Enzymatic Degradation of Zearalenone in the Gastrointestinal Tract of Pigs, Chickens, and Rainbow Trout

 , , ,
, , ,
Abstract
1. Introduction
2. Results
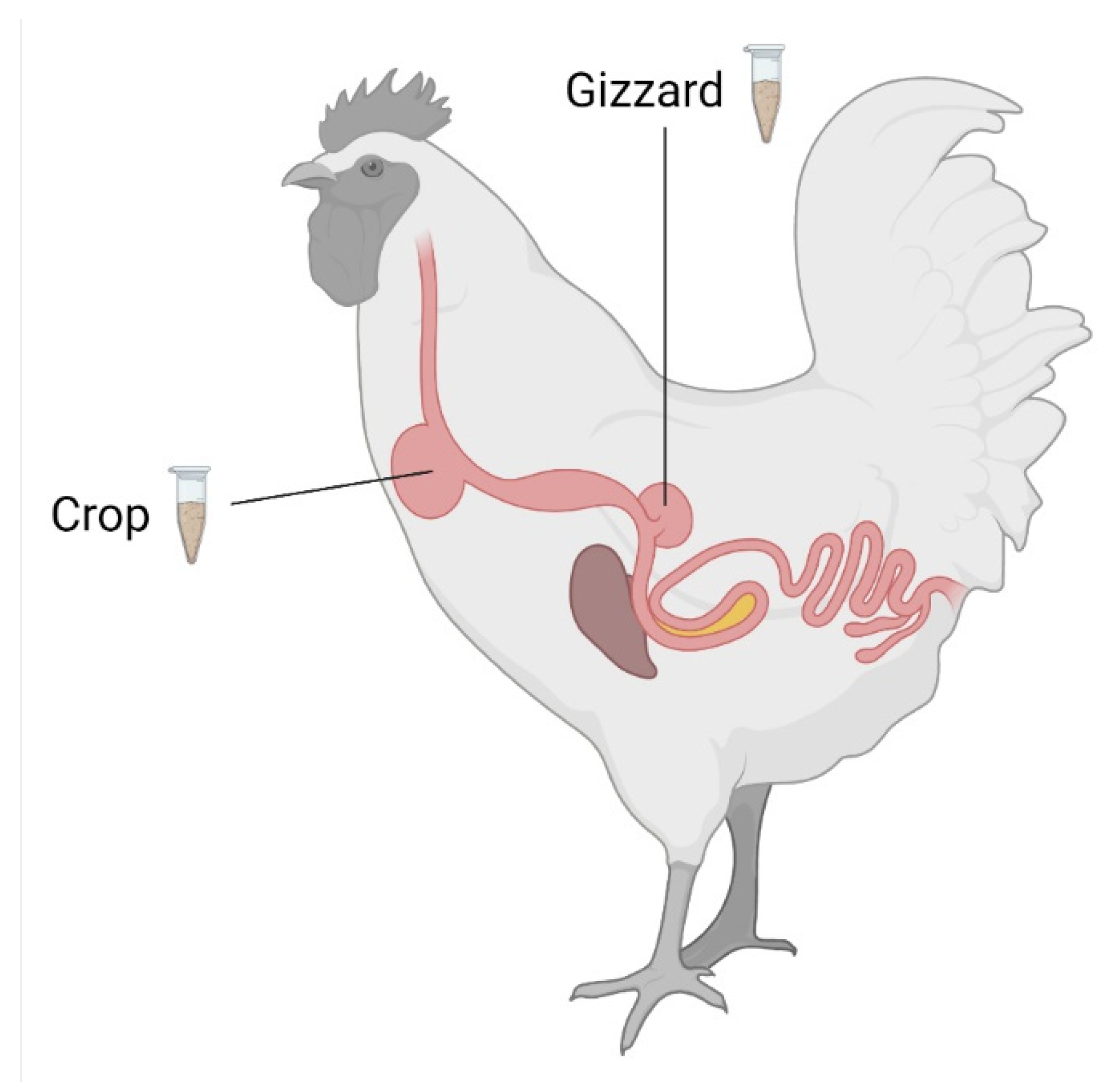
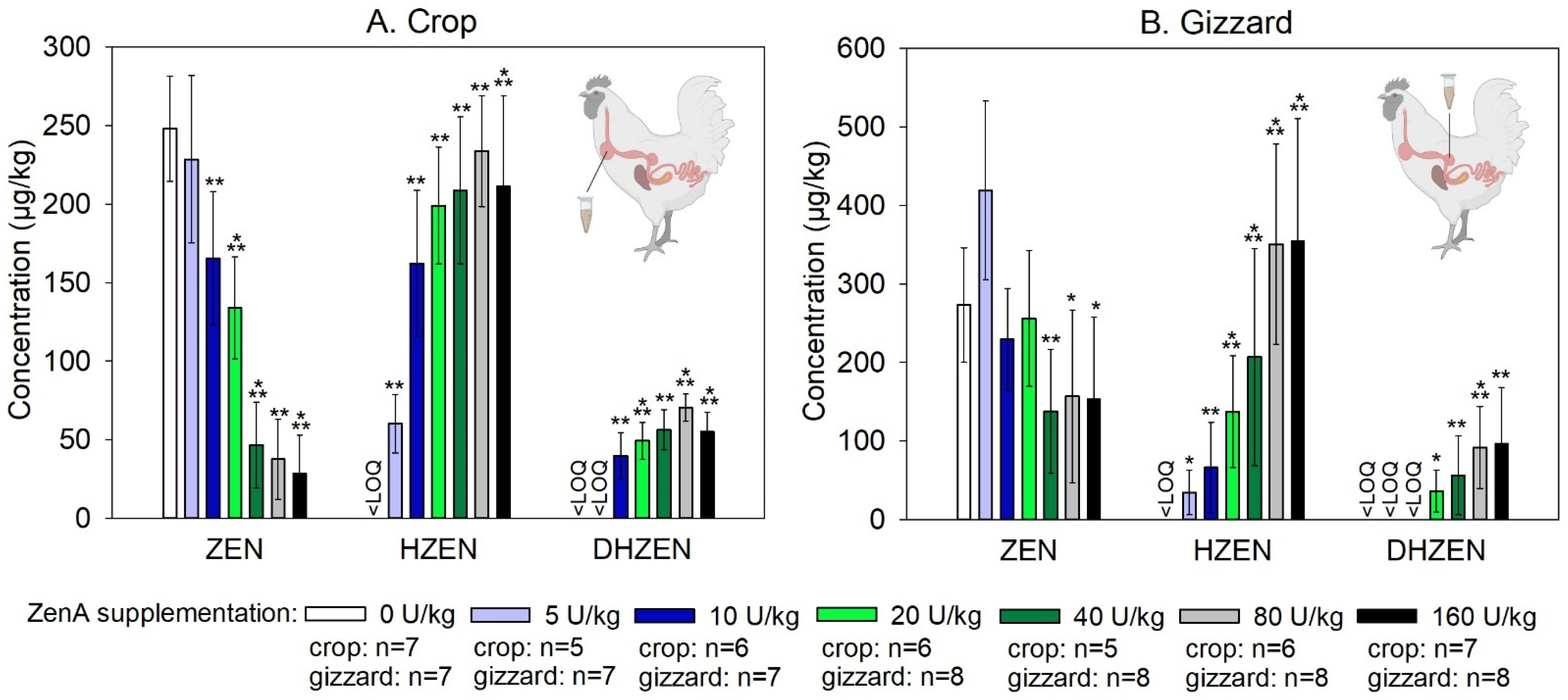
2.1. Efficacy of ZenA in Broiler Chickens
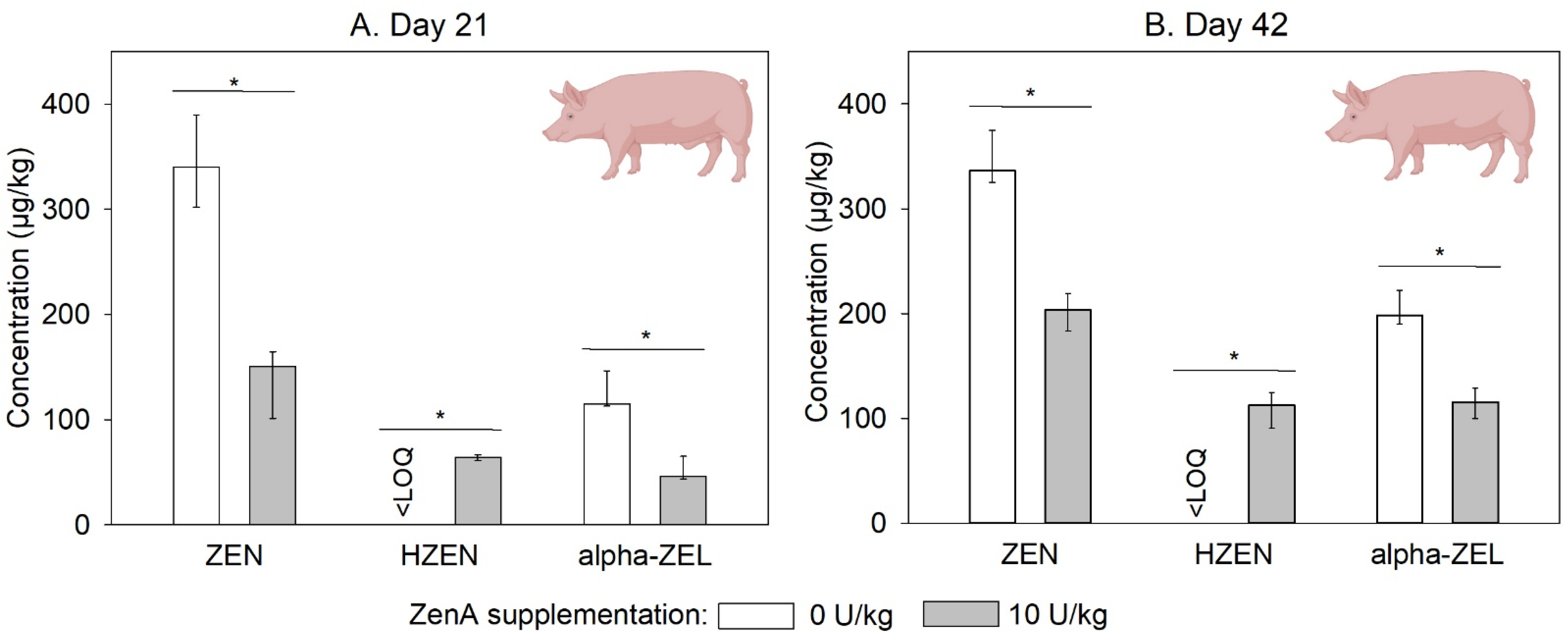
2.2. Efficacy of ZenA in Pigs
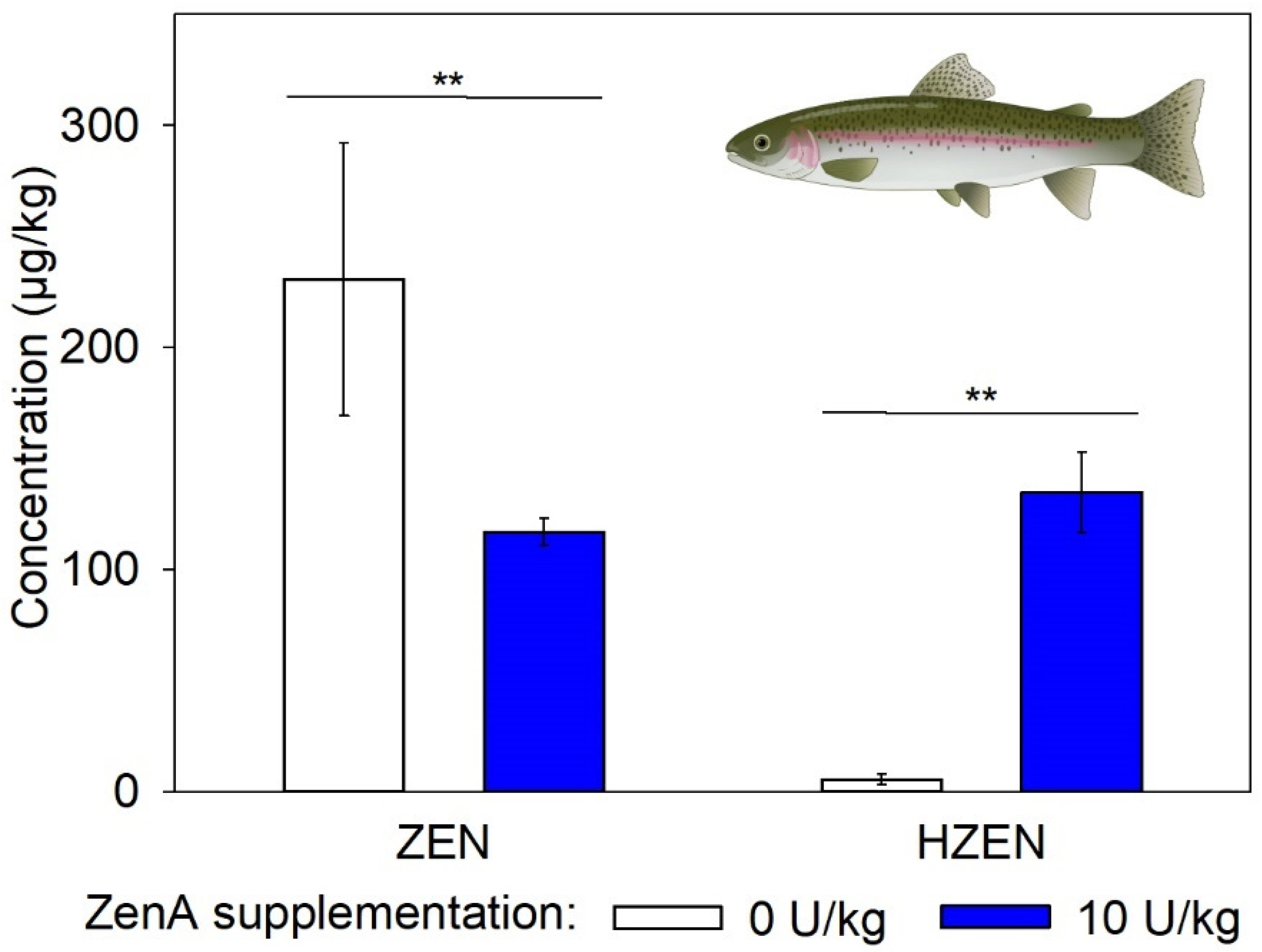
2.3. Efficacy of ZenA in Rainbow Trout
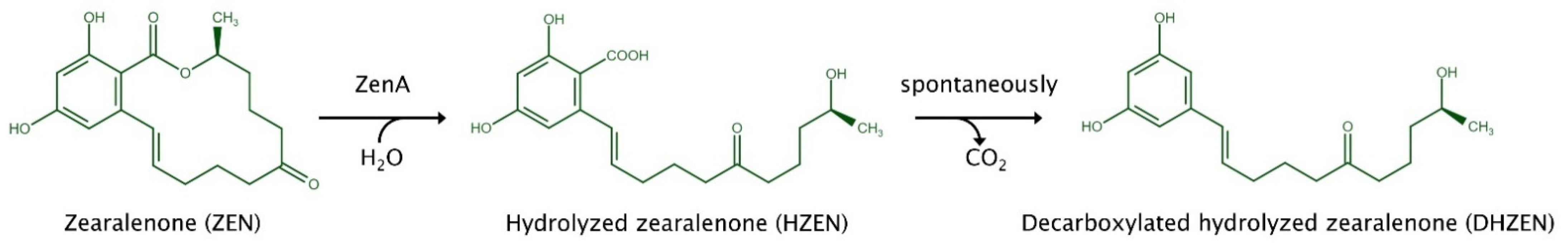
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Standards
5.2. Broiler Chicken Feeding Trial
5.2.1. Experimental Setup
5.2.2. Diet
5.2.3. Preparation of ZEN-Contaminated and ZenA-Supplemented Feed
5.2.4. Sampling and Sample Preparation
5.2.5. Analysis of ZEN and its Metabolites in Digesta and Analysis of ZEN in Feed
5.3. Pig Feeding Trial
5.3.1. Experimental Setup
5.3.2. Diet
5.3.3. Preparation of ZEN-Contaminated and ZenA-Supplemented Feed
5.3.4. Sampling and Sample Preparation
5.3.5. Analysis of ZEN, HZEN, DHZEN, and α-ZEL in Feces
5.4. Fish Feeding Trial
5.4.1. Experimental Setup
5.4.2. Diet
5.4.3. Preparation of ZEN-Contaminated and ZenA-Supplemented Feed
- (A)
- In a first step, sorbent was added to the pellets. To this end, 2928 g pellets were placed in a drum mixer (63 l, 27.5 rpm; VidaXL, Limburg, The Netherlands), and rotation was started. In total, 100 g of colloidal silica (Aerosil® 200; Evonik Operations GmbH, Essen, Germany) were gradually added to the rotating pellets, and the mixture was stirred for 3 min until a uniform adhesive layer formed on the pellet surface.
- (B)
- For each experimental group, a separate coating dispersion was prepared.
- (C)
- For pan coating, the coating dispersion (B) was gradually poured onto the pellets with adhered sorbent (A) over a period of 10 min. After coating, pellets were distributed in plastic trays in 2 cm layers and dried in a laboratory incubator (Binder BD 240; Binder GmbH, Tuttlingen, Germany) with fan-assisted forced air circulation for 24 h. Samples were dried at 45 °C with 60 rpm fan speed and completely opened an air flap to maximize air exchange.
5.4.4. Sampling and Sample Preparation
5.4.5. Analysis of ZEN, HZEN, and DHZEN in Digesta
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2019, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Cortiñas Abrahantes, J.; Dorne, J.-L.; Battilani, P.; Toscano, P. Occurrence and co-occurrence of mycotoxins in cereal-based feed and food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Metzler, M.; Pfeiffer, E.; Hildebrand, A. Zearalenone and its metabolites as endocrine disrupting chemicals. World Mycotoxin J. 2010, 3, 385–401. [Google Scholar] [CrossRef]
- Tiemann, U.; Dänicke, S. In vivo and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs: A review. Food Addit. Contam. 2007, 24, 306–314. [Google Scholar] [CrossRef]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and Reproductive Function in Farm Animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef]
- Braicu, C.; Cojocneanu-Petric, R.; Jurj, A.; Gulei, D.; Taranu, I.; Gras, A.M.; Marin, D.E.; Berindan-Neagoe, I. Microarray based gene expression analysis of Sus Scrofa duodenum exposed to zearalenone: Significance to human health. BMC Genom. 2016, 17, 646. [Google Scholar] [CrossRef]
- Obremski, K.; Gonkowski, S.; Wojtacha, P. Zearalenone-induced changes in the lymphoid tissue and mucosal nerve fibers in the porcine ileum. Pol. J. Vet. Sci. 2015, 18, 357–365. [Google Scholar] [CrossRef]
- Obremski, K.; Poniatowska-Broniek, G. Zearalenone induces apoptosis and inhibits proliferation in porcine ileal Peyer’s patch lymphocytes. Pol. J. Vet. Sci. 2015, 18, 153–161. [Google Scholar] [CrossRef]
- Obremski, K.; Wojtacha, P.; Podlasz, P.; Żmigrodzka, M. The influence of experimental administration of low zearalenone doses on the expression of Th1 and Th2 cytokines and on selected subpopulations of lymphocytes in intestinal lymph nodes. Pol. J. Vet. Sci. 2015, 18, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.S.; Mirocha, C.J.; Weaver, G.A.; Kurtz, H.J. Effect of zearalenone on female White Leghorn chickens. Appl. Environ. Microbiol. 1980, 39, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Maryamma, K.I.; Manomohan, C.B.; Nair, M.G.; Ismail, P.K.; Sreekumaran, T.; Rajan, A. Pathology of zearalenone toxicosis in chicken and evaluation of zearalenone residues in tissues. Indian J. Anim. Sci. 1992, 62, 105–107. [Google Scholar]
- Allen, N.K.; Mirocha, C.J.; Weaver, G.; Aakhus-Allen, S.A.; Bates, F. Effects of Dietary Zearalenone on Finishing Broiler Chickens and Young Turkey Poults. Poult. Sci. 1981, 60, 124–131. [Google Scholar] [CrossRef] [PubMed]
- European Commission (EC). Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off. J. Eur. Union 2006, L 229, 7–9. [Google Scholar]
- Woźny, M.; Dobosz, S.; Hliwa, P.; Gomułka, P.; Król, J.; Obremski, K.; Blahova, J.; Svobodova, Z.; Michalik, O.; Ocalewicz, K.; et al. Feed-borne exposure to zearalenone impairs reproduction of rainbow trout. Aquaculture 2020, 528, 735522. [Google Scholar] [CrossRef]
- Woźny, M.; Obremski, K.; Hliwa, P.; Gomułka, P.; Różyński, R.; Wojtacha, P.; Florczyk, M.; Segner, H.; Brzuzan, P. Feed contamination with zearalenone promotes growth but affects the immune system of rainbow trout. Fish Shellfish Immunol. 2018, 84, 680–694. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed. Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed. Sci. Technol. 2007, 137, 342–362. [Google Scholar] [CrossRef]
- Palumbo, R.; Gonçalves, A.; Gkrillas, A.; Logrieco, A.; Dorne, J.-L.; Dall’Asta, C.; Venâncio, A.; Battilani, P. Mycotoxins in maize: Mitigation actions, with a chain management approach. Phytopathol. Mediterr. 2020, 59, 5–28. [Google Scholar] [CrossRef]
- Phillips, T.D.; Wang, M.; Elmore, S.E.; Hearon, S.; Wang, J.-S. NovaSil Clay for the Protection of Humans and Animals from Aflatoxins and Other Contaminants. Clays Clay Miner. 2019, 67, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, M.C.; De Neeff, D.V.; Jager, A.V.; Corassin, C.H.; de Pinho Carão, Á.C.; de Albuquerque, R.; Azevedo, A.C.; Oliveira, C.A.F. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Elliott, C.T.; Connolly, L.; Kolawole, O. Potential adverse effects on animal health and performance caused by the addition of mineral adsorbents to feeds to reduce mycotoxin exposure. Mycotoxin Res. 2020, 36, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and Methodologies for Developing Microbial Detoxification Systems to Mitigate Mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mule, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef]
- Fruhauf, S.; Novak, B.; Nagl, V.; Hackl, M.; Hartinger, D.; Rainer, V.; Labudová, S.; Adam, G.; Aleschko, M.; Moll, W.-D.; et al. Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo. Toxins 2019, 11, 481. [Google Scholar] [CrossRef]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the Mycotoxin, Zearalenone, to a Non-estrogenic Compound by a Fungal Strain of Clonostachy ssp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef]
- Vekiru, E.; Frühauf, S.; Hametner, C.; Schatzmayr, G.; Krska, R.; Moll, W.D.; Schuhmacher, R. Isolation and characterisation of enzymatic zearalenone hydrolysis reaction products. World Mycotoxin J. 2016, 9, 353–363. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B.; Aleschko, M.; Stoiber, C.; Höbartner-Gußl, A.; Schöndorfer, K.; Killinger, M.; Zebeli, Q.; Schatzmayr, D. Metabolism of Zearalenone in the Rumen of Dairy Cows with and without Application of a Zearalenone-Degrading Enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef]
- Debevere, S.; Schatzmayr, D.; Reisinger, N.; Aleschko, M.; Haesaert, G.; Rychlik, M.; Croubels, S.; Fievez, V. Evaluation of the Efficacy of Mycotoxin Modifiers and Mycotoxin Binders by Using an In Vitro Rumen Model as a First Screening Tool. Toxins 2020, 12, 405. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.A.; Schatzmayr, D.; Hofstetter, U.; Santos, G.A. Occurrence of mycotoxins in aquaculture: Preliminary overview of Asian and European plant ingredients and finished feeds. World Mycotoxin J. 2017, 10, 183–194. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Edler, L.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on the appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar] [CrossRef]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef]
- Brezina, U.; Rempe, I.; Kersten, S.; Valenta, H.; Humpf, H.-U.; Dänicke, S. Determination of zearalenone, deoxynivalenol and metabolites in bile of piglets fed diets with graded levels of Fusarium toxin contaminated maize. World Mycotoxin J. 2016, 9, 179–193. [Google Scholar] [CrossRef]
- Dänicke, S.; Brüssow, K.-P.; Valenta, H.; Ueberschär, K.-H.; Tiemann, U.; Schollenberger, M. On the effects of graded levels of Fusarium toxin contaminated wheat in diets for gilts on feed intake, growth performance and metabolism of deoxynivalenol and zearalenone. Mol. Nutr. Food Res. 2005, 49, 932–943. [Google Scholar] [CrossRef]
- Brezina, U.; Rempe, I.; Kersten, S.; Valenta, H.; Humpf, H.-U.; Dänicke, S. Diagnosis of intoxications of piglets fed with Fusarium toxin-contaminated maize by the analysis of mycotoxin residues in serum, liquor and urine with LC-MS/MS. Arch. Anim. Nutr. 2014, 68, 425–447. [Google Scholar] [CrossRef]
- Olsen, M.E.; Pettersson, H.I.; Sandholm, K.A.; Kiessling, K.-H.C. Quantitative Liquid Chromatographic Method Using Fluorescence Detection for Determining Zearalenone and Its Metabolites in Blood Plasma and Urine. J. AOAC Int. 1985, 68, 632–635. [Google Scholar] [CrossRef]
- Focker, M.; van der Fels-Klerx, H.J.; Magan, N.; Edwards, S.G.; Grahovac, M.; Bagi, F.; Budakov, D.; Suman, M.; Schatzmayr, G.; Krska, R.; et al. The impact of management practices to prevent and control mycotoxins in the European food supply chain: MyToolBox project results. World Mycotoxin J. 2021, 14, 139–154. [Google Scholar] [CrossRef]
- Chang, X.; Liu, H.; Sun, J.; Wang, J.; Zhao, C.; Zhang, W.; Zhang, J.; Sun, C. Zearalenone Removal from Corn Oil by an Enzymatic Strategy. Toxins 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, L.; Rozhkova, A.; Osipov, D.; Zorov, I.; Mikityuk, O.; Statsyuk, N.; Sinitsyna, O.; Dzhavakhiya, V.; Sinitsyn, A. Effective Zearalenone Degradation in Model Solutions and Infected Wheat Grain Using a Novel Heterologous Lactonohydrolase Secreted by Recombinant Penicillium canescens. Toxins 2020, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Zhang, W.; Xiao, Z.; Zhang, D. Characterization, expression and application of a zearalenone degrading enzyme from Neurospora crassa. AMB Express 2018, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Higa-Nishiyama, A.; Takahashi-Ando, N.; Shimizu, T.; Kudo, T.; Yamaguchi, I.; Kimura, M. A Model Transgenic Cereal Plant with Detoxification Activity for the Estrogenic Mycotoxin Zearalenone. Transgenic Res. 2005, 14, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Igawa, T.; Takahashi-Ando, N.; Ochiai, N.; Ohsato, S.; Shimizu, T.; Kudo, T.; Yamaguchi, I.; Kimura, M. Reduced Contamination by the Fusarium Mycotoxin Zearalenone in Maize Kernels through Genetic Modification with a Detoxification Gene. Appl. Environ. Microbiol. 2007, 73, 1622–1629. [Google Scholar] [CrossRef]
- European Commission. Directive 2010/63/eu of the European Parliament and of the council of 22 september 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, L 276/33, 1–47. [Google Scholar]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of > 500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef]
- Dobsikova, R.; Blahova, J.; Franc, A.; Jakubik, J.; Mikulikova, I.; Modra, H.; Novotna, K.; Svobodova, Z. Effect of ?-1.3/1.6-D-glucan derived from oyster mushroom Pleurotus ostreatus on biometrical, haematological, biochemical, and immunological indices in rainbow trout (Oncorhynchus mykiss). Neuro Endocrinol. Lett. 2012, 33 (Suppl. S3), 96–106. [Google Scholar]
- Vishwanath, V.; Sulyok, M.; Labuda, R.; Bicker, W.; Krska, R. Simultaneous determination of 186 fungal and bacterial metabolites in indoor matrices by liquid chromatography/tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 395, 1355–1372. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group 1 | Total Feed Intake per Animal (Average) | Weight per Pen at Trial Start | Weight per Pen at Trial End |
|---|---|---|---|
| Negative control | 1.31 kg | 3.22 kg | 9.70 kg |
| Positive control | 1.28 kg | 3.17 kg | 9.70 kg |
| 5 U/kg ZenA | 1.31 kg | 3.18 kg | 9.62 kg |
| 10 U/kg ZenA | 1.37 kg | 3.17 kg | 9.48 kg |
| 20 U/kg ZenA | 1.31 kg | 3.15 kg | 9.40 kg |
| 40 U/kg ZenA | 1.33 kg | 3.13 kg | 9.69 kg |
| 80 U/kg ZenA | 1.24 kg | 3.17 kg | 9.22 kg |
| 160 U/kg ZenA | 1.35 kg | 3.14 kg | 9.99 kg |
| Treatment Group 1 | Total Feed Intake per Animal (Average) | Weight per Animal at Trial Start (Average) | Weight per Animal at Trial End (Average) |
|---|---|---|---|
| Negative control | 45.85 kg | 9.21 kg | 34.85 kg |
| Positive control | 46.16 kg | 9.21 kg | 34.95 kg |
| ZenA | 46.30 kg | 9.21 kg | 35.60 kg |
| Treatment Group 1 | Feed Intake per Animal per Day (Average) | Weight per Animal at Trial Start (Average) | Weight per Animal at Trial end (Average) |
|---|---|---|---|
| Negative control | 0.49 g | 9.8 g | 50.5 g |
| Positive control | 0.48 g | 9.8 g | 50.9 g |
| ZenA | 0.48 g | 9.8 g | 51.3 g |
| Ingredients | % |
|---|---|
| Maize | 62.00 |
| Soy 48% | 23.80 |
| Full-fat soy | 5.55 |
| Sunflower oil | 2.00 |
| Monocalcium phosphate | 1.70 |
| Calcium carbonate | 1.55 |
| Fat powder | 1.50 |
| Pumpkin cake | 0.58 |
| Sodium bicarbonate | 0.28 |
| L-Lysine | 0.27 |
| DL-Methionine | 0.20 |
| Sodium chloride | 0.18 |
| Magnesium phosphate | 0.10 |
| L-Threonine | 0.10 |
| Cholinchloride | 0.07 |
| Vitamin and trace element premix 1 | 0.12 |
| Analyzed composition | |
| Dry matter (%) | 88.1 |
| Crude protein (%) | 18.3 |
| Crude fiber (%) | 2.6 |
| Crude fat (%) | 7.5 |
| Neutral detergent fiber (%) | 8.7 |
| Non-fiber carbohydrates (%) | 47.5 |
| N-free extract (%) | 53.6 |
| Starch (%) | 38.2 |
| Crude ash (%) | 6.1 |
| Metabolizable energy (MJ) | 11.83 |
| Analyte 1 | Q1 Mass (m/z) | Q3 Mass (m/z) 2 | Declustering Potential (V) | Collision Energy (V) 2 |
|---|---|---|---|---|
| ZEN | 317.1 | 131.0/175.0 | −120 | −42/−34 |
| α-ZEL | 319.1 | 275.1/160.0 | −125 | −30/−42 |
| HZEN | 335.0 | 149.0/161.0 | −100 | −34/−34 |
| DHZEN | 291.1 | 149.1/161.1 | −100 | −25/−25 |
| Ingredients | % |
|---|---|
| Wheat | 60.00 |
| Soy 48% | 20.00 |
| Barley | 14.00 |
| Calcium carbonate | 1.20 |
| Wheat bran | 1.00 |
| Monocalcium phosphate | 0.90 |
| Vinasse | 0.80 |
| Sodium chloride | 0.45 |
| Rye bran | 0.40 |
| L-Lysine | 0.25 |
| Rapeseed oil | 0.20 |
| Magnesium phosphate | 0.20 |
| DL-Methionine | 0.19 |
| L-Threonine | 0.10 |
| Tryptophan | 0.07 |
| Trace element and Vitamin premix 1 | 0.30 |
| Analyzed composition | |
| Dry matter (%) | 87.4 |
| Crude protein (%) | 16.9 |
| Crude fat (%) | 6.4 |
| Crude fiber (%) | 3.0 |
| N-free extract (%) | 53.5 |
| Crude ash (%) | 7.4 |
| Starch (%) | 35.8 |
| Metabolizable energy (MJ) | 13.37 |
| Analyzed composition of 1.5 mm pellets | |
| Dry matter (g/kg) | 939 |
| Crude protein (g/kg) | 469 |
| Crude fat (g/kg) | 229 |
| Crude fiber (g/kg) | 6 |
| Crude ash (g/kg) | 61 |
| Starch (g/kg) | 145 |
| Analyzed composition of 2.5 mm pellets | |
| Dry matter (g/kg) | 929 |
| Crude protein (g/kg) | 589 |
| Crude fat (g/kg) | 140 |
| Crude fiber (g/kg) | 6 |
| Crude ash (g/kg) | 85 |
| Starch (g/kg) | 105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruber-Dorninger, C.; Killinger, M.; Höbartner-Gußl, A.; Rosen, R.; Doupovec, B.; Aleschko, M.; Schwartz-Zimmermann, H.; Greitbauer, O.; Marković, Z.; Stanković, M.; et al. Enzymatic Degradation of Zearalenone in the Gastrointestinal Tract of Pigs, Chickens, and Rainbow Trout. Toxins 2023, 15, 48. https://doi.org/10.3390/toxins15010048
Gruber-Dorninger C, Killinger M, Höbartner-Gußl A, Rosen R, Doupovec B, Aleschko M, Schwartz-Zimmermann H, Greitbauer O, Marković Z, Stanković M, et al. Enzymatic Degradation of Zearalenone in the Gastrointestinal Tract of Pigs, Chickens, and Rainbow Trout. Toxins. 2023; 15(1):48. https://doi.org/10.3390/toxins15010048
Chicago/Turabian StyleGruber-Dorninger, Christiane, Manuela Killinger, Andreas Höbartner-Gußl, Roy Rosen, Barbara Doupovec, Markus Aleschko, Heidi Schwartz-Zimmermann, Oliver Greitbauer, Zoran Marković, Marko Stanković, and et al. 2023. "Enzymatic Degradation of Zearalenone in the Gastrointestinal Tract of Pigs, Chickens, and Rainbow Trout" Toxins 15, no. 1: 48. https://doi.org/10.3390/toxins15010048
APA StyleGruber-Dorninger, C., Killinger, M., Höbartner-Gußl, A., Rosen, R., Doupovec, B., Aleschko, M., Schwartz-Zimmermann, H., Greitbauer, O., Marković, Z., Stanković, M., Schöndorfer, K., Vukmirovic, D., Wein, S., & Schatzmayr, D. (2023). Enzymatic Degradation of Zearalenone in the Gastrointestinal Tract of Pigs, Chickens, and Rainbow Trout. Toxins, 15(1), 48. https://doi.org/10.3390/toxins15010048