Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors

 ,
,  , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
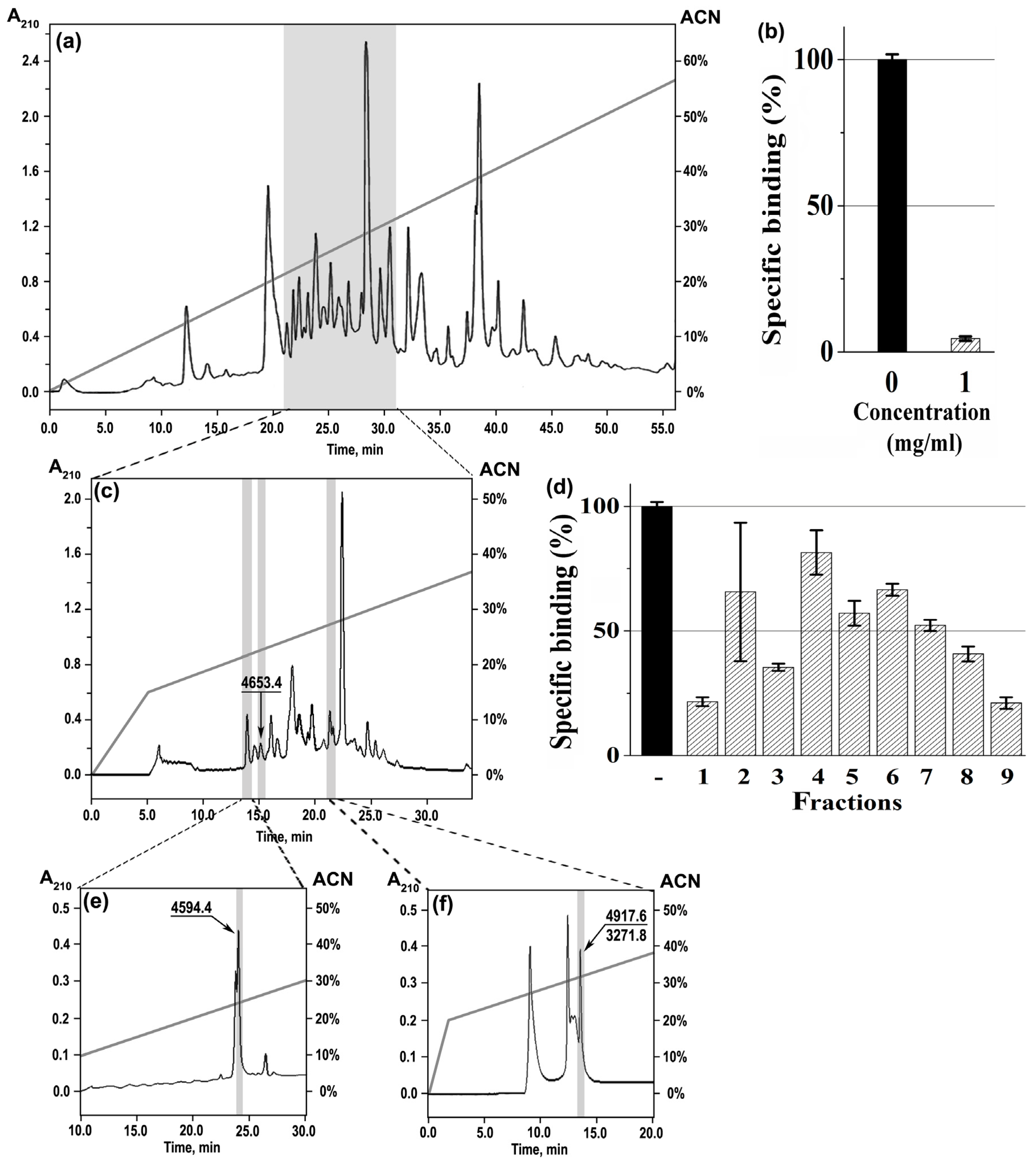
2.1. Isolation of nAChR-Targeting Peptides and Their Primary Structure Determination
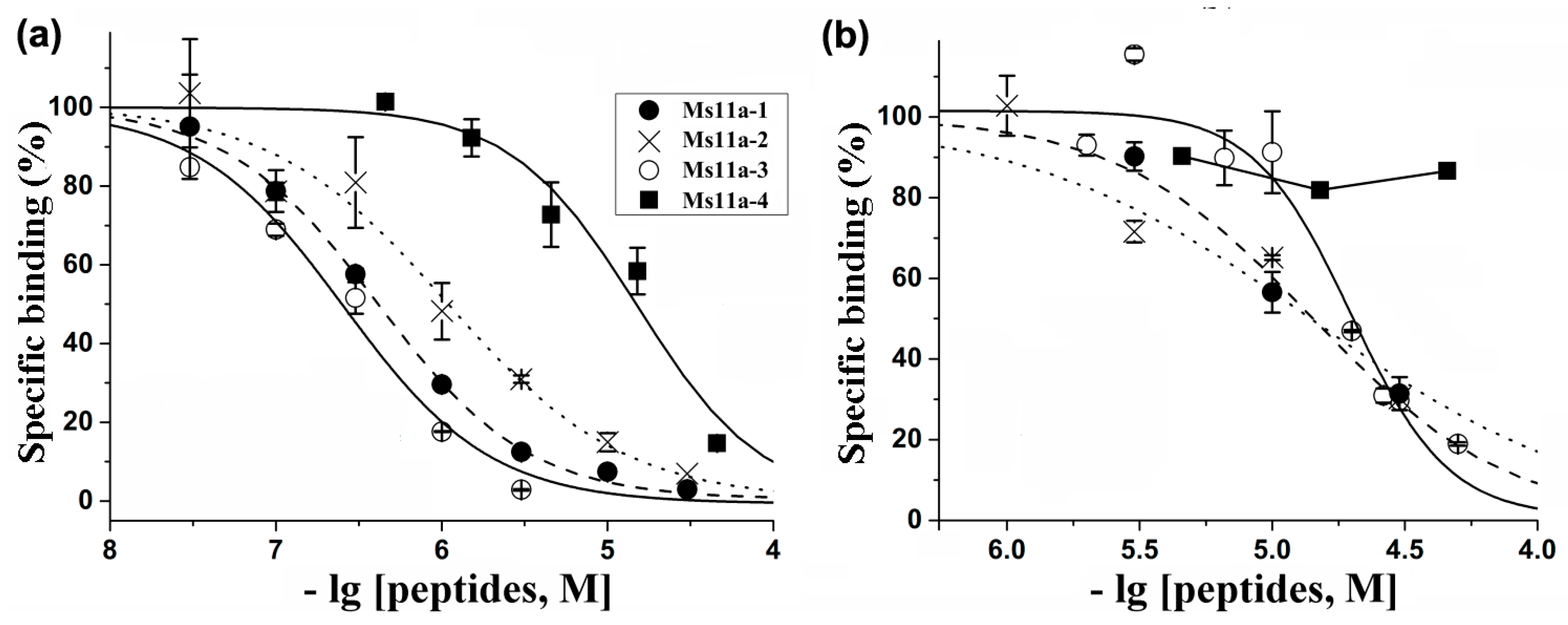
2.2. Production of Recombinant Peptides and Their Binding and Functional Activity
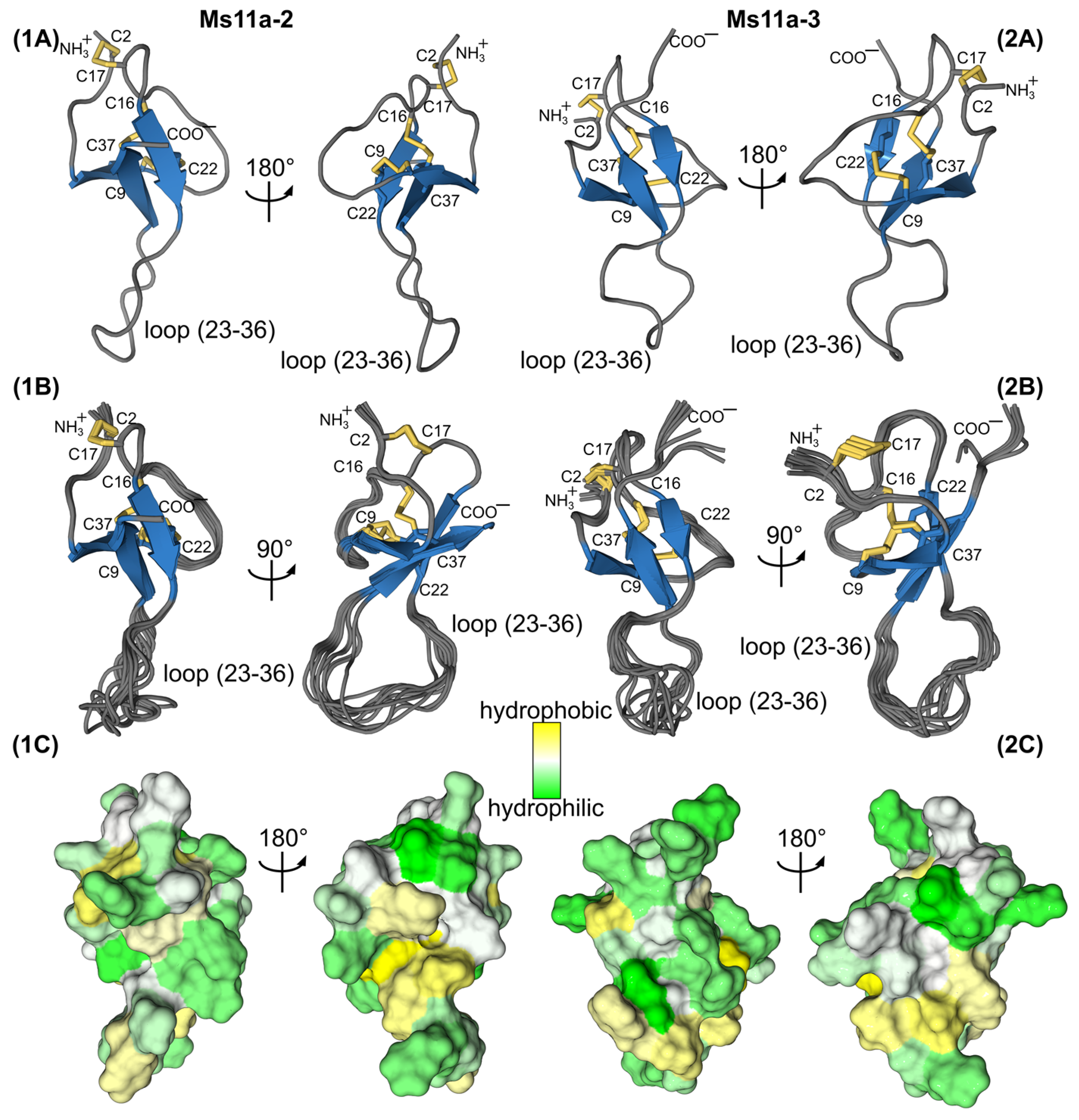
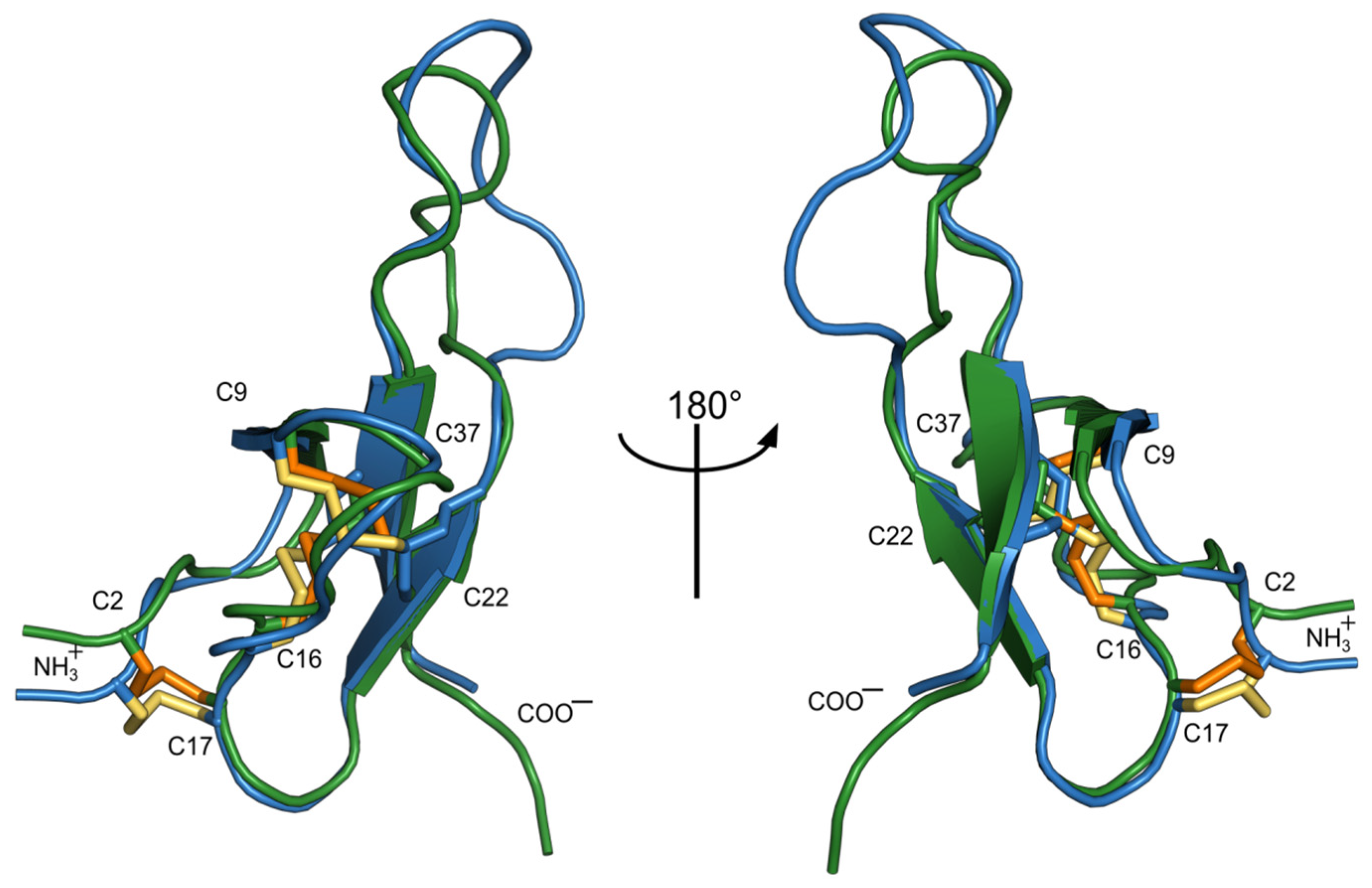
2.3. NMR Study of Recombinant Sea Anemone Peptides
3. Discussion
4. Conclusions
5. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quesada, A.J.; Acuña, F.H.; Cortés, J. Diet of the sea anemone Anthopleura nigrescens: Composition and variation between daytime and nighttime high tides. Zool. Stud. 2014, 53, 1–7. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed]
- Shick, J.M. A Functional Biology of Sea Anemones; Springer: Berlin, Germany, 1991. [Google Scholar] [CrossRef]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Lotan, A.; Fishman, L.; Loya, Y.; Zlotkin, E. Delivery of a nematocyst toxin. Nature 1995, 375, 456. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef]
- Menezes, C.; Thakur, N.L. Sea anemone venom: Ecological interactions and bioactive potential. Toxicon 2022, 208, 31–46. [Google Scholar] [CrossRef]
- Van Iten, H.; Marques, A.C.; de Leme, J.M.; Pacheco, M.L.A.F.; Simões, M.G. Origin and early diversification of the phylum Cnidaria Verrill: Major developments in the analysis of the taxon’s Proterozoic–Cambrian history. Palaeontology 2014, 57, 677–690. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea anemone peptide with uncommon β-hairpin structure inhibits acid-sensing ion channel 3 (ASIC3) and reveals analgesic activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef]
- Mann, P.C.; Vahle, J.; Keenan, C.M.; Baker, J.F.; Bradley, A.E.; Goodman, D.G.; Harada, T.; Herbert, R.; Kaufmann, W.; Kellner, R.; et al. International Harmonization of Toxicologic Pathology Nomenclature: An Overview and Review of Basic Principles. Toxicol. Pathol. 2012, 40, 7S–13S. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.A.; Salceda, E.; Garateix, A.G.; Zaharenko, A.J.; Peigneur, S.; López, O.; Pons, T.; Richardson, M.; Díaz, M.; Hernández, Y.; et al. A novel sea anemone peptide that inhibits acid-sensing ion channels. Peptides 2014, 53, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Kalina, R.S.; Koshelev, S.G.; Zelepuga, E.A.; Kim, N.Y.; Kozlov, S.A.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; Gladkikh, I.N. APETx-like peptides from the sea anemone Heteractis crispa, diverse in their effect on ASIC1a and ASIC3 ion channels. Toxins 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Philyppov, I.B.; Paduraru, O.N.; Andreev, Y.A.; Grishin, E.V.; Shuba, Y.M. Modulation of TRPV1-dependent contractility of normal and diabetic bladder smooth muscle by analgesic toxins from sea anemone Heteractis crispa. Life Sci. 2012, 91, 912–920. [Google Scholar] [CrossRef]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvåg, K.; et al. Peptide from Sea Anemone Metridium senile Affects Transient Receptor Potential Ankyrin-repeat 1 (TRPA1) Function and Produces Analgesic Effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [PubMed]
- Logashina, Y.A.; Solstad, R.G.; Mineev, K.S.; Korolkova, Y.V.; Mosharova, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Arseniev, A.S.; et al. New Disulfide-Stabilized Fold Provides Sea Anemone Peptide to Exhibit Both Antimicrobial and TRPA1 Potentiating Properties. Toxins 2017, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Lee, Y.J.; Go, H.J.; Oh, H.Y.; Lee, T.K.; Park, J.B.; Park, N.G. Defensin-neurotoxin dyad in a basally branching metazoan sea anemone. FEBS J. 2017, 284, 3320–3338. [Google Scholar] [CrossRef]
- Grafskaia, E.N.; Polina, N.F.; Babenko, V.V.; Kharlampieva, D.D.; Bobrovsky, P.A.; Manuvera, V.A.; Farafonova, T.E.; Anikanov, N.A.; Lazarev, V.N. Discovery of novel antimicrobial peptides: A transcriptomic study of the sea anemone Cnidopus japonicus. J. Bioinform. Comput. Biol. 2018, 16, 1840006. [Google Scholar] [CrossRef]
- Antuch, W.; Berndt, K.D.; Chavez, M.A.; Delfin, J.; Wuthrich, K. The NMR solution structure of a Kunitz-type proteinase inhibitor from the sea anemone Stichodactyla helianthus. Eur. J. Biochem. 1993, 212, 675–684. [Google Scholar] [CrossRef]
- Tysoe, C.; Williams, L.K.; Keyzers, R.; Nguyen, N.T.; Tarling, C.; Wicki, J.; Goddard-Borger, E.D.; Aguda, A.H.; Perry, S.; Foster, L.J.; et al. Potent human alpha;-amylase inhibition by the β-defensin-like protein helianthamide. ACS Cent. Sci. 2016, 2, 154–161. [Google Scholar] [CrossRef]
- Tysoe, C.; Withers, S.G. Structural Dissection of Helianthamide Reveals the Basis of Its Potent Inhibition of Human Pancreatic α-Amylase. Biochemistry 2018, 57, 5384–5387. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.; Gladkikh, I.; Kalinovskii, A.; Zelepuga, E.; Monastyrnaya, M.; Kim, N.; Shevchenko, L.; Peigneur, S.; Tytgat, J.; Kozlovskaya, E.; et al. Magnificamide, a β-Defensin-like peptide from the mucus of the sea anemone heteractis magnifica, is a strong inhibitor of Mammalian α-Amylases. Mar. Drugs 2019, 17, 542. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Clementi, F.; Fornari, A.; Gaimarri, A.; Guiducci, S.; Manfredi, I.; Moretti, M.; Pedrazzi, P.; Pucci, L.; Zoli, M. Structural and functional diversity of native brain neuronal nicotinic receptors. Biochem. Pharmacol. 2009, 78, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Dineley, K.T.; Pandya, A.A.; Yakel, J.L. Nicotinic ACh receptors as therapeutic targets in CNS disorders. Trends Pharmacol. Sci. 2015, 36, 96–108. [Google Scholar] [CrossRef]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef]
- Abraham, N.; Lewis, R.J. Neuronal Nicotinic Acetylcholine Receptor Modulators from Cone Snails. Mar. Drugs 2018, 16, 208. [Google Scholar] [CrossRef]
- Kasheverov, I.E.; Oparin, P.B.; Zhmak, M.N.; Egorova, N.S.; Ivanov, I.A.; Gigolaev, A.M.; Nekrasova, O.V.; Serebryakova, M.V.; Kudryavtsev, D.S.; Prokopev, N.A.; et al. Scorpion toxins interact with nicotinic acetylcholine receptors. FEBS Lett. 2019, 593, 2779–2789. [Google Scholar] [CrossRef]
- Kozlov, S.; Grishin, E. Convenient nomenclature of cysteine-rich polypeptide toxins from sea anemones. Peptides 2012, 33, 240–244. [Google Scholar] [CrossRef]
- Wimley, W.C.; White, S.H. Experimentally determined hydrophobicity scale for proteins at membrane interfaces. Nat. Struct. Biol. 1996, 3, 842–848. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef]
- de Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.B.; Hurwood, D.A.; Prentis, P.J. Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef] [PubMed]
- Nirthanan, S. Snake three-finger α-neurotoxins and nicotinic acetylcholine receptors: Molecules, mechanisms and medicine. Biochem. Pharmacol. 2020, 181, 114168. [Google Scholar] [CrossRef]
- Chambers, C.; Cutler, P.; Huang, Y.H.; Goodchild, J.A.; Blythe, J.; Wang, C.K.; Bigot, A.; Kaas, Q.; Craik, D.J.; Sabbadin, D.; et al. Insecticidal spider toxins are high affinity positive allosteric modulators of the nicotinic acetylcholine receptor. FEBS Lett. 2019, 593, 1336–1350. [Google Scholar] [CrossRef]
- Pan, N.C.; Zhang, T.; Hu, S.; Liu, C.; Wang, Y. Fast desensitization of acetylcholine receptors induced by a spider toxin. Channels 2021, 15, 507–515. [Google Scholar] [CrossRef]
- Kasheverov, I.E.; Zhmak, M.N.; Vulfius, C.A.; Gorbacheva, E.V.; Mordvintsev, D.Y.; Utkin, Y.N.; Van Elk, R.; Smit, A.B.; Tsetlin, V.I. Alpha-conotoxin analogs with additional positive charge show increased selectivity towards Torpedo californica and some neuronal subtypes of nicotinic acetylcholine receptors. FEBS J. 2006, 273, 4470–4481. [Google Scholar] [CrossRef]
- Ellison, M.; Haberlandt, C.; Gomez-Casati, M.E.; Watkins, M.; Elgoyhen, A.B.; McIntosh, J.M.; Olivera, B.M. α-RgIA: A novel conotoxin that specifically and potently blocks the α9α10 nAChR. Biochemistry 2006, 45, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kaas, Q.; Wu, Y.; Zhu, X.; Hu, Y.; Li, X.; Tsetlin, V.I.; Christensen, S.; et al. Cloning, synthesis, and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, E4026–E4035. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Secondary-structure matching (SSM), a new tool for fast protein structure alignment in three dimensions. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2256–2268. [Google Scholar] [CrossRef]
- Windley, M.J.; Vetter, I.; Lewis, R.J.; Nicholson, G.M. Lethal effects of an insecticidal spider venom peptide involve positive allosteric modulation of insect nicotinic acetylcholine receptors. Neuropharmacology 2017, 127, 224–242. [Google Scholar] [CrossRef]
- Orts, D.J.B.; Moran, Y.; Cologna, C.T.; Peigneur, S.; Madio, B.; Praher, D.; Quinton, L.; De Pauw, E.; Bicudo, J.E.P.W.; Tytgat, J.; et al. BcsTx3 is a founder of a novel sea anemone toxin family of potassium channel blocker. FEBS J. 2013, 280, 4839–4852. [Google Scholar] [CrossRef] [PubMed]
- Jacobsson, E.; Andersson, H.S.; Strand, M.; Peigneur, S.; Eriksson, C.; Lodén, H.; Shariatgorji, M.; Andrén, P.E.; Lebbe, E.K.M.; Rosengren, K.J.; et al. Peptide ion channel toxins from the bootlace worm, the longest animal on Earth. Sci. Rep. 2018, 8, 4596. [Google Scholar] [CrossRef] [PubMed]
- Jacobsson, E.; Peigneur, S.; Andersson, H.S.; Laborde, Q.; Strand, M.; Tytgat, J.; Göransson, U. Functional Characterization of the Nemertide α Family of Peptide Toxins. J. Nat. Prod. 2021, 84, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Marder, E. Cholinergic motor neurones in the stomatogastric system of the lobster. J. Physiol. 1976, 257, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Malloy, C.A.; Somasundaram, E.; Omar, A.; Bhutto, U.; Medley, M.; Dzubuk, N.; Cooper, R.L. Pharmacological identification of cholinergic receptor subtypes: Modulation of locomotion and neural circuit excitability in Drosophila larvae. Neuroscience 2019, 411, 47–64. [Google Scholar] [CrossRef]
- Sebens, K.P.; Koehl, M.A.R. Predation on zooplankton by the benthic anthozoans Alcyonium siderium (Alcyonacea) and Metridium senile (Actiniaria) in the New England subtidal. Mar. Biol. 1984, 81, 255–271. [Google Scholar] [CrossRef]
- Purcell, J.E. The diet of large and small individuals of the sea anemone Metridium senile. Bull. S. Calif. Acad. Sci. 1977, 76, 168–172. [Google Scholar]
- Wilhelm, E.; Bückmann, D.; Tomaschko, K.H. Life cycle and population dynamics of Pycnogonum litorale (Pycnogonida) in a natural habitat. Mar. Biol. 1997, 129, 601–606. [Google Scholar] [CrossRef]
- Harris, L.G. Size-selective predation in a sea anemone, nudi-branch, and fish food chain. Veliger 1986, 29, 38–47. [Google Scholar]
- Perron, F. the Habitat and Feeding Behavior of the Wentletrap Epitonium Greenlandicum. Malacologia 1978, 17, 63–72. [Google Scholar]
- Mattacola, A.D. An unusual diet for bream. J. Mar. Biol. Assoc. 1976, 56, 810. [Google Scholar] [CrossRef]
- Keats, D.W. Food of winter flounder Pseudopleuronectes americanus in a sea urchin dominated community in eastern Newfoundland. Mar. Ecol. Prog. Ser. 1990, 60, 13–22. [Google Scholar] [CrossRef]
- Greenwood, P.G.; Garry, K.; Hunter, A.; Jennings, M. Adaptable defense: A nudibranch mucus inhibits nematocyst discharge and changes with prey type. Biol. Bull. 2004, 206, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Elgoyhen, A.B. The α9α10 nicotinic acetylcholine receptor: A compelling drug target for hearing loss? Expert Opin. Ther. Targets 2022, 26, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Andreev, Y.A.; Kozlov, S.A.; Vassilevski, A.A.; Grishin, E.V. Cyanogen bromide cleavage of proteins in salt and buffer solutions. Anal. Biochem. 2010, 407, 144–146. [Google Scholar] [CrossRef]
- Zhu, X.; Pan, S.; Xu, M.; Zhang, L.; Yu, J.; Yu, J.; Wu, Y.; Fan, Y.; Li, H.; Kasheverov, I.E.; et al. High Selectivity of an α-Conotoxin LvIA Analogue for α3β2 Nicotinic Acetylcholine Receptors Is Mediated by β2 Functionally Important Residues. J. Med. Chem. 2020, 63, 13656–13668. [Google Scholar] [CrossRef]
- Wüthrich, K. NMR with Proteins and Nucleic Acids. Europhys. News 1986, 17, 11–13. [Google Scholar] [CrossRef]
- Herrmann, T.; Güntert, P.; Wüthrich, K. Protein NMR structure determination with automated NOE assignment using the new software CANDID and the torsion angle dynamics algorithm DYANA. J. Mol. Biol. 2002, 319, 209–227. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | [125I]α-Bgt-Competitive Binding Assay | |||
|---|---|---|---|---|
| T. californica α1β1γδ nAChR | Human α7 nAChR | |||
| IC50 (μM) | Hill Slope | IC50 (μM) | Hill Slope | |
| Ms11a-1 | 0.408 (0.395–0.421) | 0.96 (0.94–0.99) | 14.16 (13.48–14.87) | 1.18 (1.11–1.25) |
| Ms11a-2 | 1.080 (0.947–12.28) | 0.82 (0.75–0.89) | 14.13 (12.43–16.07) | 0.80 (0.69–0.91) |
| Ms11a-3 | 0.256 (0.236–0.277) | 0.98 (0.91–1.05) | 19.81 (18.44–21.28) | 2.37 (2.00–2.75) |
| Ms11a-4 | 14.95 (13.46–16.61) | 1.14 (1.02–1.26) | >45 | - |
| nAChR Subtype a | IC50 (μM) | Hill Slope |
|---|---|---|
| mα1β1δε | 1.215 (0.904–1.633) | 1.35 (0.71–1.99) |
| rα2β2 | na b | - |
| rα2β4 | na b | - |
| rα3β2 | na b | - |
| rα3β4 | 5.173 (4.19–6.388) | 0.84 (0.67–1.00) |
| rα4β2 | na b | - |
| rα4β4 | na b | - |
| rα6/α3β4 | na b | - |
| rα7 | 4.786 (3.055–7.497) | 0.99 (0.59–1.40) |
| hα7 | 8.869 (5.218–15.08) | 0.95 (0.30–1.61) |
| rα9α10 | 0.202 (0.141–0.29) | 0.71 (0.52–0.90) |
| Peptide | Ms11a-2 | Ms11a-3 |
|---|---|---|
| PDB ID | 6XYH | 6XYI |
| Distance and angle restraints | ||
| Total NOEs | 467 | 558 |
| intraresidual | 102 | 267 |
| interresidual | 365 | 291 |
| sequential (|i-j| = 1) | 110 | 86 |
| medium range (1 < |i-j| ≤ 4) | 42 | 36 |
| long-range (|i-j| > 4) | 213 | 169 |
| Hydrogen bond restraints (upper/lower) | 30/30 | 41/41 |
| S-S bond restraints (upper/lower) | 9/9 | 9/9 |
| J-couplings | 31 | 38 |
| JHNHα | 31 | 38 |
| Angles | 22 | 19 |
| χ1 | 19 | 19 |
| χ2 | 3 | 0 |
| Total restraints/per residue | 598/15 | 715/17 |
| Statistics of the obtained set of structures | ||
| CYANA target function | 1.26 ± 0.13 | 2.12 ± 0.19 |
| Restraints violations | ||
| distance (>0.2Å) | 2 | 5 |
| angle (>5°) | 1 | 3 |
| RMSD (Å) | ||
| for all residues | ||
| backbone | 0.98 ± 0.2 | 1.14 ± 0.32 |
| all heavy atoms | 1.98 ± 0.3 | 2.21 ± 0.42 |
| Ramachandran analysis | ||
| % residues in most favored regions | 66.7 | 60 |
| % residues in additional allowed regions | 33.3 | 40 |
| % residues in generously allowed regions | 0 | 0 |
| % residues in disallowed regions | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasheverov, I.E.; Logashina, Y.A.; Kornilov, F.D.; Lushpa, V.A.; Maleeva, E.E.; Korolkova, Y.V.; Yu, J.; Zhu, X.; Zhangsun, D.; Luo, S.; et al. Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors. Toxins 2023, 15, 28. https://doi.org/10.3390/toxins15010028
Kasheverov IE, Logashina YA, Kornilov FD, Lushpa VA, Maleeva EE, Korolkova YV, Yu J, Zhu X, Zhangsun D, Luo S, et al. Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors. Toxins. 2023; 15(1):28. https://doi.org/10.3390/toxins15010028
Chicago/Turabian StyleKasheverov, Igor E., Yulia A. Logashina, Fedor D. Kornilov, Vladislav A. Lushpa, Ekaterina E. Maleeva, Yuliya V. Korolkova, Jinpeng Yu, Xiaopeng Zhu, Dongting Zhangsun, Sulan Luo, and et al. 2023. "Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors" Toxins 15, no. 1: 28. https://doi.org/10.3390/toxins15010028
APA StyleKasheverov, I. E., Logashina, Y. A., Kornilov, F. D., Lushpa, V. A., Maleeva, E. E., Korolkova, Y. V., Yu, J., Zhu, X., Zhangsun, D., Luo, S., Stensvåg, K., Kudryavtsev, D. S., Mineev, K. S., & Andreev, Y. A. (2023). Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors. Toxins, 15(1), 28. https://doi.org/10.3390/toxins15010028