Diagnosis and Treatment for Shiga Toxin-Producing Escherichia coli Associated Hemolytic Uremic Syndrome


Abstract
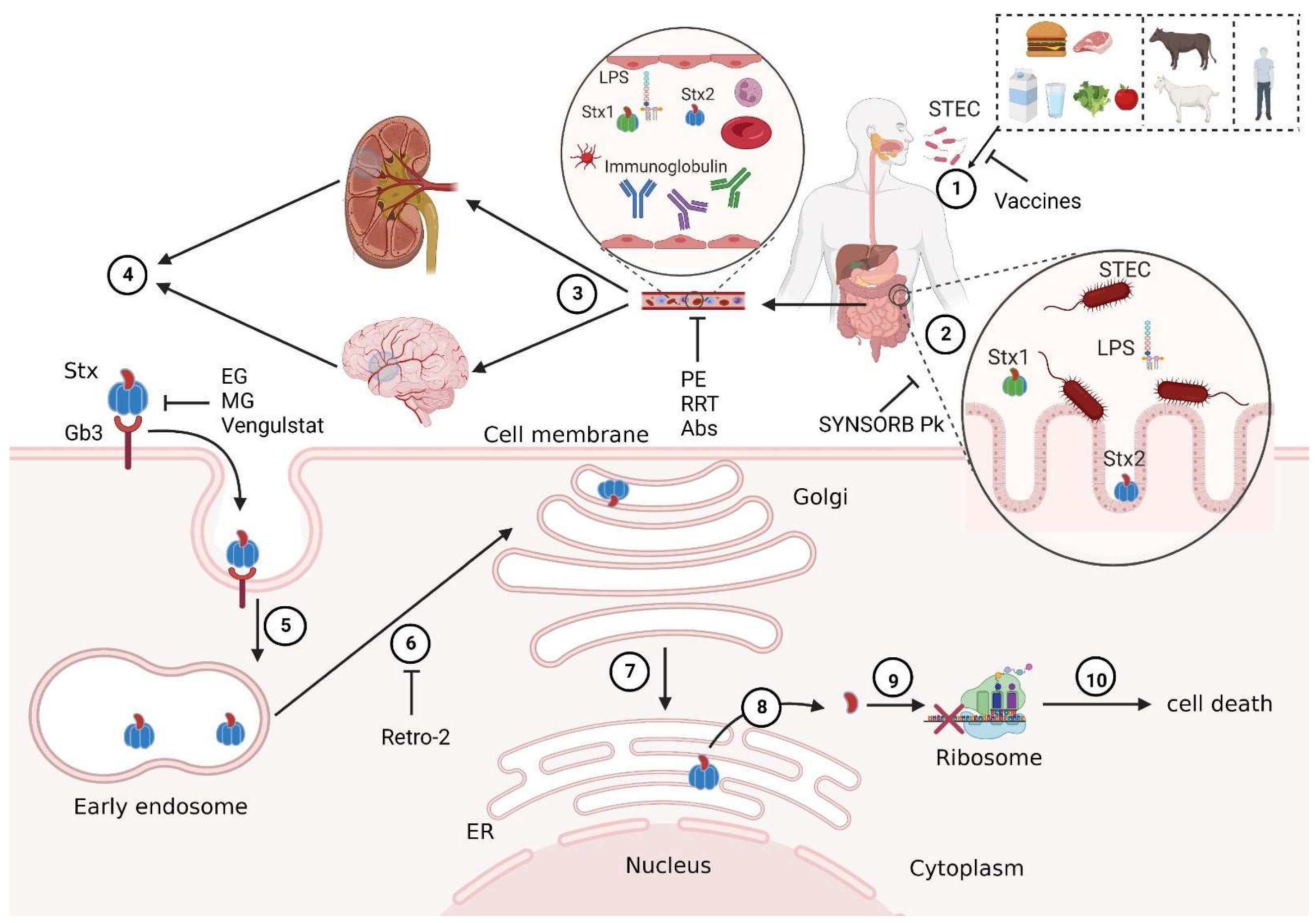
1. Introduction
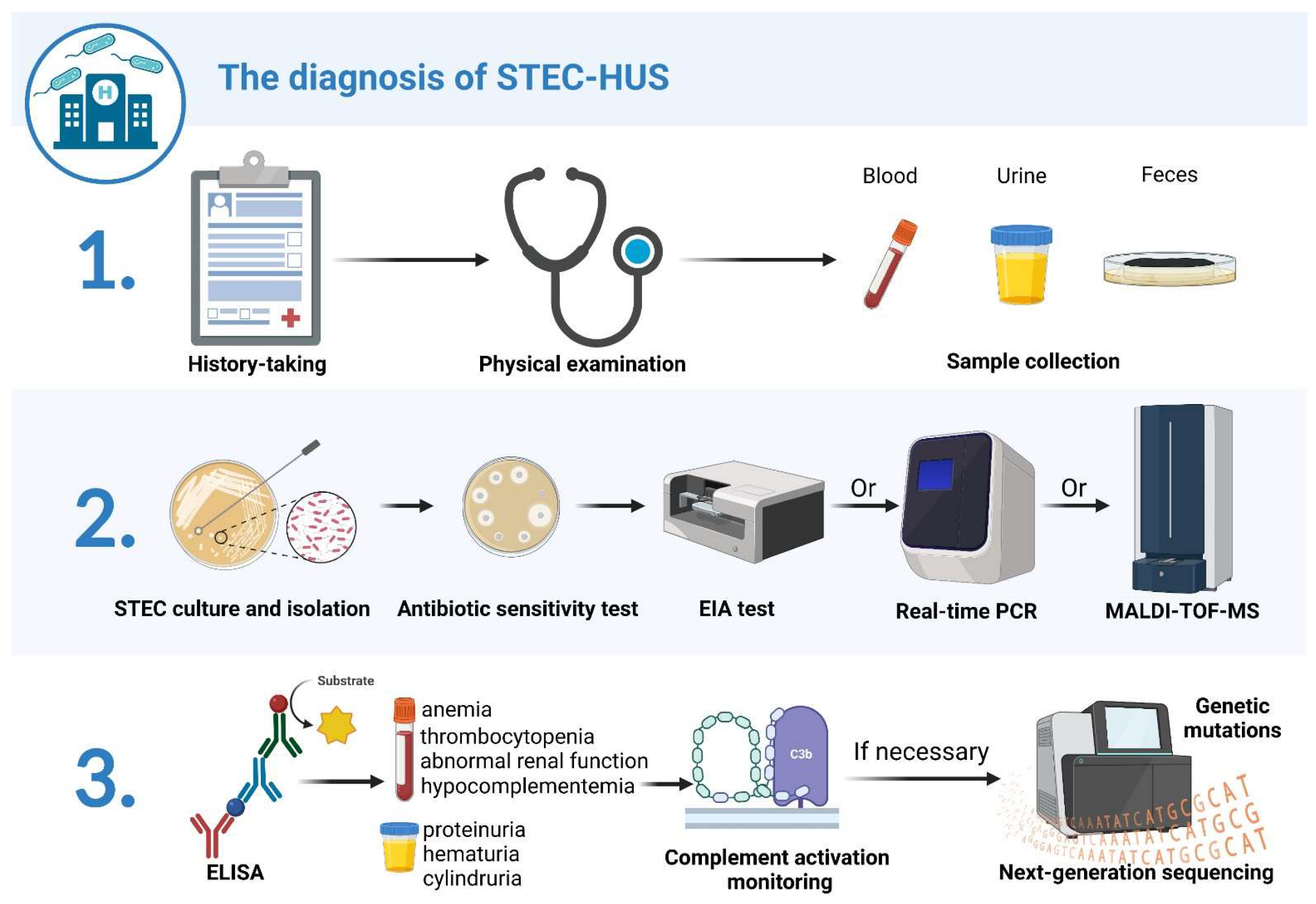
2. STEC-HUS Diagnosis
2.1. History Taking
2.2. Clinical Manifestations
2.3. Fecal Diagnostics
2.4. Serological Detection for STEC
2.5. Progress and Limitations of STEC Detection Technology
2.6. Complement Activation Monitoring
3. STEC-HUS Prevention and Precaution
4. STEC-HUS Therapeutic Strategies
4.1. Volume Expansion
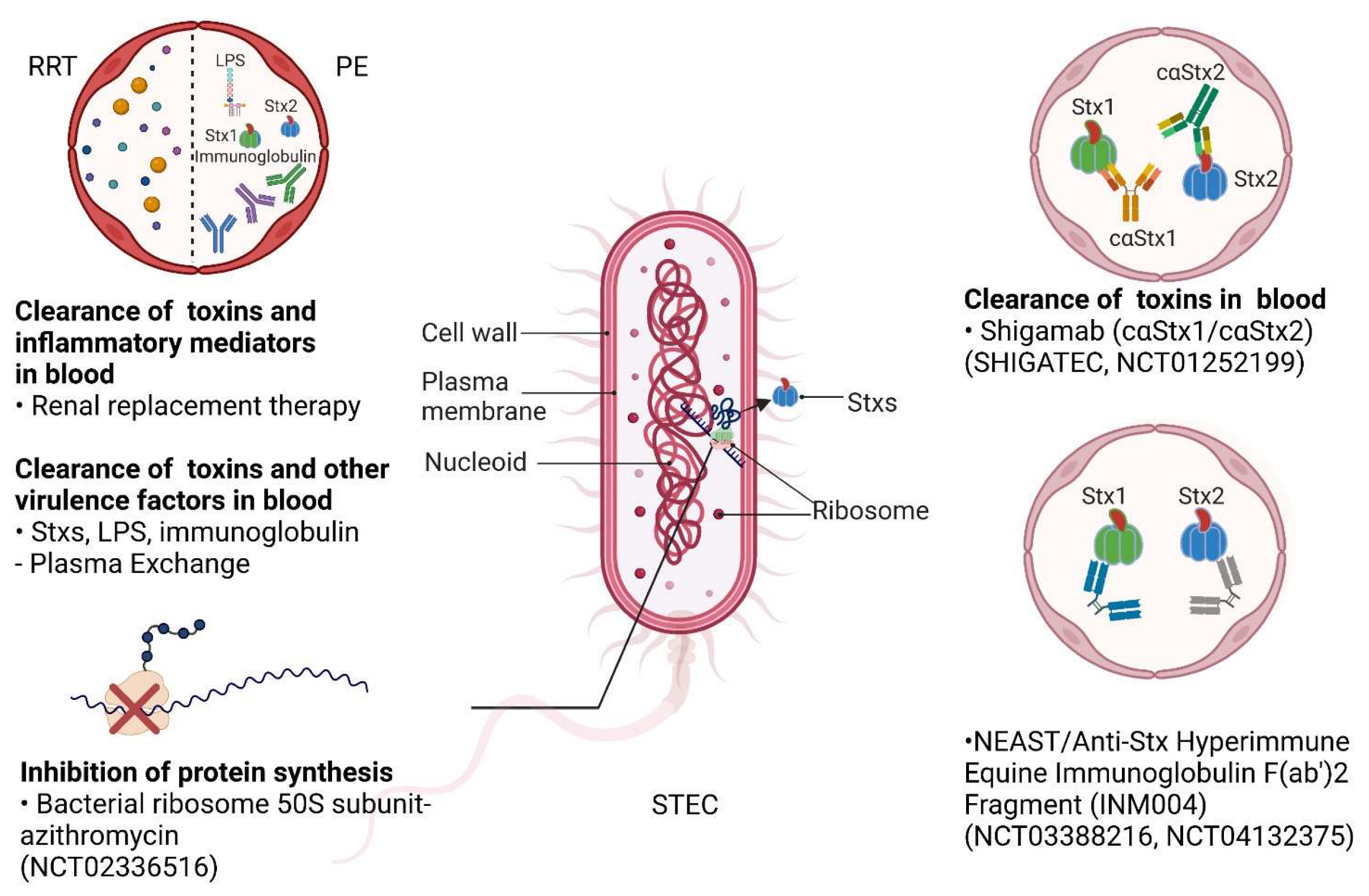
4.2. Renal Replacement Therapy
4.3. Antibiotics
4.4. Plasma Exchange
4.5. Eculizumab
4.6. Antibodies
4.7. Gb3Cer Inhibitors
4.8. SYNSORB Pk
4.9. Retro-2
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noris, M.; Remuzzi, G. Hemolytic uremic syndrome. J. Am. Soc. Nephrol. 2005, 16, 1035–1050. [Google Scholar] [CrossRef]
- Noris, M.; Remuzzi, G. Atypical hemolytic-uremic syndrome. N. Engl. J. Med. 2009, 361, 1676–1687. [Google Scholar] [CrossRef]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga Toxin-Associated Hemolytic Uremic Syndrome: A Narrative Review. Toxins 2020, 12, 67. [Google Scholar] [CrossRef]
- Ashida, A.; Matsumura, H.; Sawai, T.; Fujimaru, R.; Fujii, Y.; Shirasu, A.; Nakakura, H.; Iijima, K. Clinical features in a series of 258 Japanese pediatric patients with thrombotic microangiopathy. Clin. Exp. Nephrol. 2018, 22, 924–930. [Google Scholar] [CrossRef]
- Hamano, S.; Nakanishi, Y.; Nara, T.; Seki, T.; Ohtani, T.; Oishi, T.; Joh, K.; Oikawa, T.; Muramatsu, Y.; Ogawa, Y.; et al. Neurological manifestations of hemorrhagic colitis in the outbreak of Escherichia coli O157:H7 infection in Japan. Acta Paediatr. 1993, 82, 454–458. [Google Scholar] [CrossRef]
- Krogvold, L.; Henrichsen, T.; Bjerre, A.; Brackman, D.; Dollner, H.; Gudmundsdottir, H.; Syversen, G.; Naess, P.A.; Bangstad, H.J. Clinical aspects of a nationwide epidemic of severe haemolytic uremic syndrome (HUS) in children. Scand. J. Trauma Resusc. Emerg. Med. 2011, 19, 44. [Google Scholar] [CrossRef]
- Verweyen, H.M.; Karch, H.; Allerberger, F.; Zimmerhackl, L.B. Enterohemorrhagic Escherichia coli (EHEC) in pediatric hemolytic-uremic syndrome: A prospective study in Germany and Austria. Infection 1999, 27, 341–347. [Google Scholar] [CrossRef]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Bale, J.F., Jr.; Brasher, C.; Siegler, R.L. CNS manifestations of the hemolytic-uremic syndrome. Relationship to metabolic alterations and prognosis. Am. J. Dis. Child. 1980, 134, 869–872. [Google Scholar] [CrossRef]
- Cimolai, N.; Morrison, B.J.; Carter, J.E. Risk factors for the central nervous system manifestations of gastroenteritis-associated hemolytic-uremic syndrome. Pediatrics 1992, 90, 616–621. [Google Scholar] [CrossRef]
- Sheth, K.J.; Swick, H.M.; Haworth, N. Neurological involvement in hemolytic-uremic syndrome. Ann. Neurol. 1986, 19, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Tesh, V.L. Roles of Shiga Toxins in Immunopathology. Toxins 2019, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tian, S.; Thaker, H.; Dong, M. Shiga Toxins: An Update on Host Factors and Biomedical Applications. Toxins 2021, 13, 222. [Google Scholar] [CrossRef] [PubMed]
- te Loo, D.M.; Monnens, L.A.; van Der Velden, T.J.; Vermeer, M.A.; Preyers, F.; Demacker, P.N.; van Den Heuvel, L.P.; van Hinsbergh, V.W. Binding and transfer of verocytotoxin by polymorphonuclear leukocytes in hemolytic uremic syndrome. Blood 2000, 95, 3396–3402. [Google Scholar] [CrossRef]
- Ramegowda, B.; Samuel, J.E.; Tesh, V.L. Interaction of Shiga toxins with human brain microvascular endothelial cells: Cytokines as sensitizing agents. J. Infect. Dis. 1999, 180, 1205–1213. [Google Scholar] [CrossRef]
- Harrison, L.M.; van Haaften, W.C.; Tesh, V.L. Regulation of proinflammatory cytokine expression by Shiga toxin 1 and/or lipopolysaccharides in the human monocytic cell line THP-1. Infect. Immun. 2004, 72, 2618–2627. [Google Scholar] [CrossRef]
- Arvidsson, I.; Stahl, A.L.; Hedstrom, M.M.; Kristoffersson, A.C.; Rylander, C.; Westman, J.S.; Storry, J.R.; Olsson, M.L.; Karpman, D. Shiga toxin-induced complement-mediated hemolysis and release of complement-coated red blood cell-derived microvesicles in hemolytic uremic syndrome. J. Immunol. 2015, 194, 2309–2318. [Google Scholar] [CrossRef]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin triggers endothelial and podocyte injury: The role of complement activation. Pediatr. Nephrol. 2019, 34, 379–388. [Google Scholar] [CrossRef]
- Dhaliwal, G.; Cornett, P.A.; Tierney, L.M., Jr. Hemolytic anemia. Am. Fam. Physician 2004, 69, 2599–2606. [Google Scholar]
- Geerdink, L.M.; Westra, D.; van Wijk, J.A.; Dorresteijn, E.M.; Lilien, M.R.; Davin, J.C.; Komhoff, M.; Van Hoeck, K.; van der Vlugt, A.; van den Heuvel, L.P.; et al. Atypical hemolytic uremic syndrome in children: Complement mutations and clinical characteristics. Pediatr. Nephrol. 2012, 27, 1283–1291. [Google Scholar] [CrossRef]
- Jenkins, C.; Byrne, L.; Vishram, B.; Sawyer, C.; Balasegaram, S.; Ahyow, L.; Johnson, S. Shiga toxin-producing Escherichia coli haemolytic uraemic syndrome (STEC-HUS): Diagnosis, surveillance and public-health management in England. J. Med. Microbiol. 2020, 69, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- Awofisayo-Okuyelu, A.; Brainard, J.; Hall, I.; McCarthy, N. Incubation Period of Shiga Toxin-Producing Escherichia coli. Epidemiol. Rev. 2019, 41, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gulhan, B.; Ozaltin, F. Hemolytic Uremic Syndrome in Children. Turk. Arch. Pediatr. 2021, 56, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, K.; Barwick, K.; Fishaut, M.; Kashgarian, M.; Siegel, N.J. The importance of nonrenal involvement in hemolytic-uremic syndrome. Pediatrics 1980, 65, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Tapper, D.; Tarr, P.; Avner, E.; Brandt, J.; Waldhausen, J. Lessons learned in the management of hemolytic uremic syndrome in children. J. Pediatr. Surg. 1995, 30, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Grodinsky, S.; Telmesani, A.; Robson, W.L.; Fick, G.; Scott, R.B. Gastrointestinal manifestations of hemolytic uremic syndrome: Recognition of pancreatitis. J. Pediatr. Gastroenterol. Nutr. 1990, 11, 518–524. [Google Scholar] [CrossRef]
- Suri, R.S.; Clark, W.F.; Barrowman, N.; Mahon, J.L.; Thiessen-Philbrook, H.R.; Rosas-Arellano, M.P.; Zarnke, K.; Garland, J.S.; Garg, A.X. Diabetes during diarrhea-associated hemolytic uremic syndrome: A systematic review and meta-analysis. Diabetes Care 2005, 28, 2556–2562. [Google Scholar] [CrossRef]
- Loos, S.; Ahlenstiel, T.; Kranz, B.; Staude, H.; Pape, L.; Hartel, C.; Vester, U.; Buchtala, L.; Benz, K.; Hoppe, B.; et al. An outbreak of Shiga toxin-producing Escherichia coli O104:H4 hemolytic uremic syndrome in Germany: Presentation and short-term outcome in children. Clin. Infect. Dis. 2012, 55, 753–759. [Google Scholar] [CrossRef]
- Pena, D.R.; Vaccarello, M.; Neiberger, R.E. Severe hemolytic uremic syndrome associated with rhabdomyolysis and insulin-dependent diabetes mellitus. Child Nephrol. Urol. 1991, 11, 223–227. [Google Scholar]
- Khalid, M.; Andreoli, S. Extrarenal manifestations of the hemolytic uremic syndrome associated with Shiga toxin-producing Escherichia coli (STEC HUS). Pediatr. Nephrol. 2019, 34, 2495–2507. [Google Scholar] [CrossRef]
- Espie, E.; Grimont, F.; Mariani-Kurkdjian, P.; Bouvet, P.; Haeghebaert, S.; Filliol, I.; Loirat, C.; Decludt, B.; Minh, N.N.; Vaillant, V.; et al. Surveillance of hemolytic uremic syndrome in children less than 15 years of age, a system to monitor O157 and non-O157 Shiga toxin-producing Escherichia coli infections in France, 1996–2006. Pediatr. Infect. Dis. J. 2008, 27, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Klein, E.J.; Galanakis, E.; Thomas, A.A.; Stapp, J.R.; Rich, S.; Buccat, A.M.; Tarr, P.I. Real-Time PCR Assay for Detection and Differentiation of Shiga Toxin-Producing Escherichia coli from Clinical Samples. J. Clin. Microbiol. 2015, 53, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Gould, L.H.; Bopp, C.; Strockbine, N.; Atkinson, R.; Baselski, V.; Body, B.; Carey, R.; Crandall, C.; Hurd, S.; Kaplan, R.; et al. Recommendations for diagnosis of shiga toxin-producing Escherichia coli infections by clinical laboratories. MMWR Recomm. Rep. 2009, 58, RR-12. [Google Scholar] [CrossRef]
- Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000–2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Ardissino, G.; Vignati, C.; Masia, C.; Capone, V.; Colombo, R.; Tel, F.; Daprai, L.; Testa, S.; Dodaro, A.; Paglialonga, F.; et al. Bloody Diarrhea and Shiga Toxin-Producing Escherichia coli Hemolytic Uremic Syndrome in Children: Data from the ItalKid-HUS Network. J. Pediatr. 2021, 237, 34–40.e31. [Google Scholar] [CrossRef]
- Werber, D.; Fruth, A.; Buchholz, U.; Prager, R.; Kramer, M.H.; Ammon, A.; Tschape, H. Strong association between shiga toxin-producing Escherichia coli O157 and virulence genes stx2 and eae as possible explanation for predominance of serogroup O157 in patients with haemolytic uraemic syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 726–730. [Google Scholar] [CrossRef]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Ruter, C.; Kunsmann, L.; Greune, L.; Bauwens, A.; Zhang, W.; Kuczius, T.; Kim, K.S.; Mellmann, A.; Schmidt, M.A.; et al. Enterohemorrhagic Escherichia coli hemolysin employs outer membrane vesicles to target mitochondria and cause endothelial and epithelial apoptosis. PLoS Pathog. 2013, 9, e1003797. [Google Scholar] [CrossRef]
- van den Beld, M.J.; Reubsaet, F.A. Differentiation between Shigella, enteroinvasive Escherichia coli (EIEC) and noninvasive Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 899–904. [Google Scholar] [CrossRef]
- Wijnsma, K.L.; van Bommel, S.A.; van der Velden, T.; Volokhina, E.; Schreuder, M.F.; van den Heuvel, L.P.; van de Kar, N.C. Fecal diagnostics in combination with serology: Best test to establish STEC-HUS. Pediatr. Nephrol. 2016, 31, 2163–2170. [Google Scholar] [CrossRef]
- Jenkins, C.; Chart, H.; Smith, H.R.; Hartland, E.L.; Batchelor, M.; Delahay, R.M.; Dougan, G.; Frankel, G. Antibody response of patients infected with verocytotoxin-producing Escherichia coli to protein antigens encoded on the LEE locus. J. Med. Microbiol. 2000, 49, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Frey, E.; Mackenzie, A.M.; Finlay, B.B. Human response to Escherichia coli O157:H7 infection: Antibodies to secreted virulence factors. Infect. Immun. 2000, 68, 5090–5095. [Google Scholar] [CrossRef] [PubMed]
- Rastawicki, W.; Smietanska, K.; Rokosz-Chudziak, N.; Wolkowicz, T. Antibody response to lipopolysaccharides and recombinant proteins of Shiga toxin (STX)-producing Escherichia coli (STEC) in children with haemolytic uraemic syndrome in Poland. Lett. Appl. Microbiol. 2020, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Wijnsma, K.L.; Veissi, S.T.; van Bommel, S.A.M.; Heuver, R.; Volokhina, E.B.; Comerci, D.J.; Ugalde, J.E.; van de Kar, N.; van den Heuvel, L. Glyco-iELISA: A highly sensitive and unambiguous serological method to diagnose STEC-HUS caused by serotype O157. Pediatr. Nephrol. 2019, 34, 631–639. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Kock, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef]
- Travert, B.; Dossier, A.; Jamme, M.; Cointe, A.; Delmas, Y.; Malot, S.; Wynckel, A.; Seguin, A.; Presne, C.; Hie, M.; et al. Shiga Toxin-Associated Hemolytic Uremic Syndrome in Adults, France, 2009–2017. Emerg. Infect. Dis. 2021, 27, 1876–1885. [Google Scholar] [CrossRef]
- Fagerquist, C.K.; Sultan, O. Top-down proteomic identification of furin-cleaved alpha-subunit of Shiga toxin 2 from Escherichia coli O157:H7 using MALDI-TOF-TOF-MS/MS. J. Biomed. Biotechnol. 2010, 2010, 123460. [Google Scholar] [CrossRef]
- McLean, K.; Palarea-Albaladejo, J.; Currie, C.G.; Imrie, L.H.J.; Manson, E.D.T.; Fraser-Pitt, D.; Wright, F.; Alexander, C.J.; Pollock, K.G.J.; Allison, L.; et al. Rapid and robust analytical protocol for E. coli STEC bacteria subspecies differentiation using whole cell MALDI mass spectrometry. Talanta 2018, 182, 164–170. [Google Scholar] [CrossRef]
- Kubo, Y.; Ueda, O.; Nagamitsu, S.; Yamanishi, H.; Nakamura, A.; Komatsu, M. Novel strategy of rapid typing of Shiga toxin-producing Escherichia coli using MALDI Biotyper and ClinProTools analysis. J. Infect. Chemother. 2021, 27, 1137–1142. [Google Scholar] [CrossRef]
- Elbehiry, A.; Aldubaib, M.; Abalkhail, A.; Marzouk, E.; Albeloushi, A.; Moussa, I.; Ibrahem, M.; Albazie, H.; Alqarni, A.; Anagreyyah, S.; et al. How MALDI-TOF Mass Spectrometry Technology Contributes to Microbial Infection Control in Healthcare Settings. Vaccines 2022, 10, 1881. [Google Scholar] [CrossRef]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.K.; McKee, M.L.; O’Brien, A.D. Two copies of Shiga-like toxin II-related genes common in enterohemorrhagic Escherichia coli strains are responsible for the antigenic heterogeneity of the O157:H-strain E32511. Infect. Immun. 1991, 59, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.J.; Erickson-Beltran, M.L.; Skinner, C.B.; Dynin, I.; Hui, C.; Patfield, S.A.; Carter, J.M.; He, X. Safe and effective means of detecting and quantitating Shiga-like toxins in attomole amounts. Anal. Chem. 2014, 86, 4698–4706. [Google Scholar] [CrossRef] [PubMed]
- Fagerquist, C.K.; Zaragoza, W.J. Shiga toxin 2 subtypes of enterohemorrhagic E. coli O157:H- E32511 analyzed by RT-qPCR and top-down proteomics using MALDI-TOF-TOF-MS. J. Am. Soc. Mass Spectrom. 2015, 26, 788–799. [Google Scholar] [CrossRef]
- Skinner, C.; Patfield, S.; Stanker, L.; He, X. Development of monoclonal antibodies and immunoassays for sensitive and specific detection of Shiga toxin Stx2f. PLoS ONE 2013, 8, e76563. [Google Scholar] [CrossRef]
- Skinner, C.; Patfield, S.; Hernlem, B.J.; He, X. New Stx2e Monoclonal Antibodies for Immunological Detection and Distinction of Stx2 Subtypes. PLoS ONE 2015, 10, e0132419. [Google Scholar] [CrossRef]
- He, X.; Qi, W.; Quinones, B.; McMahon, S.; Cooley, M.; Mandrell, R.E. Sensitive detection of Shiga Toxin 2 and some of its variants in environmental samples by a novel immuno-PCR assay. Appl. Environ. Microbiol. 2011, 77, 3558–3564. [Google Scholar] [CrossRef]
- Pang, B.; Zhao, C.; Li, L.; Song, X.; Xu, K.; Wang, J.; Liu, Y.; Fu, K.; Bao, H.; Song, D.; et al. Development of a low-cost paper-based ELISA method for rapid Escherichia coli O157:H7 detection. Anal. Biochem. 2018, 542, 58–62. [Google Scholar] [CrossRef]
- Vrioni, G.; Tsiamis, C.; Oikonomidis, G.; Theodoridou, K.; Kapsimali, V.; Tsakris, A. MALDI-TOF mass spectrometry technology for detecting biomarkers of antimicrobial resistance: Current achievements and future perspectives. Ann. Transl. Med. 2018, 6, 240. [Google Scholar] [CrossRef]
- Tighe, P.J.; Ryder, R.R.; Todd, I.; Fairclough, L.C. ELISA in the multiplex era: Potentials and pitfalls. Proteomics Clin. Appl. 2015, 9, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A.L.; Sartz, L.; Karpman, D. Complement activation on platelet-leukocyte complexes and microparticles in enterohemorrhagic Escherichia coli-induced hemolytic uremic syndrome. Blood 2011, 117, 5503–5513. [Google Scholar] [CrossRef] [PubMed]
- Buelli, S.; Zoja, C.; Remuzzi, G.; Morigi, M. Complement Activation Contributes to the Pathophysiology of Shiga Toxin-Associated Hemolytic Uremic Syndrome. Microorganisms 2019, 7, 15. [Google Scholar] [CrossRef]
- Orth, D.; Khan, A.B.; Naim, A.; Grif, K.; Brockmeyer, J.; Karch, H.; Joannidis, M.; Clark, S.J.; Day, A.J.; Fidanzi, S.; et al. Shiga toxin activates complement and binds factor H: Evidence for an active role of complement in hemolytic uremic syndrome. J. Immunol. 2009, 182, 6394–6400. [Google Scholar] [CrossRef] [PubMed]
- Poolpol, K.; Orth-Holler, D.; Speth, C.; Zipfel, P.F.; Skerka, C.; de Cordoba, S.R.; Brockmeyer, J.; Bielaszewska, M.; Wurzner, R. Interaction of Shiga toxin 2 with complement regulators of the factor H protein family. Mol. Immunol. 2014, 58, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ehrlenbach, S.; Rosales, A.; Posch, W.; Wilflingseder, D.; Hermann, M.; Brockmeyer, J.; Karch, H.; Satchell, S.C.; Wurzner, R.; Orth-Holler, D. Shiga toxin 2 reduces complement inhibitor CD59 expression on human renal tubular epithelial and glomerular endothelial cells. Infect. Immun. 2013, 81, 2678–2685. [Google Scholar] [CrossRef]
- Ferraris, J.R.; Ferraris, V.; Acquier, A.B.; Sorroche, P.B.; Saez, M.S.; Ginaca, A.; Mendez, C.F. Activation of the alternative pathway of complement during the acute phase of typical haemolytic uraemic syndrome. Clin. Exp. Immunol. 2015, 181, 118–125. [Google Scholar] [CrossRef]
- Canpolat, N. Hemolytic uremic syndrome. Turk. Pediatri. Ars. 2015, 50, 73–82. [Google Scholar] [CrossRef]
- Ozaki, M.; Kang, Y.; Tan, Y.S.; Pavlov, V.I.; Liu, B.; Boyle, D.C.; Kushak, R.I.; Skjoedt, M.O.; Grabowski, E.F.; Taira, Y.; et al. Human mannose-binding lectin inhibitor prevents Shiga toxin-induced renal injury. Kidney Int. 2016, 90, 774–782. [Google Scholar] [CrossRef]
- Caillaud, C.; Zaloszyc, A.; Licht, C.; Pichault, V.; Fremeaux-Bacchi, V.; Fischbach, M. CFH gene mutation in a case of Shiga toxin-associated hemolytic uremic syndrome (STEC-HUS). Pediatr. Nephrol. 2016, 31, 157–161. [Google Scholar] [CrossRef]
- Dowen, F.; Wood, K.; Brown, A.L.; Palfrey, J.; Kavanagh, D.; Brocklebank, V. Rare genetic variants in Shiga toxin-associated haemolytic uraemic syndrome: Genetic analysis prior to transplantation is essential. Clin. Kidney J. 2017, 10, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Alberti, M.; Valoti, E.; Piras, R.; Bresin, E.; Galbusera, M.; Tripodo, C.; Thaiss, F.; Remuzzi, G.; Noris, M. Two patients with history of STEC-HUS, posttransplant recurrence and complement gene mutations. Am. J. Transplant. 2013, 13, 2201–2206. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, L.; Netti, G.S.; Torres, D.D.; Piscopo, G.; Carbone, V.; Losito, L.; Milella, L.; Lasorella, M.L.; Conti, P.; Gagliardi, D.; et al. Peripheral nervous system manifestations of Shiga toxin-producing E. coli-induced haemolytic uremic syndrome in children. Ital. J. Pediatr. 2021, 47, 181. [Google Scholar] [CrossRef] [PubMed]
- Valoti, E.; Alberti, M.; Iatropoulos, P.; Piras, R.; Mele, C.; Breno, M.; Cremaschi, A.; Bresin, E.; Donadelli, R.; Alizzi, S.; et al. Rare Functional Variants in Complement Genes and Anti-FH Autoantibodies-Associated aHUS. Front. Immunol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Noris, M.; Bresin, E.; Mele, C.; Remuzzi, G. Genetic Atypical Hemolytic-Uremic Syndrome. In GeneReviews((R)); Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Adrovic, A.; Canpolat, N.; Caliskan, S.; Sever, L.; Kiykim, E.; Agbas, A.; Baumgartner, M.R. Cobalamin C defect-hemolytic uremic syndrome caused by new mutation in MMACHC. Pediatr. Int. 2016, 58, 763–765. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, M.S.; Kim, J.H. Recent Updates on Outbreaks of Shiga Toxin-Producing Escherichia coli and Its Potential Reservoirs. Front. Cell Infect. Microbiol. 2020, 10, 273. [Google Scholar] [CrossRef]
- Rivas, M.; Chinen, I.; Guth , B.E.C. Enterohemorrhagic (Shiga Toxin-Producing) Escherichia coli. In Escherichia coli in the Americas; Torres, A., Ed.; Springer: Cham, Switzerland, 2016; pp. 97–123. [Google Scholar]
- Smith, D.R. Vaccination of Cattle against Escherichia coli O157:H7. Microbiol. Spectr. 2014, 2, 505–529. [Google Scholar] [CrossRef]
- Potter, A.A.; Klashinsky, S.; Li, Y.; Frey, E.; Townsend, H.; Rogan, D.; Erickson, G.; Hinkley, S.; Klopfenstein, T.; Moxley, R.A.; et al. Decreased shedding of Escherichia coli O157:H7 by cattle following vaccination with type III secreted proteins. Vaccine 2004, 22, 362–369. [Google Scholar] [CrossRef]
- Snedeker, K.G.; Campbell, M.; Sargeant, J.M. A systematic review of vaccinations to reduce the shedding of Escherichia coli O157 in the faeces of domestic ruminants. Zoonoses Public Health 2012, 59, 126–138. [Google Scholar] [CrossRef]
- Fingermann, M.; Avila, L.; De Marco, M.B.; Vazquez, L.; Di Biase, D.N.; Muller, A.V.; Lescano, M.; Dokmetjian, J.C.; Fernandez Castillo, S.; Perez Quinoy, J.L. OMV-based vaccine formulations against Shiga toxin producing Escherichia coli strains are both protective in mice and immunogenic in calves. Hum. Vaccin. Immunother. 2018, 14, 2208–2213. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.; Feng, S.; Zhu, L.; Guo, X.; Qi, C. Towards an attenuated enterohemorrhagic Escherichia coli O157:H7 vaccine characterized by a deleted ler gene and containing apathogenic Shiga toxins. Vaccine 2009, 27, 5929–5935. [Google Scholar] [CrossRef] [PubMed]
- Calderon Toledo, C.; Arvidsson, I.; Karpman, D. Cross-reactive protection against enterohemorrhagic Escherichia coli infection by enteropathogenic E. coli in a mouse model. Infect. Immun. 2011, 79, 2224–2233. [Google Scholar] [CrossRef]
- Oliveira, A.F.; Cardoso, S.A.; Almeida, F.B.; de Oliveira, L.L.; Pitondo-Silva, A.; Soares, S.G.; Hanna, E.S. Oral immunization with attenuated Salmonella vaccine expressing Escherichia coli O157:H7 intimin gamma triggers both systemic and mucosal humoral immunity in mice. Microbiol. Immunol. 2012, 56, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Naito, M.; Yutsudo, T.; Matsumoto, S.; Heatherly, D.P.; Yamada, T.; Kobayashi, H.; Yoshida, S.; Obrig, T. Protection by a recombinant Mycobacterium bovis Bacillus Calmette-Guerin vaccine expressing Shiga toxin 2 B subunit against Shiga toxin-producing Escherichia coli in mice. Clin. Vaccine Immunol. 2012, 19, 1932–1937. [Google Scholar] [CrossRef] [PubMed]
- Schaut, R.G.; Boggiatto, P.M.; Loving, C.L.; Sharma, V.K. Cellular and Mucosal Immune Responses Following Vaccination with Inactivated Mutant of Escherichia coli O157:H7. Sci. Rep. 2019, 9, 6401. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Feng, Y.; Luo, P.; Gu, J.; Yu, S.; Zhang, W.J.; Liu, Y.Q.; Wang, Q.X.; Zou, Q.M.; Mao, X.H. Fusion expression and immunogenicity of EHEC EspA-Stx2Al protein: Implications for the vaccine development. J. Microbiol. 2009, 47, 498–505. [Google Scholar] [CrossRef]
- Gao, X.; Cai, K.; Li, T.; Wang, Q.; Hou, X.; Tian, R.; Liu, H.; Tu, W.; Xiao, L.; Fang, L.; et al. Novel fusion protein protects against adherence and toxicity of enterohemorrhagic Escherichia coli O157:H7 in mice. Vaccine 2011, 29, 6656–6663. [Google Scholar] [CrossRef]
- Sadri Najafabadi, Z.; Nazarian, S.; Kargar, M.; Kafilzadeh, F. Designing of a chimeric protein contains StxB, intimin and EscC against toxicity and adherence of enterohemorrhagic Escherichia coli O157:H7 and evaluation of serum antibody titers against it. Mol. Immunol. 2021, 134, 218–227. [Google Scholar] [CrossRef]
- Wan, C.S.; Zhou, Y.; Yu, Y.; Peng, L.J.; Zhao, W.; Zheng, X.L. B-cell epitope KT-12 of enterohemorrhagic Escherichia coli O157:H7: A novel peptide vaccine candidate. Microbiol. Immunol. 2011, 55, 247–253. [Google Scholar] [CrossRef]
- Garcia-Angulo, V.A.; Kalita, A.; Kalita, M.; Lozano, L.; Torres, A.G. Comparative genomics and immunoinformatics approach for the identification of vaccine candidates for enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2014, 82, 2016–2026. [Google Scholar] [CrossRef]
- Riquelme-Neira, R.; Rivera, A.; Saez, D.; Fernandez, P.; Osorio, G.; del Canto, F.; Salazar, J.C.; Vidal, R.M.; Onate, A. Vaccination with DNA Encoding Truncated Enterohemorrhagic Escherichia coli (EHEC) Factor for Adherence-1 Gene (efa-1′) Confers Protective Immunity to Mice Infected with E. coli O157:H7. Front. Cell Infect. Microbiol. 2015, 5, 104. [Google Scholar] [CrossRef] [PubMed]
- Mayr, U.B.; Kudela, P.; Atrasheuskaya, A.; Bukin, E.; Ignatyev, G.; Lubitz, W. Rectal single dose immunization of mice with Escherichia coli O157:H7 bacterial ghosts induces efficient humoral and cellular immune responses and protects against the lethal heterologous challenge. Microb. Biotechnol. 2012, 5, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Tu, W.; Liu, Y.; Li, T.; Wang, H. Novel fusion antigen displayed-bacterial ghosts vaccine candidate against infection of Escherichia coli O157:H7. Sci. Rep. 2015, 5, 17479. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Li, J.; Shiloach, Y.; Robbins, J.B.; Szu, S.C. Safety and immunogenicity of Escherichia coli O157 O-specific polysaccharide conjugate vaccine in 2–5-year-old children. J. Infect. Dis. 2006, 193, 515–521. [Google Scholar] [CrossRef]
- Cataldi, A.; Yevsa, T.; Vilte, D.A.; Schulze, K.; Castro-Parodi, M.; Larzabal, M.; Ibarra, C.; Mercado, E.C.; Guzman, C.A. Efficient immune responses against Intimin and EspB of enterohaemorragic Escherichia coli after intranasal vaccination using the TLR2/6 agonist MALP-2 as adjuvant. Vaccine 2008, 26, 5662–5667. [Google Scholar] [CrossRef]
- Zhang, X.H.; He, K.W.; Zhang, S.X.; Lu, W.C.; Zhao, P.D.; Luan, X.T.; Ye, Q.; Wen, L.B.; Li, B.; Guo, R.L.; et al. Subcutaneous and intranasal immunization with Stx2B-Tir-Stx1B-Zot reduces colonization and shedding of Escherichia coli O157:H7 in mice. Vaccine 2011, 29, 3923–3929. [Google Scholar] [CrossRef]
- Iannino, F.; Uriza, P.J.; Duarte, C.M.; Pepe, M.V.; Roset, M.S.; Briones, G. Development of a Salmonella-based oral vaccine to control intestinal colonization of Shiga-toxin-producing Escherichia coli (STEC) in animals. Vaccine 2022, 40, 1065–1073. [Google Scholar] [CrossRef]
- Sanchez-Villamil, J.I.; Tapia, D.; Torres, A.G. Development of a Gold Nanoparticle Vaccine against Enterohemorrhagic Escherichia coli O157:H7. mBio 2019, 10, e01869-19. [Google Scholar] [CrossRef]
- Rojas-Lopez, M.; Monterio, R.; Pizza, M.; Desvaux, M.; Rosini, R. Intestinal Pathogenic Escherichia coli: Insights for Vaccine Development. Front. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Beneke, J.; Sartison, A.; Kielstein, J.T.; Haller, H.; Nitschke, M.; Kunzendorf, U.; Loos, S.; Kemper, M.J.; Stahl, R.A.; Menne, J.; et al. Clinical and Laboratory Consequences of Platelet Transfusion in Shiga Toxin-Mediated Hemolytic Uremic Syndrome. Transfus. Med. Rev. 2017, 31, 51–55. [Google Scholar] [CrossRef]
- Mele, C.; Remuzzi, G.; Noris, M. Hemolytic uremic syndrome. Semin. Immunopathol. 2014, 36, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Harkins, V.J.; McAllister, D.A.; Reynolds, B.C. Shiga-Toxin E. coli Hemolytic Uremic Syndrome: Review of Management and Long-term Outcome. Curr. Pediatr. Rep. 2020, 8, 16–25. [Google Scholar] [CrossRef]
- Bruyand, M.; Mariani-Kurkdjian, P.; Gouali, M.; de Valk, H.; King, L.A.; Le Hello, S.; Bonacorsi, S.; Loirat, C. Hemolytic uremic syndrome due to Shiga toxin-producing Escherichia coli infection. Médecine Mal. Infect. 2018, 48, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.; Kamariski, M.; Principi, I.; Bocanegra, V.; Valles, P.G. Severely ill pediatric patients with Shiga toxin-associated hemolytic uremic syndrome (STEC-HUS) who suffered from multiple organ involvement in the early stage. Pediatr. Nephrol. 2021, 36, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.W.; Estes, M.K.; Langridge, W.H. Oral immunization with a shiga toxin B subunit: Rotavirus NSP4(90) fusion protein protects mice against gastroenteritis. Vaccine 2005, 23, 5168–5176. [Google Scholar] [CrossRef]
- Tremblay, J.M.; Mukherjee, J.; Leysath, C.E.; Debatis, M.; Ofori, K.; Baldwin, K.; Boucher, C.; Peters, R.; Beamer, G.; Sheoran, A.; et al. A single VHH-based toxin-neutralizing agent and an effector antibody protect mice against challenge with Shiga toxins 1 and 2. Infect. Immun. 2013, 81, 4592–4603. [Google Scholar] [CrossRef]
- Muhlen, S.; Dersch, P. Treatment Strategies for Infections With Shiga Toxin-Producing Escherichia coli. Front. Cell Infect. Microbiol. 2020, 10, 169. [Google Scholar] [CrossRef]
- Silberstein, C.; Lucero, M.S.; Zotta, E.; Copeland, D.P.; Lingyun, L.; Repetto, H.A.; Ibarra, C. A glucosylceramide synthase inhibitor protects rats against the cytotoxic effects of shiga toxin 2. Pediatr. Res. 2011, 69, 390–394. [Google Scholar] [CrossRef]
- Trachtman, H.; Cnaan, A.; Christen, E.; Gibbs, K.; Zhao, S.; Acheson, D.W.; Weiss, R.; Kaskel, F.J.; Spitzer, A.; Hirschman, G.H.; et al. Effect of an oral Shiga toxin-binding agent on diarrhea-associated hemolytic uremic syndrome in children: A randomized controlled trial. JAMA 2003, 290, 1337–1344. [Google Scholar] [CrossRef]
- Armstrong, G.D.; Rowe, P.C.; Goodyer, P.; Orrbine, E.; Klassen, T.P.; Wells, G.; MacKenzie, A.; Lior, H.; Blanchard, C.; Auclair, F.; et al. A phase I study of chemically synthesized verotoxin (Shiga-like toxin) Pk-trisaccharide receptors attached to chromosorb for preventing hemolytic-uremic syndrome. J. Infect. Dis. 1995, 171, 1042–1045. [Google Scholar] [CrossRef]
- Imdad, A.; Mackoff, S.P.; Urciuoli, D.M.; Syed, T.; Tanner-Smith, E.E.; Huang, D.; Gomez-Duarte, O.G. Interventions for preventing diarrhoea-associated haemolytic uraemic syndrome. Cochrane Database Syst. Rev. 2021, 7, CD012997. [Google Scholar] [CrossRef] [PubMed]
- Bitzan, M.; Poole, R.; Mehran, M.; Sicard, E.; Brockus, C.; Thuning-Roberson, C.; Riviere, M. Safety and pharmacokinetics of chimeric anti-Shiga toxin 1 and anti-Shiga toxin 2 monoclonal antibodies in healthy volunteers. Antimicrob. Agents Chemother. 2009, 53, 3081–3087. [Google Scholar] [CrossRef] [PubMed]
- Strockbine, N.A.; Marques, L.R.; Holmes, R.K.; O’Brien, A.D. Characterization of monoclonal antibodies against Shiga-like toxin from Escherichia coli. Infect. Immun. 1985, 50, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Dowling, T.C.; Chavaillaz, P.A.; Young, D.G.; Melton-Celsa, A.; O’Brien, A.; Thuning-Roberson, C.; Edelman, R.; Tacket, C.O. Phase 1 safety and pharmacokinetic study of chimeric murine-human monoclonal antibody c alpha Stx2 administered intravenously to healthy adult volunteers. Antimicrob. Agents Chemother. 2005, 49, 1808–1812. [Google Scholar] [CrossRef] [PubMed]
- Muff-Luett, M.; Sanderson, K.R.; Engen, R.M.; Zahr, R.S.; Wenderfer, S.E.; Tran, C.L.; Sharma, S.; Cai, Y.; Ingraham, S.; Winnicki, E.; et al. Eculizumab exposure in children and young adults: Indications, practice patterns, and outcomes-a Pediatric Nephrology Research Consortium study. Pediatr. Nephrol. 2021, 36, 2349–2360. [Google Scholar] [CrossRef]
- Yanina, H.; Romina, P.; Lucas, B.; Constanza, L.; Luciana, M.; L, B.; Mariana, C.; Ortega, H.; A, G.; Santiago, S.; et al. Preclinical Studies of NEAST (Neutralizing Equine Anti-Shiga To xin): A Potential Treatment for Prevention of Stec-Hus. Int. J. Drug Dev. Res. 2019, 11. [Google Scholar] [CrossRef]
- Balestracci, A.; Capone, M.A.; Meni Battaglia, L.; Toledo, I.; Martin, S.M.; Beaudoin, L.; Balbaryski, J.; Gomez, L. Erythropoietin in children with hemolytic uremic syndrome: A pilot randomized controlled trial. Pediatr. Nephrol. 2022, 37, 2383–2392. [Google Scholar] [CrossRef]
- Balestracci, A.; Martin, S.M.; Toledo, I.; Alvarado, C.; Wainsztein, R.E. Early erythropoietin in post-diarrheal hemolytic uremic syndrome: A case-control study. Pediatr. Nephrol. 2015, 30, 339–344. [Google Scholar] [CrossRef]
- Pape, L.; Ahlenstiel, T.; Kreuzer, M.; Drube, J.; Froede, K.; Franke, D.; Ehrich, J.H.; Haubitz, M. Early erythropoietin reduced the need for red blood cell transfusion in childhood hemolytic uremic syndrome: A randomized prospective pilot trial. Pediatr. Nephrol. 2009, 24, 1061–1064. [Google Scholar] [CrossRef]
- Ake, J.A.; Jelacic, S.; Ciol, M.A.; Watkins, S.L.; Murray, K.F.; Christie, D.L.; Klein, E.J.; Tarr, P.I. Relative nephroprotection during Escherichia coli O157:H7 infections: Association with intravenous volume expansion. Pediatrics 2005, 115, e673–e680. [Google Scholar] [CrossRef]
- Balestracci, A.; Martin, S.M.; Toledo, I.; Alvarado, C.; Wainsztein, R.E. Dehydration at admission increased the need for dialysis in hemolytic uremic syndrome children. Pediatr. Nephrol. 2012, 27, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Ardissino, G.; Dacco, V.; Testa, S.; Civitillo, C.F.; Tel, F.; Possenti, I.; Belingheri, M.; Castorina, P.; Bolsa-Ghiringhelli, N.; Tedeschi, S.; et al. Hemoconcentration: A major risk factor for neurological involvement in hemolytic uremic syndrome. Pediatr. Nephrol. 2015, 30, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Grisaru, S.; Xie, J.; Samuel, S.; Hartling, L.; Tarr, P.I.; Schnadower, D.; Freedman, S.B.; Alberta Alberta Provincial Pediatric Enteric Infection Team. Associations Between Hydration Status, Intravenous Fluid Administration, and Outcomes of Patients Infected With Shiga Toxin-Producing Escherichia coli: A Systematic Review and Meta-analysis. JAMA Pediatr. 2017, 171, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.K.; Van De Kar, N.; Tarr, P.I. Shiga Toxin/Verocytotoxin-Producing Escherichia coli Infections: Practical Clinical Perspectives. Microbiol. Spectr. 2014, 2, EHEC-0025-2014. [Google Scholar] [CrossRef] [PubMed]
- Hickey, C.A.; Beattie, T.J.; Cowieson, J.; Miyashita, Y.; Strife, C.F.; Frem, J.C.; Peterson, J.M.; Butani, L.; Jones, D.P.; Havens, P.L.; et al. Early volume expansion during diarrhea and relative nephroprotection during subsequent hemolytic uremic syndrome. Arch. Pediatr. Adolesc. Med. 2011, 165, 884–889. [Google Scholar] [CrossRef]
- Ardissino, G.; Tel, F.; Possenti, I.; Testa, S.; Consonni, D.; Paglialonga, F.; Salardi, S.; Borsa-Ghiringhelli, N.; Salice, P.; Tedeschi, S.; et al. Early Volume Expansion and Outcomes of Hemolytic Uremic Syndrome. Pediatrics 2016, 137, e20152153. [Google Scholar] [CrossRef]
- Mody, R.K.; Gu, W.; Griffin, P.M.; Jones, T.F.; Rounds, J.; Shiferaw, B.; Tobin-D’Angelo, M.; Smith, G.; Spina, N.; Hurd, S.; et al. Postdiarrheal hemolytic uremic syndrome in United States children: Clinical spectrum and predictors of in-hospital death. J. Pediatr. 2015, 166, 1022–1029. [Google Scholar] [CrossRef]
- Wong, C.S.; Mooney, J.C.; Brandt, J.R.; Staples, A.O.; Jelacic, S.; Boster, D.R.; Watkins, S.L.; Tarr, P.I. Risk factors for the hemolytic uremic syndrome in children infected with Escherichia coli O157:H7: A multivariable analysis. Clin. Infect. Dis. 2012, 55, 33–41. [Google Scholar] [CrossRef]
- Trachtman, H.; Austin, C.; Lewinski, M.; Stahl, R.A. Renal and neurological involvement in typical Shiga toxin-associated HUS. Nat. Rev. Nephrol. 2012, 8, 658–669. [Google Scholar] [CrossRef]
- Coccia, P.A.; Ramirez, F.B.; Suarez, A.D.C.; Alconcher, L.F.; Balestracci, A.; Garcia Chervo, L.A.; Principi, I.; Vazquez, A.; Ratto, V.M.; Planells, M.C.; et al. Acute peritoneal dialysis, complications and outcomes in 389 children with STEC-HUS: A multicenter experience. Pediatr Nephrol 2021, 36, 1597–1606. [Google Scholar] [CrossRef]
- de Galasso, L.; Picca, S.; Guzzo, I. Dialysis modalities for the management of pediatric acute kidney injury. Pediatr. Nephrol. 2020, 35, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Chionh, C.Y.; Soni, S.S.; Finkelstein, F.O.; Ronco, C.; Cruz, D.N. Use of peritoneal dialysis in AKI: A systematic review. Clin. J. Am. Soc. Nephrol. 2013, 8, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Ponce, D.; Gobo-Oliveira, M.; Balbi, A.L. Peritoneal Dialysis Treatment Modality Option in Acute Kidney Injury. Blood Purif. 2017, 43, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Cullis, B.; Abdelraheem, M.; Abrahams, G.; Balbi, A.; Cruz, D.N.; Frishberg, Y.; Koch, V.; McCulloch, M.; Numanoglu, A.; Nourse, P.; et al. Peritoneal dialysis for acute kidney injury. Perit. Dial. Int. 2014, 34, 494–517. [Google Scholar] [CrossRef]
- Walters, S.; Porter, C.; Brophy, P.D. Dialysis and pediatric acute kidney injury: Choice of renal support modality. Pediatr. Nephrol. 2009, 24, 37–48. [Google Scholar] [CrossRef]
- Strazdins, V.; Watson, A.R.; Harvey, B.; European Pediatric Peritoneal Sialysis Working, G. Renal replacement therapy for acute renal failure in children: European guidelines. Pediatr. Nephrol. 2004, 19, 199–207. [Google Scholar] [CrossRef]
- Symons, J.M.; Chua, A.N.; Somers, M.J.; Baum, M.A.; Bunchman, T.E.; Benfield, M.R.; Brophy, P.D.; Blowey, D.; Fortenberry, J.D.; Chand, D.; et al. Demographic characteristics of pediatric continuous renal replacement therapy: A report of the prospective pediatric continuous renal replacement therapy registry. Clin. J. Am. Soc. Nephrol. 2007, 2, 732–738. [Google Scholar] [CrossRef]
- Ostermann, M.; Bellomo, R.; Burdmann, E.A.; Doi, K.; Endre, Z.H.; Goldstein, S.L.; Kane-Gill, S.L.; Liu, K.D.; Prowle, J.R.; Shaw, A.D.; et al. Controversies in acute kidney injury: Conclusions from a Kidney Disease: Improving Global Outcomes (KDIGO) Conference. Kidney Int. 2020, 98, 294–309. [Google Scholar] [CrossRef]
- Hoste, E.A.J.; Kellum, J.A.; Selby, N.M.; Zarbock, A.; Palevsky, P.M.; Bagshaw, S.M.; Goldstein, S.L.; Cerda, J.; Chawla, L.S. Global epidemiology and outcomes of acute kidney injury. Nat. Rev. Nephrol. 2018, 14, 607–625. [Google Scholar] [CrossRef]
- Agger, M.; Scheutz, F.; Villumsen, S.; Molbak, K.; Petersen, A.M. Antibiotic treatment of verocytotoxin-producing Escherichia coli (VTEC) infection: A systematic review and a proposal. J. Antimicrob. Chemother. 2015, 70, 2440–2446. [Google Scholar] [CrossRef]
- Freedman, S.B.; Xie, J.; Neufeld, M.S.; Hamilton, W.L.; Hartling, L.; Tarr, P.I.; Alberta Provincial Pediatric Enteric Infection, T.; Nettel-Aguirre, A.; Chuck, A.; Lee, B.; et al. Shiga Toxin-Producing Escherichia coli Infection, Antibiotics, and Risk of Developing Hemolytic Uremic Syndrome: A Meta-analysis. Clin. Infect. Dis. 2016, 62, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Safdar, N.; Said, A.; Gangnon, R.E.; Maki, D.G. Risk of hemolytic uremic syndrome after antibiotic treatment of Escherichia coli O157:H7 enteritis: A meta-analysis. JAMA 2002, 288, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.S.; Jelacic, S.; Habeeb, R.L.; Watkins, S.L.; Tarr, P.I. The risk of the hemolytic-uremic syndrome after antibiotic treatment of Escherichia coli O157:H7 infections. N. Engl. J. Med. 2000, 342, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Kakoullis, L.; Papachristodoulou, E.; Chra, P.; Panos, G. Shiga toxin-induced haemolytic uraemic syndrome and the role of antibiotics: A global overview. J. Infect. 2019, 79, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Ida, O.; Kimoto, K.; Takatorige, T.; Nakanishi, N.; Tatara, K. Effect of early fosfomycin treatment on prevention of hemolytic uremic syndrome accompanying Escherichia coli O157:H7 infection. Clin. Nephrol. 1999, 52, 357–362. [Google Scholar] [PubMed]
- Slutsker, L.; Ries, A.A.; Maloney, K.; Wells, J.G.; Greene, K.D.; Griffin, P.M. A nationwide case-control study of Escherichia coli O157:H7 infection in the United States. J. Infect. Dis. 1998, 177, 962–966. [Google Scholar] [CrossRef]
- Dundas, S.; Todd, W.T.; Stewart, A.I.; Murdoch, P.S.; Chaudhuri, A.K.; Hutchinson, S.J. The central Scotland Escherichia coli O157:H7 outbreak: Risk factors for the hemolytic uremic syndrome and death among hospitalized patients. Clin. Infect. Dis. 2001, 33, 923–931. [Google Scholar] [CrossRef]
- Proulx, F.; Turgeon, J.P.; Delage, G.; Lafleur, L.; Chicoine, L. Randomized, controlled trial of antibiotic therapy for Escherichia coli O157:H7 enteritis. J. Pediatr. 1992, 121, 299–303. [Google Scholar] [CrossRef]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef]
- Kimmitt, P.T.; Harwood, C.R.; Barer, M.R. Toxin gene expression by shiga toxin-producing Escherichia coli: The role of antibiotics and the bacterial SOS response. Emerg. Infect. Dis. 2000, 6, 458–465. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef] [PubMed]
- McGannon, C.M.; Fuller, C.A.; Weiss, A.A. Different classes of antibiotics differentially influence shiga toxin production. Antimicrob. Agents Chemother. 2010, 54, 3790–3798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Donohue-Rolfe, A.; Krautz-Peterson, G.; Sevo, M.; Parry, N.; Abeijon, C.; Tzipori, S. Gnotobiotic piglet infection model for evaluating the safe use of antibiotics against Escherichia coli O157:H7 infection. J. Infect. Dis. 2009, 199, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Walterspiel, J.N.; Ashkenazi, S.; Morrow, A.L.; Cleary, T.G. Effect of subinhibitory concentrations of antibiotics on extracellular Shiga-like toxin I. Infection 1992, 20, 25–29. [Google Scholar] [CrossRef]
- Grif, K.; Dierich, M.P.; Karch, H.; Allerberger, F. Strain-specific differences in the amount of Shiga toxin released from enterohemorrhagic Escherichia coli O157 following exposure to subinhibitory concentrations of antimicrobial agents. Eur. J. Clin. Microbiol. Infect. Dis. 1998, 17, 761–766. [Google Scholar] [CrossRef]
- Corogeanu, D.; Willmes, R.; Wolke, M.; Plum, G.; Utermohlen, O.; Kronke, M. Therapeutic concentrations of antibiotics inhibit Shiga toxin release from enterohemorrhagic E. coli O104:H4 from the 2011 German outbreak. BMC Microbiol. 2012, 12, 160. [Google Scholar] [CrossRef]
- Amran, M.Y.; Fujii, J.; Suzuki, S.O.; Kolling, G.L.; Villanueva, S.Y.; Kainuma, M.; Kobayashi, H.; Kameyama, H.; Yoshida, S. Investigation of encephalopathy caused by Shiga toxin 2c-producing Escherichia coli infection in mice. PLoS ONE 2013, 8, e58959. [Google Scholar] [CrossRef]
- Yoshimura, K.; Fujii, J.; Taniguchi, H.; Yoshida, S. Chemotherapy for enterohemorrhagic Escherichia coli O157:H infection in a mouse model. FEMS Immunol. Med. Microbiol. 1999, 26, 101–108. [Google Scholar] [CrossRef]
- Sawamura, S.; Tanaka, K.; Koga, Y. Therapeutic effects of antibiotics against enterohemorrhagic Escherichia coli (EHEC) O157:H7 (O157) infection: In vivo analysis using germfree mice. Kansenshogaku Zasshi 1999, 73, 1054–1063. [Google Scholar] [CrossRef]
- Kurioka, T.; Yunou, Y.; Harada, H.; Kita, E. Efficacy of antibiotic therapy for infection with Shiga-like toxin-producing Escherichia coli O157:H7 in mice with protein-calorie malnutrition. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 561–571. [Google Scholar] [CrossRef]
- Launders, N.; Byrne, L.; Jenkins, C.; Harker, K.; Charlett, A.; Adak, G.K. Disease severity of Shiga toxin-producing E. coli O157 and factors influencing the development of typical haemolytic uraemic syndrome: A retrospective cohort study, 2009–2012. BMJ Open 2016, 6, e009933. [Google Scholar] [CrossRef] [PubMed]
- Geerdes-Fenge, H.F.; Lobermann, M.; Nurnberg, M.; Fritzsche, C.; Koball, S.; Henschel, J.; Hohn, R.; Schober, H.C.; Mitzner, S.; Podbielski, A.; et al. Ciprofloxacin reduces the risk of hemolytic uremic syndrome in patients with Escherichia coli O104:H4-associated diarrhea. Infection 2013, 41, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Nakamura-Uchiyama, F. Does levofloxacin induce hemolytic uremic syndrome in patients infected with verotoxin-producing Escherichia coli O157 infections? Jpn. J. Infect. Dis. 2012, 65, 442–443. [Google Scholar] [CrossRef] [PubMed]
- Menne, J.; Nitschke, M.; Stingele, R.; Abu-Tair, M.; Beneke, J.; Bramstedt, J.; Bremer, J.P.; Brunkhorst, R.; Busch, V.; Dengler, R.; et al. Validation of treatment strategies for enterohaemorrhagic Escherichia coli O104:H4 induced haemolytic uraemic syndrome: Case-control study. BMJ 2012, 345, e4565. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Freedman, S.B. Why antibiotics should not be used to treat Shiga toxin-producing Escherichia coli infections. Curr. Opin. Gastroenterol. 2022, 38, 30–38. [Google Scholar] [CrossRef]
- Ramstad, S.N.; Taxt, A.M.; Naseer, U.; Wasteson, Y.; Bjornholt, J.V.; Brandal, L.T. Effects of antimicrobials on Shiga toxin production in high-virulent Shiga toxin-producing Escherichia coli. Microb. Pathog. 2021, 152, 104636. [Google Scholar] [CrossRef]
- Keenswijk, W.; Raes, A.; De Clerck, M.; Vande Walle, J. Is Plasma Exchange Efficacious in Shiga Toxin-Associated Hemolytic Uremic Syndrome? A Narrative Review of Current Evidence. Ther. Apher. Dial. 2019, 23, 118–125. [Google Scholar] [CrossRef]
- Gerber, A.; Karch, H.; Allerberger, F.; Verweyen, H.M.; Zimmerhackl, L.B. Clinical course and the role of shiga toxin-producing Escherichia coli infection in the hemolytic-uremic syndrome in pediatric patients, 1997-2000, in Germany and Austria: A prospective study. J. Infect. Dis. 2002, 186, 493–500. [Google Scholar] [CrossRef]
- Martin, D.L.; MacDonald, K.L.; White, K.E.; Soler, J.T.; Osterholm, M.T. The epidemiology and clinical aspects of the hemolytic uremic syndrome in Minnesota. N. Engl. J. Med. 1990, 323, 1161–1167. [Google Scholar] [CrossRef]
- Banatvala, N.; Griffin, P.M.; Greene, K.D.; Barrett, T.J.; Bibb, W.F.; Green, J.H.; Wells, J.G.; Hemolytic Uremic Syndrome Study, C. The United States National Prospective Hemolytic Uremic Syndrome Study: Microbiologic, serologic, clinical, and epidemiologic findings. J. Infect. Dis. 2001, 183, 1063–1070. [Google Scholar] [CrossRef]
- Colic, E.; Dieperink, H.; Titlestad, K.; Tepel, M. Management of an acute outbreak of diarrhoea-associated haemolytic uraemic syndrome with early plasma exchange in adults from southern Denmark: An observational study. Lancet 2011, 378, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Dundas, S.; Murphy, J.; Soutar, R.L.; Jones, G.A.; Hutchinson, S.J.; Todd, W.T. Effectiveness of therapeutic plasma exchange in the 1996 Lanarkshire Escherichia coli O157:H7 outbreak. Lancet 1999, 354, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, T.; Tsuchida, K.; Yoshimura, R.; Sugimura, K.; Takemoto, Y. Plasma exchange therapy for the treatment of Escherichia coli O-157 associated hemolytic uremic syndrome. Int. J. Mol. Med. 2002, 10, 585–588. [Google Scholar] [PubMed]
- Igarashi, T.; Ito, S.; Sako, M.; Saitoh, A.; Hataya, H.; Mizuguchi, M.; Morishima, T.; Ohnishi, K.; Kawamura, N.; Kitayama, H.; et al. Guidelines for the management and investigation of hemolytic uremic syndrome. Clin. Exp. Nephrol. 2014, 18, 525–557. [Google Scholar] [CrossRef]
- Legendre, C.M.; Licht, C.; Muus, P.; Greenbaum, L.A.; Babu, S.; Bedrosian, C.; Bingham, C.; Cohen, D.J.; Delmas, Y.; Douglas, K.; et al. Terminal complement inhibitor eculizumab in atypical hemolytic-uremic syndrome. N. Engl. J. Med. 2013, 368, 2169–2181. [Google Scholar] [CrossRef]
- Fakhouri, F.; Hourmant, M.; Campistol, J.M.; Cataland, S.R.; Espinosa, M.; Gaber, A.O.; Menne, J.; Minetti, E.E.; Provot, F.; Rondeau, E.; et al. Terminal Complement Inhibitor Eculizumab in Adult Patients With Atypical Hemolytic Uremic Syndrome: A Single-Arm, Open-Label Trial. Am. J. Kidney Dis. 2016, 68, 84–93. [Google Scholar] [CrossRef]
- Umscheid, J.H.; Nevil, C.; Vasudeva, R.; Ali, M.F.; Agasthya, N. Treatment of Shiga-Toxin Hus with Severe Neurologic Features with Eculizumab. Case Rep. Pediatr. 2021, 2021, 8053246. [Google Scholar] [CrossRef]
- Mahat, U.; Matar, R.B.; Rotz, S.J. Use of complement monoclonal antibody eculizumab in Shiga toxin producing Escherichia coli associated hemolytic uremic syndrome: A review of current evidence. Pediatr. Blood Cancer 2019, 66, e27913. [Google Scholar] [CrossRef]
- Lapeyraque, A.L.; Malina, M.; Fremeaux-Bacchi, V.; Boppel, T.; Kirschfink, M.; Oualha, M.; Proulx, F.; Clermont, M.J.; Le Deist, F.; Niaudet, P.; et al. Eculizumab in severe Shiga-toxin-associated HUS. N. Engl. J. Med. 2011, 364, 2561–2563. [Google Scholar] [CrossRef]
- Monet-Didailler, C.; Chevallier, A.; Godron-Dubrasquet, A.; Allard, L.; Delmas, Y.; Contin-Bordes, C.; Brissaud, O.; Llanas, B.; Harambat, J. Outcome of children with Shiga toxin-associated haemolytic uraemic syndrome treated with eculizumab: A matched cohort study. Nephrol. Dial. Transplant. 2020, 35, 2147–2153. [Google Scholar] [CrossRef]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef] [PubMed]
- Henrique, I.M.; Sacerdoti, F.; Ferreira, R.L.; Henrique, C.; Amaral, M.M.; Piazza, R.M.F.; Luz, D. Therapeutic Antibodies Against Shiga Toxins: Trends and Perspectives. Front. Cell Infect. Microbiol. 2022, 12, 825856. [Google Scholar] [CrossRef] [PubMed]
- Dixit, R.; Herz, J.; Dalton, R.; Booy, R. Benefits of using heterologous polyclonal antibodies and potential applications to new and undertreated infectious pathogens. Vaccine 2016, 34, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Mejias, M.P.; Ghersi, G.; Craig, P.O.; Panek, C.A.; Bentancor, L.V.; Baschkier, A.; Goldbaum, F.A.; Zylberman, V.; Palermo, M.S. Immunization with a chimera consisting of the B subunit of Shiga toxin type 2 and brucella lumazine synthase confers total protection against Shiga toxins in mice. J. Immunol. 2013, 191, 2403–2411. [Google Scholar] [CrossRef]
- Hiriart, Y.; Pardo, R.; Bukata, L.; Lauche, C.; Munoz, L.; Colonna, M.; Goldbaum, F.; Sanguineti, S.; Zylberman, V. [Development of a product anti-Shiga toxin for prevention of the hemolytic uremic syndrome]. Medicina 2018, 78, 107–112. [Google Scholar]
- Lopez, E.L.; Contrini, M.M.; Glatstein, E.; Gonzalez Ayala, S.; Santoro, R.; Allende, D.; Ezcurra, G.; Teplitz, E.; Koyama, T.; Matsumoto, Y.; et al. Safety and pharmacokinetics of urtoxazumab, a humanized monoclonal antibody, against Shiga-like toxin 2 in healthy adults and in pediatric patients infected with Shiga-like toxin-producing Escherichia coli. Antimicrob. Agents Chemother. 2010, 54, 239–243. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R.; Carvalho, H.M.; Thuning-Roberson, C.; O’Brien, A.D. Protective efficacy and pharmacokinetics of human/mouse chimeric anti-Stx1 and anti-Stx2 antibodies in mice. Clin. Vaccine Immunol. 2015, 22, 448–455. [Google Scholar] [CrossRef]
- Moxley, R.A.; Francis, D.H.; Tamura, M.; Marx, D.B.; Santiago-Mateo, K.; Zhao, M. Efficacy of Urtoxazumab (TMA-15 Humanized Monoclonal Antibody Specific for Shiga Toxin 2) Against Post-Diarrheal Neurological Sequelae Caused by Escherichia coli O157:H7 Infection in the Neonatal Gnotobiotic Piglet Model. Toxins 2017, 9, 49. [Google Scholar] [CrossRef]
- Yamagami, S.; Motoki, M.; Kimura, T.; Izumi, H.; Takeda, T.; Katsuura, Y.; Matsumoto, Y. Efficacy of postinfection treatment with anti-Shiga toxin (Stx) 2 humanized monoclonal antibody TMA-15 in mice lethally challenged with Stx-producing Escherichia coli. J. Infect. Dis. 2001, 184, 738–742. [Google Scholar] [CrossRef]
- Kimura, T.; Co, M.S.; Vasquez, M.; Wei, S.; Xu, H.; Tani, S.; Sakai, Y.; Kawamura, T.; Matsumoto, Y.; Nakao, H.; et al. Development of humanized monoclonal antibody TMA-15 which neutralizes Shiga toxin 2. Hybrid. Hybridomics 2002, 21, 161–168. [Google Scholar] [CrossRef]
- Lee, L.; Abe, A.; Shayman, J.A. Improved inhibitors of glucosylceramide synthase. J. Biol. Chem. 1999, 274, 14662–14669. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Gregory, S.; Lee, L.; Killen, P.D.; Brady, R.O.; Kulkarni, A.; Shayman, J.A. Reduction of globotriaosylceramide in Fabry disease mice by substrate deprivation. J. Clin. Investig. 2000, 105, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Feitz, W.J.C.; Bouwmeester, R.; van der Velden, T.; Goorden, S.; Licht, C.; van den Heuvel, L.; van de Kar, N. The Shiga Toxin Receptor Globotriaosylceramide as Therapeutic Target in Shiga Toxin E. coli Mediated HUS. Microorganisms 2021, 9, 2157. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.S.; Fischer Sigel, L.K.; Balestracci, A.; Ibarra, C.; Amaral, M.M.; Silberstein, C. Eliglustat prevents Shiga toxin 2 cytotoxic effects in human renal tubular epithelial cells. Pediatr. Res. 2022, 91, 1121–1129. [Google Scholar] [CrossRef]
- Girard, M.C.; Sacerdoti, F.; Rivera, F.P.; Repetto, H.A.; Ibarra, C.; Amaral, M.M. Prevention of renal damage caused by Shiga toxin type 2: Action of Miglustat on human endothelial and epithelial cells. Toxicon 2015, 105, 27–33. [Google Scholar] [CrossRef]
- Amaral, M.M.; Sacerdoti, F.; Jancic, C.; Repetto, H.A.; Paton, A.W.; Paton, J.C.; Ibarra, C. Action of shiga toxin type-2 and subtilase cytotoxin on human microvascular endothelial cells. PLoS ONE 2013, 8, e70431. [Google Scholar] [CrossRef]
- Morace, I.; Pilz, R.; Federico, G.; Jennemann, R.; Krunic, D.; Nordstrom, V.; von Gerichten, J.; Marsching, C.; Schiessl, I.M.; Muthing, J.; et al. Renal globotriaosylceramide facilitates tubular albumin absorption and its inhibition protects against acute kidney injury. Kidney Int. 2019, 96, 327–341. [Google Scholar] [CrossRef]
- Marshall, J.; Nietupski, J.B.; Park, H.; Cao, J.; Bangari, D.S.; Silvescu, C.; Wilper, T.; Randall, K.; Tietz, D.; Wang, B.; et al. Substrate Reduction Therapy for Sandhoff Disease through Inhibition of Glucosylceramide Synthase Activity. Mol. Ther. 2019, 27, 1495–1506. [Google Scholar] [CrossRef]
- Marshall, J.; Sun, Y.; Bangari, D.S.; Budman, E.; Park, H.; Nietupski, J.B.; Allaire, A.; Cromwell, M.A.; Wang, B.; Grabowski, G.A.; et al. CNS-accessible Inhibitor of Glucosylceramide Synthase for Substrate Reduction Therapy of Neuronopathic Gaucher Disease. Mol. Ther. 2016, 24, 1019–1029. [Google Scholar] [CrossRef]
- Viel, C.; Clarke, J.; Kayatekin, C.; Richards, A.M.; Chiang, M.S.R.; Park, H.; Wang, B.; Shihabuddin, L.S.; Sardi, S.P. Preclinical pharmacology of glucosylceramide synthase inhibitor venglustat in a GBA-related synucleinopathy model. Sci. Rep. 2021, 11, 20945. [Google Scholar] [CrossRef]
- Peterschmitt, M.J.; Crawford, N.P.S.; Gaemers, S.J.M.; Ji, A.J.; Sharma, J.; Pham, T.T. Pharmacokinetics, Pharmacodynamics, Safety, and Tolerability of Oral Venglustat in Healthy Volunteers. Clin. Pharmacol. Drug Dev. 2021, 10, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.W.; Shu, L.; Hinkovska-Galcheva, V.; Jin, Y.; Rajeswaran, W.; Abe, A.; Zhao, T.; Luo, R.; Wang, L.; Wen, B.; et al. Optimization of Eliglustat-Based Glucosylceramide Synthase Inhibitors as Substrate Reduction Therapy for Gaucher Disease Type 3. ACS Chem. Neurosci. 2020, 11, 3464–3473. [Google Scholar] [CrossRef] [PubMed]
- Stechmann, B.; Bai, S.K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of retrograde transport protects mice from lethal ricin challenge. Cell 2010, 141, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Secher, T.; Shima, A.; Hinsinger, K.; Cintrat, J.C.; Johannes, L.; Barbier, J.; Gillet, D.; Oswald, E. Retrograde Trafficking Inhibitor of Shiga Toxins Reduces Morbidity and Mortality of Mice Infected with Enterohemorrhagic Escherichia coli. Antimicrob. Agents Chemother. 2015, 59, 5010–5013. [Google Scholar] [CrossRef] [PubMed]
- Forrester, A.; Rathjen, S.J.; Daniela Garcia-Castillo, M.; Bachert, C.; Couhert, A.; Tepshi, L.; Pichard, S.; Martinez, J.; Munier, M.; Sierocki, R.; et al. Functional dissection of the retrograde Shiga toxin trafficking inhibitor Retro-2. Nat. Chem. Biol. 2020, 16, 327–336. [Google Scholar] [CrossRef]
- Gandhi, T.; Patki, M.; Kong, J.; Koya, J.; Yoganathan, S.; Reznik, S.; Patel, K. Development of an Arginine Anchored Nanoglobule with Retrograde Trafficking Inhibitor (Retro-2) for the Treatment of an Enterohemorrhagic Escherichia coli Outbreak. Mol. Pharm. 2019, 16, 4405–4415. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Identifier | Title | Interventions | Time Period | Country | Reference |
|---|---|---|---|---|---|
| NCT00004465 | Phase III Randomized Study of SYNSORB Pk in Children with E. coli-Associated Hemolytic Uremic Syndrome | SYNSORB Pk vs. placebo | 1997/7/27~2001/4/14 | United States, Canada | [111,112,113] |
| SHIGATEC | A Phase II Study Assessing Monoclonal Antibodies Against Shiga toxin 1 and 2 in Shiga toxin-producing E. coli-infected Children | Shigamabs(cαStx1/cαStx2) vs. placebo | 2010/11~2011/2 | Argentina, Chile, Peru | [113,114,115,116] |
| NCT01252199 | Study of Chimeric Monoclonal Antibodies to Shiga Toxins 1 and 2 | cαStx1/cαStx2 vs. placebo | 2011/11~2013/2 | Argentina, Chile, Peru | [113,114,115,116] |
| NCT01406288 | Outbreak of Hemolytic Uremic Syndrome Linked to Escherichia coli of Serotype O104:H4 (SHU O104 CUB) | HUS standard coverage care (including in ICU) | 2011/7~2012/3 | France | |
| NCT01410916 | Safety and Efficacy Study of Eculizumab in Shiga-Toxin Producing Escherichia coli Hemolytic-Uremic Syndrome (STEC-HUS) | Eculizumab (Soliris®) vs. placebo | 2011/7~2012/6 | Germany | |
| NCT02205541 | Eculizumab in Shiga-toxin Related Hemolytic and Uremic Syndrome Pediatric Patients (ECULISHU) | Eculizumab vs. placebo | 2015/6~2018/6 | France | [113,117] |
| NCT02336516 | Azithromycin in Post Diarrheal Haemolytic and Uremic Syndrome (ZITHROSHU) | Azithromycin vs. placebo | 2015/7~2021/4 | France | |
| ECUSTEC | ECUlizumab in Shiga-Toxin producing Escherichia coli Haemolytic Uraemic Syndrome (ECUSTEC): A Randomised, Double-Blind, Placebo-Controlled Trial | Eculizumab vs. placebo | 2016/12~2021/4 | United Kingdom | |
| NCT03388216 | Anti-Shiga Toxin Hyperimmune Equine Immunoglobulin F(ab’)2 Fragment (INM004) in Healthy Volunteers | INM004 vs. placebo | 2017/12/16~2018/9/28 | Argentina | [118] |
| NCT03776851 | Erythropoietin in Hemolytic Uremic Syndrome | Erythropoietin (EPO) | 2019/1/1~2020/12/30 | Argentina | [119,120,121] |
| NCT04132375 | Phase 2/3 Study to Evaluate PK, Safety, and Efficacy of INM004 in STEC Positive Pediatric Patients for Prevention of HUS | INM004 vs. placebo | 2019/7/17~2022/9/1 | Argentina | [113,118] |
| NCT05219110 | Hyperhydration in Children with Shiga Toxin-Producing E. coli Infection (HIKO STEC) | Hyperhydration vs. conservative fluid management | 2022/9/1~2027/8/31 | United States, Canada |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Thaker, H.; Wang, C.; Xu, Z.; Dong, M. Diagnosis and Treatment for Shiga Toxin-Producing Escherichia coli Associated Hemolytic Uremic Syndrome. Toxins 2023, 15, 10. https://doi.org/10.3390/toxins15010010
Liu Y, Thaker H, Wang C, Xu Z, Dong M. Diagnosis and Treatment for Shiga Toxin-Producing Escherichia coli Associated Hemolytic Uremic Syndrome. Toxins. 2023; 15(1):10. https://doi.org/10.3390/toxins15010010
Chicago/Turabian StyleLiu, Yang, Hatim Thaker, Chunyan Wang, Zhonggao Xu, and Min Dong. 2023. "Diagnosis and Treatment for Shiga Toxin-Producing Escherichia coli Associated Hemolytic Uremic Syndrome" Toxins 15, no. 1: 10. https://doi.org/10.3390/toxins15010010
APA StyleLiu, Y., Thaker, H., Wang, C., Xu, Z., & Dong, M. (2023). Diagnosis and Treatment for Shiga Toxin-Producing Escherichia coli Associated Hemolytic Uremic Syndrome. Toxins, 15(1), 10. https://doi.org/10.3390/toxins15010010