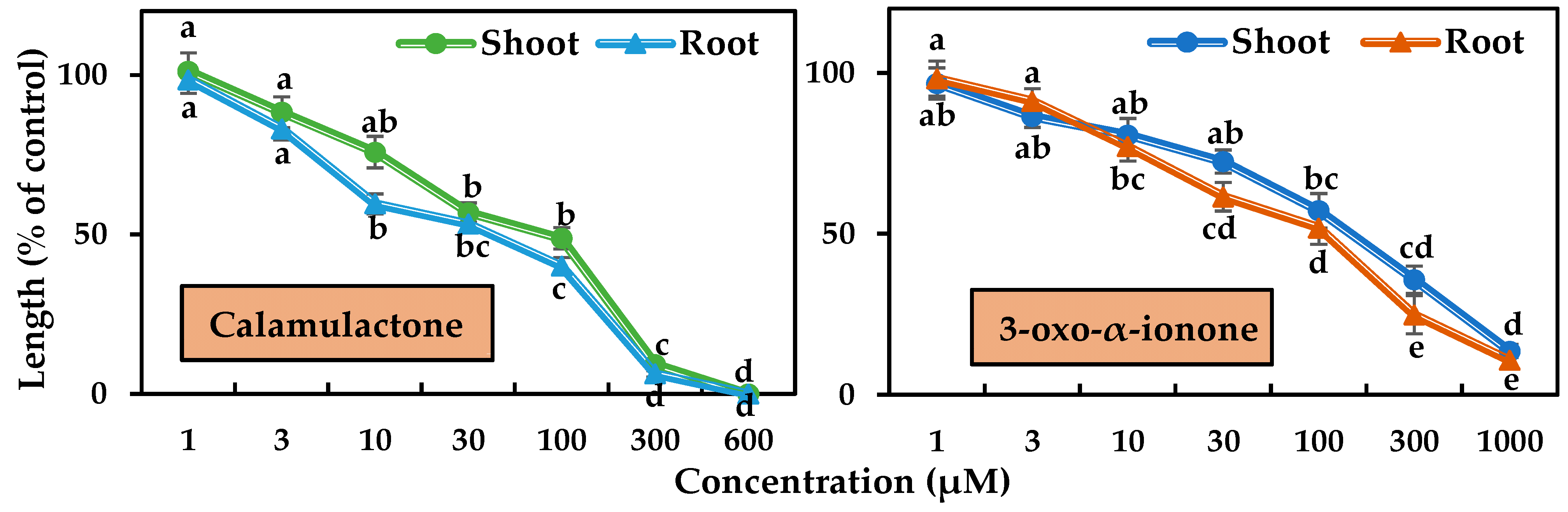
2.3. Phytotoxicity of the Identified Substances
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Samples
5.2. Extraction and Bioassay
5.3. Purification of the Active Substances
5.4. Characterization of the Compounds
Spectral Data
5.5. Bioassay of Calamulactone and 3-Oxo-α-Ionone
5.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvarado-Serrano, D.F.; Van Etten, M.L.; Chang, S.M.; Baucom, R.S. The relative contribution of natural landscapes and human-mediated factors on the connectivity of a noxious invasive weed. Heredity 2019, 122, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Ireland, K.B.; Van Klinken, R.; Cook, D.C.; Logan, D.; Jamieson, L.; Tyson, J.L.; Paini, D. Plant pest impact metric system (PPIMS): Framework and guidelines for a common set of metrics to classify and prioritise plant pests. Crop Prot. 2020, 128, 105003. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crops Res. 2015, 183, 56–68. [Google Scholar] [CrossRef]
- Adetunji, C.O.; Oloke, J.K.; Bello, O.M.; Pradeep, M.; Jolly, R.S. Isolation, structural elucidation and bioherbicidal activity of an eco-friendly bioactive 2-(hydroxymethyl) phenol, from Pseudomonas aeruginosa (C1501) and its ecotoxicological evaluation on soil. Environ. Technol. Innovat. 2019, 13, 304–317. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Moss, S. Integrated weed management (IWM): Why are farmers reluctant to adopt non-chemical alternatives to herbicides? Pest Manag. Sci. 2019, 75, 1205–1211. [Google Scholar] [CrossRef]
- Hinz, H.L.; Winston, R.L.; Schwarzländer, M. A global review of target impact and direct nontarget effects of classical weed biological control. Curr. Opin. Insect. Sci. 2020, 38, 48–54. [Google Scholar] [CrossRef]
- Davis, A.M.; Neelamraju, C. Quantifying water quality improvements through use of precision herbicide application technologies in a dry-tropical, furrow-irrigated cropping system. Water 2019, 11, 2326. [Google Scholar] [CrossRef]
- Comont, D.; Lowe, C.; Hull, R.; Crook, L.; Hicks, H.L.; Onkokesung, N.; Neve, P. Evolution of generalist resistance to herbicide mixtures reveals a trade-off in resistance management. Nat. Commun. 2020, 11, 3086. [Google Scholar] [CrossRef]
- Herbicide Resistance Action Committee. Available online: https://hracglobal.com/herbicide-resistance/overview (accessed on 27 August 2023).
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 2015, 6, 566. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Bakhtiari, M.; Formenti, L.; Xiao, Z.; Defossez, E.; Rasmann, S. Biological control beneath the feet: A review of crop protection against insect root herbivores. Insects 2016, 7, 70. [Google Scholar] [CrossRef]
- Carrubba, A.; Labruzzo, A.; Comparato, A.; Muccilli, S.; Spina, A. Use of plant water extracts for weed control in durum wheat (Triticum turgidum L. subsp. durum Desf.). Agronomy 2020, 10, 364. [Google Scholar] [CrossRef]
- Takahashi, J.A.; Barbosa, B.V.; Martins, B.D.A.; Guirlanda, P.C.; Moura, A.F.M. Use of the versatility of fungal metabolism to meet modern demands for healthy aging, functional foods, and sustainability. J. Fungi 2020, 6, 223. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.M.; Henderson, A.; Nguyễn, Q.D.N.; Ledecq, T. Systematics, Ecology, and Management of Rattans in Cambodia, Laos, and Vietnam: The Biological Bases of Sustainable Use; Agricultural Publishing House: WWF-Greater Mekong and The New York Botanical Garden: New York, NY, USA, 2014; p. 235. [Google Scholar]
- Ahmed, Z.U.; Bithi, S.S.; Khan, M.M.R.; Hossain, M.M.; Sharmin, S.; Rony, S.R. Phytochemical screening, antioxidant and cytotoxic activity of fruit extracts of Calamus tenuis Roxb. J. Coast. Life Med. 2014, 2, 645–650. [Google Scholar] [CrossRef]
- Alam, M.K. Rattans of Bangladesh; Bulletin 7, Taxonomy Series; Bangladesh Forest Research Institute: Chittagong, Bangladesh, 1990; p. 35. [Google Scholar]
- Manohara, T.N. Nutritional evaluation of shoots of two rattans of northeast India—Calamus flagellum Griff. ex Mart. and C. floribundus Griff. (Arecaceae). Econ. Bot. 2013, 67, 263–268. Available online: http://www.jstor.org/stable/43304349 (accessed on 20 August 2023). [CrossRef]
- Islam, S.A.; Miah, M.A.Q.; Habib, M.A.; Rasul, M.G. Growth performance of Calamus tenuis Roxb. (Jali bet) in the coastal homesteads of Bangladesh. J. Biosci. Agric. Res. 2015, 4, 74–79. [Google Scholar] [CrossRef]
- Khare, C.P. Indian Medicinal Plants: An Illustrated Dictionary, 1st ed.; Springer: New York, NY, USA, 2008; p. 835. [Google Scholar] [CrossRef]
- Tag, H.; Kalita, P.; Dwivedi, P.; Das, A.K.; Namsa, N.D. Herbal medicines used in the treatment of diabetes mellitus in Arunachal Himalaya, northeast, India. J. Ethnopharmacol. 2012, 141, 786–795. [Google Scholar] [CrossRef]
- Yu, G.F.; Mulabagal, V.; Diyabalanage, T.; Hurtada, W.A.; DeWitt, D.L.; Nair, M.G. Non-nutritive functional agents in rattan-shoots, a food consumed by native people in the Philippines. Food Chem. 2008, 110, 991–996. [Google Scholar] [CrossRef]
- Saikia, P.; Khan, M.L. Diversity of medicinal plants and their uses in home gardens of upper Assam, Northeast India. Asian J. Pharm. Biol. Res. 2011, 1, 296–309. [Google Scholar]
- Sarkar, H.; Sharin, T.; Al Mamun, M.S. Assessment of Calamus tenuis fruits extract on blood glucose level elevation and its antibacterial potency. J. Appl. Pharm. Res. 2018, 6, 1–6. [Google Scholar] [CrossRef]
- Hossain, M.S. Analgesic and neuropharmacological activity of methanolic extract of Calamus tenuis Roxb. Fruits. J. Sci. Innov. 2013, 2, 1067–1072. [Google Scholar]
- Thakur, P.K.; Sheth, M.; Bhambra, G.K.; Nagar, P.S.; Upadhyay, K.; Devkar, R. Phytochemical composition and cytotoxic potential of edible rattan (Calamus tenuis Roxb.) shoot extracts on MCF7 and A549 cells. World J. Pharm. Res. 2016, 5, 1738–1746. [Google Scholar]
- Kuo, Y.H.; Huang, S.L.; Chang, C.I. A phenolic and an aliphatic lactone from Diospyros maritima. Phytochemistry 1998, 49, 2505–2507. [Google Scholar] [CrossRef]
- Uchida, I.; Kuriyama, K. The π-π circular dichroism of δβ-unsaturated γ-lactones. Tetrahedron Lett. 1974, 15, 3761–3764. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Hayashi, Y.; Arita, M.; Hieda, T.; Mikami, Y. Microbial conversion of α-ionone, α-methylionone, and α-isomethylionone. Appl. Environ. Microbiol. 1998, 54, 2354–2360. [Google Scholar] [CrossRef]
- Chai, M.; Zhu, X.; Cui, H.; Jiang, C.; Zhang, J.; Shi, L. Lily cultivars have allelopathic potential in controlling Orobanche aegyptiaca Persoon. PLoS ONE 2015, 10, e0142811. [Google Scholar] [CrossRef]
- Al-Harbi, N.A. Allelopathic effect of leaf extract of two wild plants on seed germination, shoot and root length of two weed species; Portulaca oleracea and Chenopodium murale. Biosci. Biotechnol. Res. Asia 2018, 15, 929–935. [Google Scholar] [CrossRef]
- Wang, C.; Qi, J.; Liu, Q.; Wang, Y.; Wang, H. Allelopathic potential of aqueous extracts from Fleagrass (Adenosma buchneroides Bonati) against two crop and three weed species. Agriculture 2022, 12, 1103. [Google Scholar] [CrossRef]
- Ma, H.; Chen, Y.; Chen, J.; Zhang, Y.; Zhang, T.; He, H. Comparison of allelopathic effects of two typical invasive plants: Mikania micrantha and Ipomoea cairica in Hainan Island. Sci. Rep. 2020, 10, 11332. [Google Scholar] [CrossRef]
- Popovici, J.; Bertrand, C.; Jacquemoud, D.; Bellvert, F.; Fernandez, M.P.; Comte, G.; Piola, F. An allelochemical from Myrica gale with strong phytotoxic activity against highly invasive Fallopia×bohemica taxa. Molecules 2011, 16, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Tuyen, P.T.; Xuan, T.D.; Tu Anh, T.T.; Mai Van, T.; Ahmad, A.; Elzaawely, A.A.; Khanh, T.D. Weed suppressing potential and isolation of potent plant growth inhibitors from Castanea crenata Sieb. et Zucc. Molecules 2018, 23, 345. [Google Scholar] [CrossRef] [PubMed]
- Ladhari, A.; Omezzine, F.; DellaGreca, M.; Zarrelli, A.; Zuppolini, S.; Haouala, R. Phytotoxic activity of Cleome arabica L. and its principal discovered active compounds. S. Afr. J. Bot. 2013, 88, 341–351. [Google Scholar] [CrossRef]
- Khan, S.M.; Khanam, S.; Deepak, M.; Shivananda, B.G. Antioxidant activity of a new diarylheptanoid from Zingibero fficinale. Pharmacogn. Mag. 2006, 2, 254–257. [Google Scholar]
- Appiah, K.S.; Mardani, H.K.; Omari, R.A.; Eziah, V.Y.; Ofosu-Anim, J.; Onwona-Agyeman, S.; Fujii, Y. Involvement of carnosic acid in the phytotoxicity of Rosmarinus officinalis leaves. Toxins 2018, 10, 498. [Google Scholar] [CrossRef]
- Rob, M.; Hossen, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic activity and identification of phytotoxic substances from Schumannianthus dichotomus. Plants 2020, 9, 102. [Google Scholar] [CrossRef]
- Wei, C.; Zhou, S.; Li, W.; Jiang, C.; Yang, W.; Han, C.; Zhang, C.; Shao, H. Chemical composition and allelopathic, phytotoxic and pesticidal activities of Atriplex cana Ledeb. (Amaranthaceae) essential oil. Chem. Biodivers. 2019, 16, e1800595. [Google Scholar] [CrossRef]
- Madadi, E.; Fallah, S.; Sadeghpour, A.; Barani-Beiranvand, H. Exploring the use of chamomile (Matricaria chamomilla L.) bioactive compounds to control flixweed (Descurainia sophia L.) in bread wheat (Triticum aestivum L.): Implication for reducing chemical herbicide pollution. Saudi J. Biol. Sci. 2022, 29, 103421. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Zhu, Y.; Guo, L.; Ji, R.; Miao, Y.; Guo, L.; Du, H.; Liu, D. Caffeic acid, an allelochemical in Artemisia argyi, inhibits weed growth via suppression of mitogen-activated protein kinase signaling pathway and the biosynthesis of gibberellin and phytoalexin. Front. Plant Sci. 2022, 12, 802198. [Google Scholar] [CrossRef]
- Hossen, K.; Asato, Y.; Teruya, T.; Kato-Noguchi, H. Identification of four allelopathic compounds including a novel compound from Elaeocarpus floribundus Blume and determination of their allelopathic activity. J. Environ. Manag. 2023, 326, 116728. [Google Scholar] [CrossRef]
- Kobayashi, K. Factors affecting phytotoxic activity of allelochemicals in soil. Weed Biol. Manag. 2004, 4, 1–7. [Google Scholar] [CrossRef]
- Hanley, M.E.; Whiting, M.D. Insecticides and arable weeds: Effects on germination and seedling growth. Ecotoxicology 2005, 14, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Rob, M.M.; Iwasaki, A.; Suenaga, K.; Ozaki, K.; Teruya, T.; Kato-Noguchi, H. Potential use of Schumannianthus dichotomus waste: The phytotoxic activity of the waste and its identified compounds. J. Environ. Sci. Health Part B 2020, 55, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Omezzine, F.; Ladhari, A.; Rinez, A.; Haouala, R. Potent herbicidal activity of Inula crithmoïdes L. Sci. Hortic. 2011, 130, 853–861. [Google Scholar] [CrossRef]
- Miranda, M.A.; Varela, R.M.; Torres, A.; Molinillo, J.M.; Gualtieri, S.C.; Macías, F.A. Phytotoxins from Tithonia diversifolia. J. Nat. Prod. 2015, 78, 1083–1092. [Google Scholar] [CrossRef]
- Da Silva, B.P.; Nepomuceno, M.P.; Varela, R.M.; Torres, A.; Molinillo, J.M.; Alves, P.L.; Macías, F.A. Phytotoxicity study on Bidens sulphurea Sch. Bip. as a preliminary approach for weed control. J. Agric. Food Chem. 2017, 65, 5161–5172. [Google Scholar] [CrossRef]
- Jmii, G.; Molinillo, J.M.; Zorrilla, J.G.; Haouala, R. Allelopathic activity of Thapsia garganica L. leaves on lettuce and weeds, and identification of the active principles. S. Afr. J. Bot. 2020, 131, 188–194. [Google Scholar] [CrossRef]
- Rietjens, I.M.; Alink, G.M. Nutrition and health-toxic substances in food. Ned. Tijdschr. Geneeskd. 2003, 147, 2365–2370. [Google Scholar]
- Rauter, A.P.; Figueiredo, J.; Ismael, M.; Canda, T.; Font, J.; Figueredo, M. Efficient synthesis of α, β-unsaturated γ-lactones linked to sugars. Tetrahedron: Asymmetry 2001, 12, 1131–1146. [Google Scholar] [CrossRef]
- Resende, G.C.; Alvarenga, E.S.; Galindo, J.C.; Macias, F.A. Synthesis and phytotoxicity of 4,5 functionalized tetrahydrofuran-2-ones. J. Braz. Chem. Soc. 2012, 23, 2266–2270. [Google Scholar] [CrossRef]
- Hur, J.; Jang, J.; Sim, J. A review of the pharmacological activities and recent synthetic advances of γ-butyrolactones. Int. J. Mol. Sci. 2021, 22, 2769. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Putkaradze, N.; Bohnacker, S.; Jonczyk, R.; Fida, T.; Hoffmann, T.; Schwab, W. Six uridine-diphosphate glycosyltransferases catalyze the glycosylation of bioactive C13-apocarotenols. Plant Physiol. 2020, 184, 1744–1761. [Google Scholar] [CrossRef] [PubMed]
- Hossen, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxicity of the novel compound 3-hydroxy-4-oxo-β-dehydroionol and compound 3-oxo-α-ionone from Albizia richardiana (Voigt.) King & Prain. Environ. Technol. Innovat. 2021, 23, 101779. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Yamamoto, M.; Tamura, K.; Teruya, T.; Suenaga, K.; Fujii, Y. Isolation and identification of potent allelopathic substances in rattail fescue. Plant Growth Regul. 2010, 60, 127–131. [Google Scholar] [CrossRef]
- Llanos, G.G.; Varela, R.M.; Jiménez, I.A.; Molinillo, J.M.; Macías, F.A.; Bazzocchi, I.L. Metabolites from Withania aristata with potential phytotoxic activity. Nat. Prod. Commun. 2010, 5, 1934578X1000500712. [Google Scholar]
- Xian, Q.; Chen, H.; Liu, H.; Zou, H.; Yin, D. Isolation and identification of antialgal compounds from the leaves of Vallisneria spiralis L. by activity-guided fractionation. Environ. Sci. Pollut. Res. Int. 2006, 13, 233–237. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Plant Species | C. tenuis Aqueous Methanol Extract (mg DW/mL) | ||
|---|---|---|---|
| Shoot | Root | ||
| Dicotyledonous | Alfalfa | 24.60 | 14.24 |
| Cress | 18.65 | 12.97 | |
| Monocotyledonous | Barnyard grass | 47.57 | 40.33 |
| Timothy | 9.42 | 5.78 | |
| Position | δH Mult (J in Hz) a | δC b |
|---|---|---|
| 1 | 173.3 | |
| 2 | 6.11, dd (5.7, 2.0) | 121.8 |
| 3 | 7.44, dd (5.7, 1.5) | 156.3 |
| 4 | 5.03, m | 83.5 |
| 5a | 1.76, m | 33.3 |
| 5b | 1.66, m | |
| 6 | 1.44, m | 25.04 |
| 7 | 1.32, m c | 29.22 |
| 8 | 1.32, m c | 29.12 |
| 9 | 1.32, m c | 29.08 |
| 10 | 1.62, m | 24.98 |
| 11 | 2.30, t (7.4) | 34.2 |
| 12 | 174.4 | |
| 13 | 3.67, s | 51.6 |
| Test Plant | Calamulactone | 3-Oxo-α-Ionone | |
|---|---|---|---|
| (µM) | |||
| Cress | Shoot *** | 141.1 | 281.6 |
| Root *** | 105.6 | 199.2 | |
| Timothy | Shoot *** | 83.4 | 144.7 |
| Root *** | 40.5 | 107.8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rob, M.M.; Hossen, K.; Ozaki, K.; Teruya, T.; Kato-Noguchi, H. Phytotoxicity and Phytotoxic Substances in Calamus tenuis Roxb. Toxins 2023, 15, 595. https://doi.org/10.3390/toxins15100595
Rob MM, Hossen K, Ozaki K, Teruya T, Kato-Noguchi H. Phytotoxicity and Phytotoxic Substances in Calamus tenuis Roxb. Toxins. 2023; 15(10):595. https://doi.org/10.3390/toxins15100595
Chicago/Turabian StyleRob, Md. Mahfuzur, Kawsar Hossen, Kaori Ozaki, Toshiaki Teruya, and Hisashi Kato-Noguchi. 2023. "Phytotoxicity and Phytotoxic Substances in Calamus tenuis Roxb." Toxins 15, no. 10: 595. https://doi.org/10.3390/toxins15100595
APA StyleRob, M. M., Hossen, K., Ozaki, K., Teruya, T., & Kato-Noguchi, H. (2023). Phytotoxicity and Phytotoxic Substances in Calamus tenuis Roxb. Toxins, 15(10), 595. https://doi.org/10.3390/toxins15100595