Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex

 ,
,  and
and
Abstract
:1. Introduction
2. Results
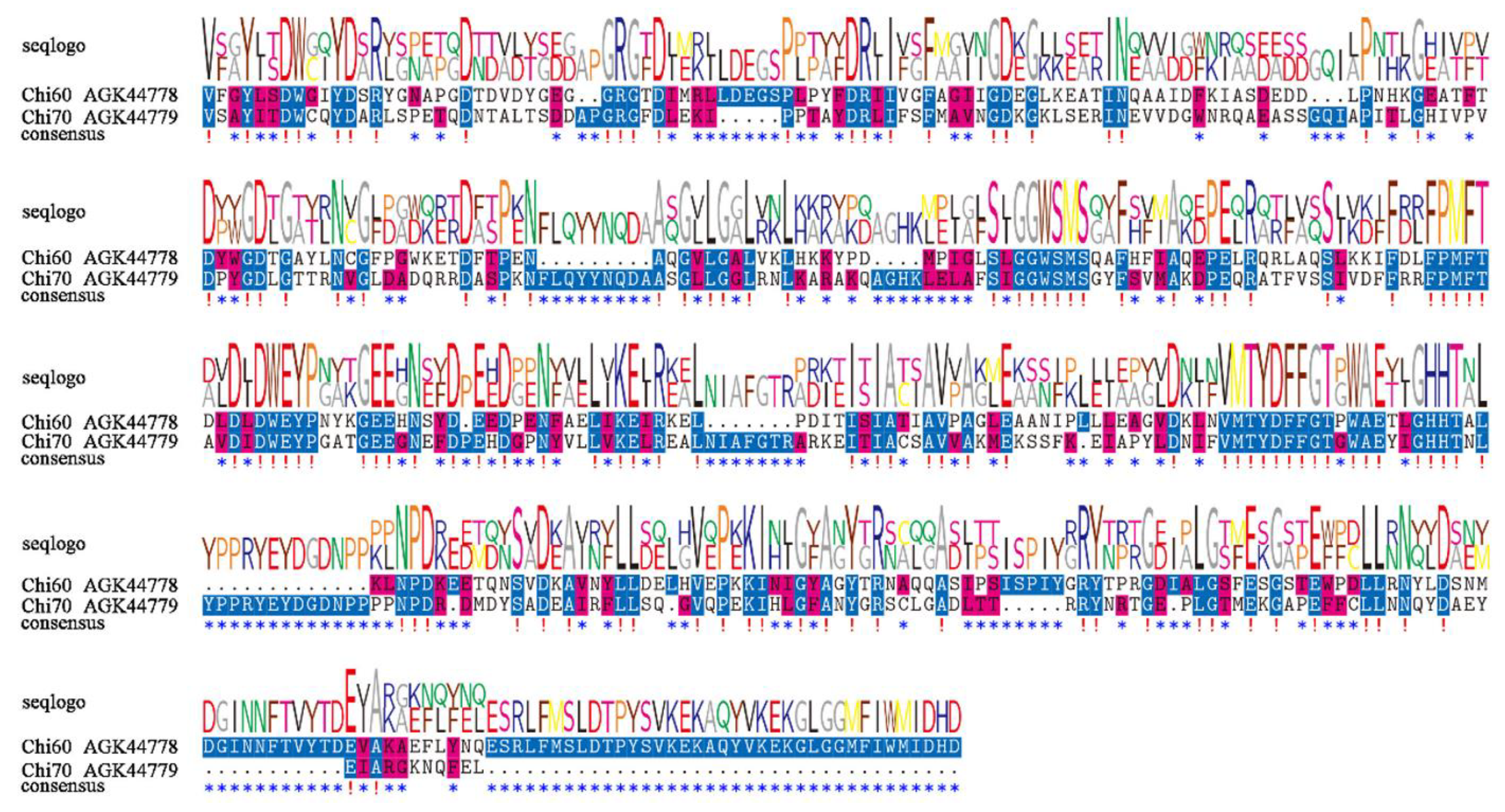
2.1. Sequence Analysis of Chitinases
2.2. Insecticidal Activity of Chitinases and Synergistic Effect with Tc Toxin
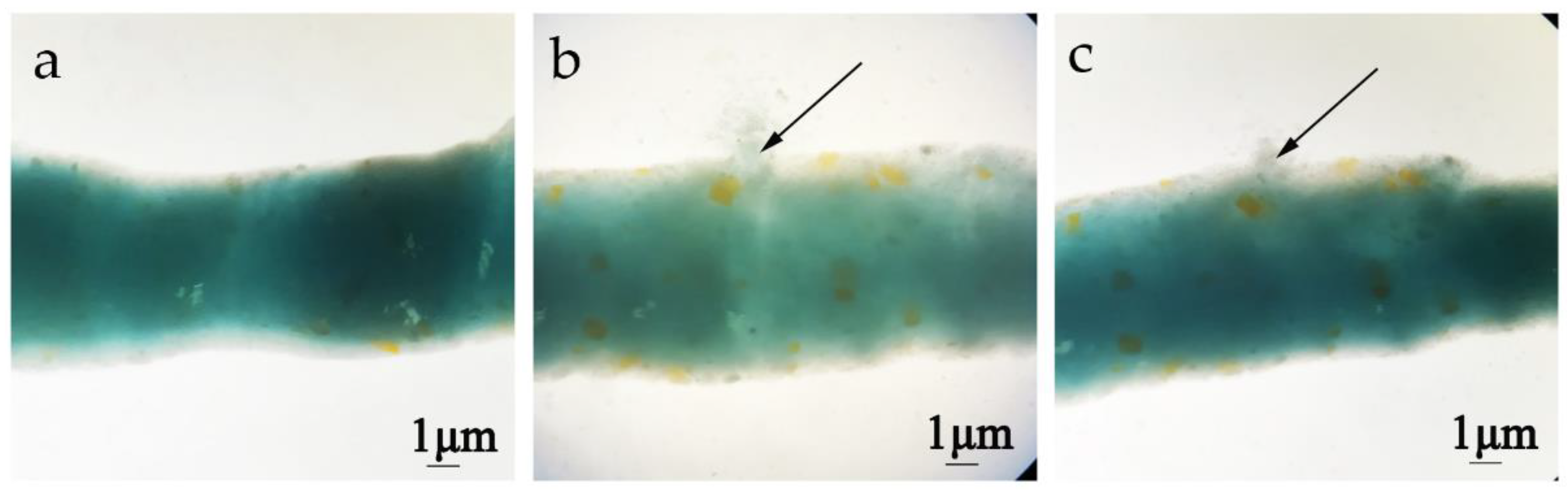
2.3. Pathological Effect of Chitinases on the Peritrophic Membrane of H. armigera
2.4. Homologous Recombination Vector Construction
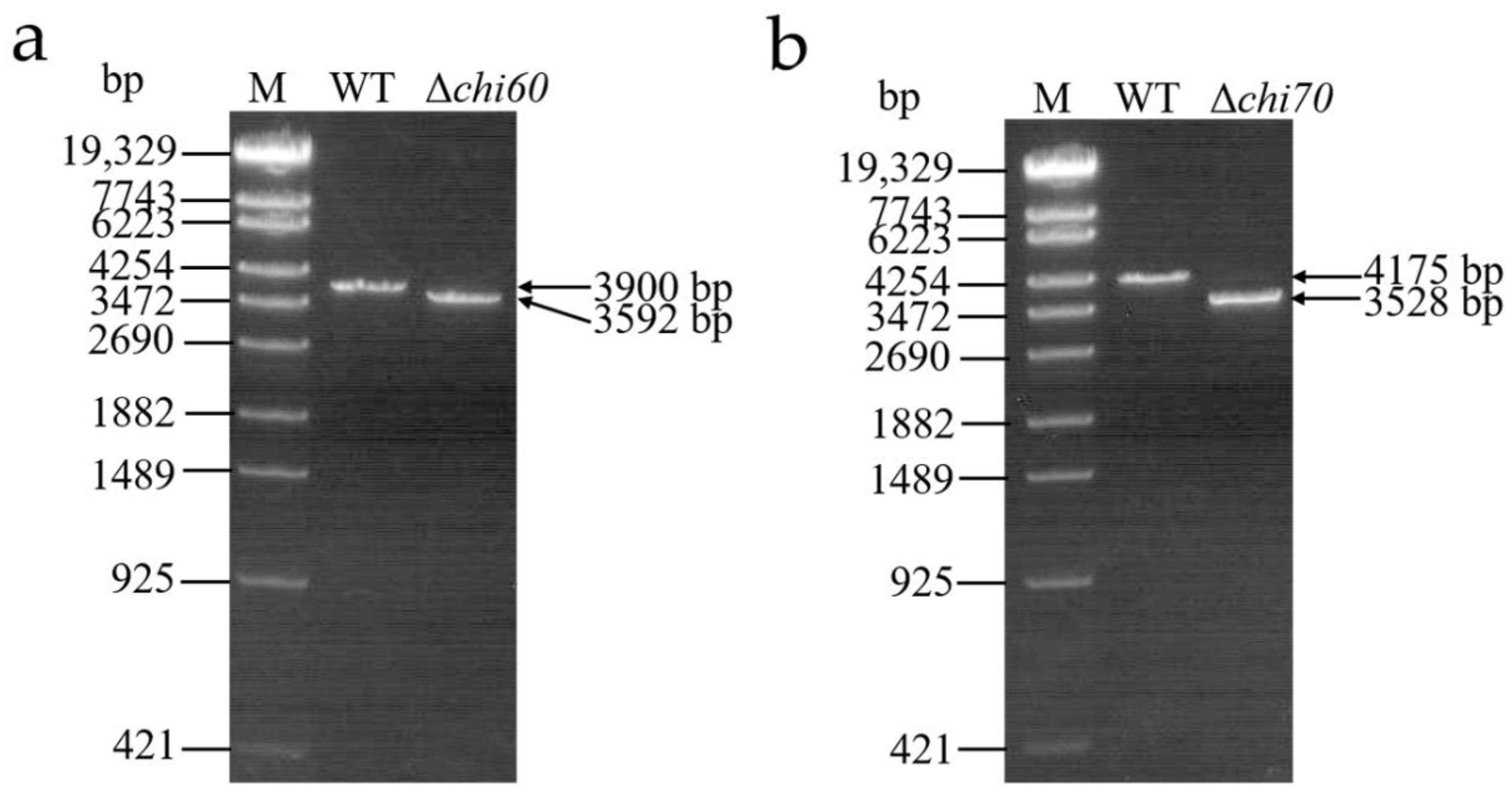
2.5. Identification of Single Gene Knockout Mutants
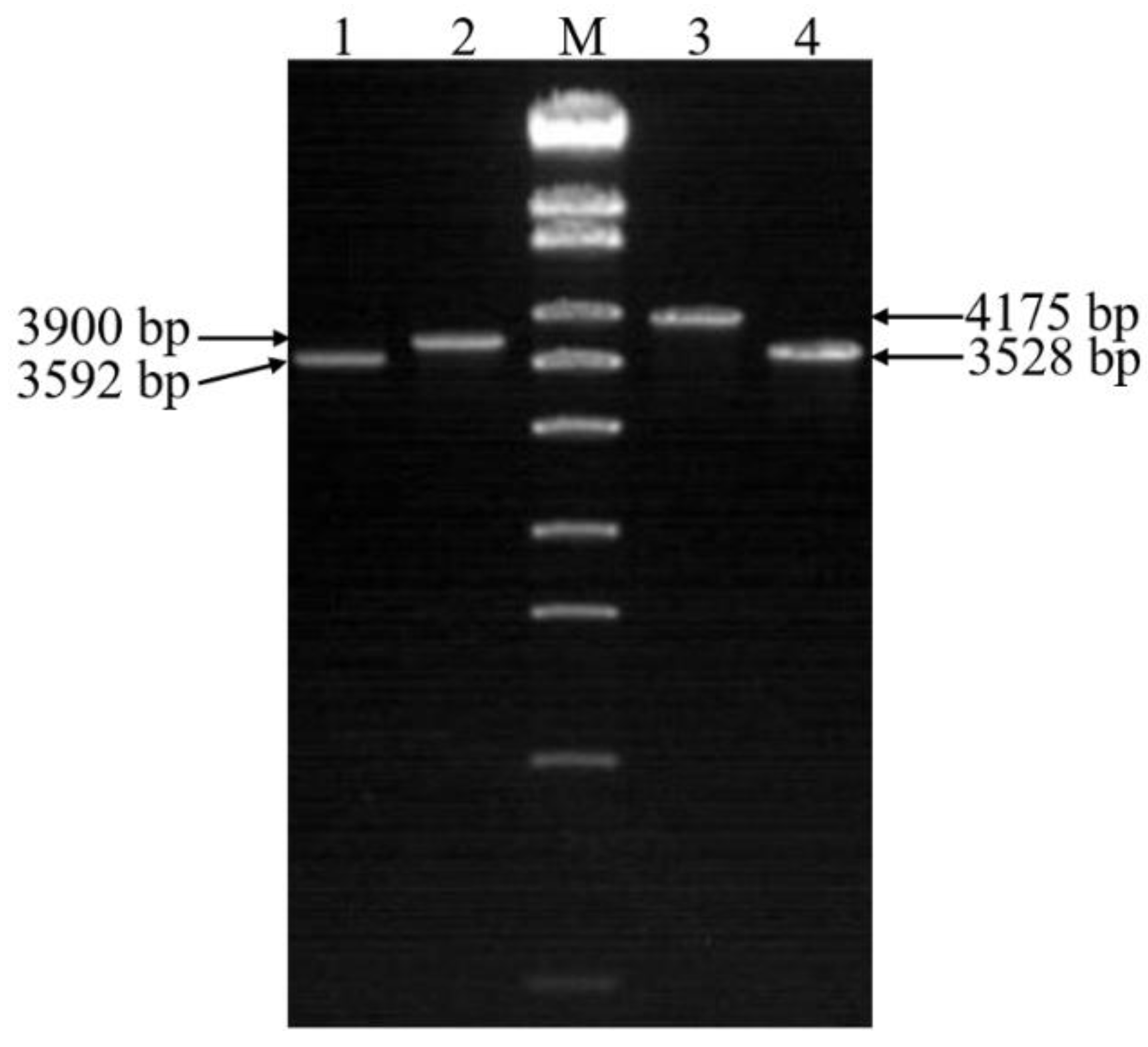
2.6. Identification of Double Gene Knockout Mutants
2.7. Western Blot Analysis
2.8. Insecticidal Activity of Tc Toxins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insects, Microorganisms and Proteins
5.2. Sequence Analysis of Chitinase
5.3. Assay for Insecticidal Activity of Chitinases and Pathological Effect
5.3.1. Assay for Insecticidal Activity of Chitinase
5.3.2. Pathological Effect of Chitinase on the Peritrophic Membrane
5.4. Knockout of the Chitinase Gene from X. nematophila HB310
5.4.1. Genomic DNA Extraction
5.4.2. Homologous Recombination Vector Construction
5.4.3. Single Gene Knockout Mutants Screening
5.4.4. Double Gene Knockout Mutant Screening
5.5. Western Blot
5.6. Assay for Insecticidal Activity of Tc Toxins
5.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Shen, N.; Wu, J.; Jiang, M.; Shi, S.; Wang, J.; Wei, Y.; Yang, L. Cloning, expression and characterization of a chitinase from Paenibacillus chitinolyticus strain UMBR 0002. PeerJ 2020, 8, e8964. [Google Scholar] [CrossRef] [PubMed]
- Paek, A.; Kim, M.J.; Park, H.Y.; Yoo, J.G.; Jeong, S.E. Functional expression of recombinant hybrid enzymes composed of bacterial and insect’s chitinase domains in E. coli. Enzyme Microb. Technol. 2020, 136, 109492. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zheng, J.; Liang, Y.; Yan, R.; Xu, X.; Lin, J. Expression and characterization of a chitinase from Serratia marcescens. Protein Expr. Purif. 2020, 171, 105613. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Kim, J.; Son, K.H.; Chung, C.W.; Shin, D.H.; Ku, B.H.; Kim, D.Y.; Park, H.Y. Novel Bi-Modular GH19 chitinase with broad pH stability from a fibrolytic intestinal symbiont of Eisenia fetida, Cellulosimicrobium funkei HY-13. Biomolecules 2021, 11, 1735. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, S.; Kumar, M.; Kumari, P.; Mahapatro, G.K.; Banerjee, N.; Sarin, N.B. Novel insecticidal chitinase from the insect pathogen Xenorhabdus nematophila. Int. J. Biol. Macromol. 2020, 159, 394–401. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, W.; Zhang, X.; Huang, J.; Wang, W.; Miao, M.; Hu, L.; Wan, C.; Yuan, Y.; Wu, B.; et al. Genome-wide identification and expression analysis of chitinase-like genes in Petunia axillaris. Plants 2022, 11, 1269. [Google Scholar] [CrossRef]
- Zhang, Y.; Guan, F.; Xu, G.; Liu, X.; Zhang, Y.; Sun, J.; Yao, B.; Huang, H.; Wu, N.; Tian, J. A novel thermophilic chitinase directly mined from the marine metagenome using the deep learning tool Preoptem. Bioresour. Bioprocess 2022, 9, 54. [Google Scholar] [CrossRef]
- Arora, N.; Ahmad, T.; Rajagopal, R.; Bhatnagar, R.K. A constitutively expressed 36 kDa exochitinase from Bacillus thuringiensis HD-1. Biochem. Biophys. Res. Commun. 2003, 307, 620–625. [Google Scholar] [CrossRef]
- Liu, J.; Nangong, Z.; Zhang, J.; Song, P.; Tang, Y.; Gao, Y.; Wang, Q. Expression and characterization of two chitinases with synergistic effect and antifungal activity from Xenorhabdus nematophila. World J. Microbiol. Biotechnol. 2019, 35, 106. [Google Scholar] [CrossRef]
- Liu, M.; Cai, Q.X.; Liu, H.Z.; Zhang, B.H.; Yan, J.P.; Yuan, Z.M. Chitinolytic activities in Bacillus thuringiensis and their synergistic effects on larvicidal activity. J. Appl. Microbiol. 2002, 93, 374–379. [Google Scholar]
- Bravo, A.; Soberon, M. How to cope with insect resistance to Bt toxins? Trends Biotechnol. 2008, 26, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Leidreiter, F.; Roderer, D.; Meusch, D.; Gatsogiannis, C.; Benz, R.; Raunser, S. Common architecture of Tc toxins from human and insect pathogenic bacteria. Sci. Adv. 2019, 5, x6497. [Google Scholar] [CrossRef]
- Fuchs, T.M.; Bresolin, G.; Marcinowski, L.; Schachtner, J.; Scherer, S. Insecticidal genes of Yersinia spp.: Taxonomical distribution, contribution to toxicity towards Manduca sexta and Galleria mellonella, and evolution. BMC Microbiol. 2008, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, M.J.; Jones, S.A.; Rothnagel, R.; Busby, J.N.; Marshall, S.D.; Simpson, R.M.; Lott, J.S.; Hankamer, B.; Hurst, M.R. 3D structure of the Yersinia entomophaga toxin complex and implications for insecticidal activity. Proc. Natl. Acad. Sci. USA 2011, 108, 20544–20549. [Google Scholar] [CrossRef] [PubMed]
- Busby, J.N.; Landsberg, M.J.; Simpson, R.M.; Jones, S.A.; Hankamer, B.; Hurst, M.R.; Lott, J.S. Structural analysis of Chi1 chitinase from Yen-Tc: The multisubunit insecticidal ABC toxin complex of Yersinia entomophaga. J. Mol. Biol. 2012, 415, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhang, J.; Li, T.; Liu, S.; Song, P.; Nangong, Z.; Wang, Q. PirAB protein from Xenorhabdus nematophila HB310 exhibits a binary toxin with insecticidal activity and cytotoxicity in Galleria mellonella. J. Invertebr. Pathol. 2017, 148, 43–50. [Google Scholar] [CrossRef]
- Liu, J.; Song, P.; Zhang, J.; Nangong, Z.; Liu, X.; Gao, Y.; Wang, Q. Characteristics and function of the chitin binding protein from Xenorhabdus nematophila. Protein Peptide Lett. 2019, 26, 414–422. [Google Scholar] [CrossRef]
- Sergeant, M.; Jarrett, P.; Ousley, M.; Morgan, J.A. Interactions of insecticidal toxin gene products from Xenorhabdus nematophilus PMFI296. Appl. Environ. Microbiol. 2003, 69, 3344–3349. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2007, 49, 436–451. [Google Scholar] [CrossRef]
- Waterfield, N.; Hares, M.; Hinchliffe, S.; Wren, B.; Ffrench-Constant, R. The insect toxin complex of Yersinia. Adv. Exp. Med. Biol. 2007, 603, 247–257. [Google Scholar]
- Lang, A.E.; Konukiewitz, J.; Aktories, K.; Benz, R. TcdA1 of Photorhabdus luminescens: Electrophysiological analysis of pore formation and effector binding. Biophys. J. 2013, 105, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Gatsogiannis, C.; Lang, A.E.; Meusch, D.; Pfaumann, V.; Hofnagel, O.; Benz, R.; Aktories, K.; Raunser, S. A syringe-like injection mechanism in Photorhabdus luminescens toxins. Nature 2013, 495, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Gatsogiannis, C.; Merino, F.; Prumbaum, D.; Roderer, D.; Leidreiter, F.; Meusch, D.; Raunser, S. Membrane insertion of a Tc toxin in near-atomic detail. Nat. Struct. Mol. Biol. 2016, 23, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Gatsogiannis, C.; Merino, F.; Roderer, D.; Balchin, D.; Schubert, E.; Kuhlee, A.; Hayer-Hartl, M.; Raunser, S. Tc toxin activation requires unfolding and refolding of a beta-propeller. Nature 2018, 563, 209–213. [Google Scholar] [CrossRef]
- Meusch, D.; Gatsogiannis, C.; Efremov, R.G.; Lang, A.E.; Hofnagel, O.; Vetter, I.R.; Aktories, K.; Raunser, S. Mechanism of Tc toxin action revealed in molecular detail. Nature 2014, 508, 61–65. [Google Scholar] [CrossRef]
- Roderer, D.; Schubert, E.; Sitsel, O.; Raunser, S. Towards the application of Tc toxins as a universal protein translocation system. Nat. Commun. 2019, 10, 5263. [Google Scholar] [CrossRef]
- Sheets, J.J.; Hey, T.D.; Fencil, K.J.; Burton, S.L.; Ni, W.; Lang, A.E.; Benz, R.; Aktories, K. Insecticidal toxin complex proteins from Xenorhabdus nematophilus: Structure and pore formation. J. Biol. Chem. 2011, 286, 22742–22749. [Google Scholar] [CrossRef]
- Roderer, D.; Raunser, S. Tc toxin complexes: Assembly, membrane permeation, and protein translocation. Annu. Rev. Microbiol. 2019, 73, 247–265. [Google Scholar] [CrossRef]
- Morgan, J.A.; Sergeant, M.; Ellis, D.; Ousley, M.; Jarrett, P. Sequence analysis of insecticidal genes from Xenorhabdus nematophilus PMFI296. Appl. Environ. Microbiol. 2001, 67, 2062–2069. [Google Scholar] [CrossRef]
- Hurst, M.R.; Glare, T.R.; Jackson, T.A.; Ronson, C.W. Plasmid-located pathogenicity determinants of Serratia entomophila, the causal agent of amber disease of grass grub, show similarity to the insecticidal toxins of Photorhabdus luminescens. J. Bacteriol. 2000, 182, 5127–5138. [Google Scholar] [CrossRef]
- Hurst, M.R.; Jones, S.M.; Tan, B.; Jackson, T.A. Induced expression of the Serratia entomophila Sep proteins shows activity towards the larvae of the New Zealand grass grub Costelytra zealandica. Fems. Microbiol. Lett. 2007, 275, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Stoilova-Mcphie, S.; Baxter, L.; Fulop, V.; Henderson, J.; Rodger, A.; Roper, D.I.; Scott, D.J.; Smith, C.J.; Morgan, J.A. Structural characterisation of the insecticidal toxin XptA1, reveals a 1.15 MDa tetramer with a cage-like structure. J. Mol. Biol. 2007, 366, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Nangong, Z.Y.; Yang, J.; Song, P.; Wang, Y.; Cui, L.; Cui, L. Toxic activity of a protein complex purified from Xenorhabdus nematophila HB310 to Plutella xylostella larvae. Insect Sci. 2012, 19, 329–336. [Google Scholar] [CrossRef]
- Chandrasekaran, R.; Revathi, K.; Thanigaivel, A.; Kirubakaran, S.A.; Senthil-Nathan, S. Bacillus subtilis chitinase identified by matrix-assisted laser desorption/ionization time-of flight/time of flight mass spectrometry has insecticidal activity against Spodoptera litura Fab. Pestic. Biochem. Physiol. 2014, 116, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Suganthi, M.; Senthilkumar, P.; Arvinth, S.; Chandrashekara, K.N. Chitinase from Pseudomonas fluorescens and its insecticidal activity against Helopeltis theivora. J. Gen. Appl. Microbiol. 2017, 63, 222–227. [Google Scholar] [CrossRef]
- Okongo, R.N.; Puri, A.K.; Wang, Z.; Singh, S.; Permaul, K. Comparative biocontrol ability of chitinases from bacteria and recombinant chitinases from the thermophilic fungus Thermomyces lanuginosus. J. Biosci. Bioeng. 2019, 127, 663–671. [Google Scholar] [CrossRef]
- Subbanna, A.; Chandrashekara, C.; Stanley, J.; Mishra, K.K.; Mishra, P.K.; Pattanayak, A. Bio-efficacy of chitinolytic Bacillus thuringiensis isolates native to northwestern Indian Himalayas and their synergistic toxicity with selected insecticides. Pestic. Biochem. Physiol. 2019, 158, 166–174. [Google Scholar] [CrossRef]
- Wiwat, C.; Thaithanun, S.; Pantuwatana, S.; Bhumiratana, A. Toxicity of chitinase-producing Bacillus thuringiensis ssp. kurstaki HD-1 (G) toward Plutella xylostella. J. Invertebr. Pathol. 2000, 76, 270–277. [Google Scholar]
- Lertcanawanichakul, M.; Wiwat, C.; Bhumiratana, A.; Dean, D.H. Expression of chitinase-encoding genes in Bacillus thuringiensis and toxicity of engineered B. thuringiensis subsp. aizawai toward Lymantria dispari larvae. Curr. Microbiol. 2004, 48, 175–181. [Google Scholar] [CrossRef]
- Liu, D.; Cai, J.; Xie, C. Purification and partial characterization of a 36-kDa chitinase from Bacillus thuringiensis subsp. colmeri, and its biocontrol potential. Enzyme. Microb. Tech. 2010, 46, 252–256. [Google Scholar] [CrossRef]
- Hurst, M.R.; Jones, S.A.; Binglin, T.; Harper, L.A.; Jackson, T.A.; Glare, T.R. The main virulence determinant of Yersinia entomophaga MH96 is a broad-host-range toxin complex active against insects. J. Bacteriol. 2011, 193, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Nagpure, A.; Gupta, R.K. Bacterial chitinases: Properties and potential. Crit. Rev. Biotechnol. 2007, 27, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cao, Y.; Zhang, W.; Liu, Z.; Li, Y.; Chen, Y.; Zhang, H.; Yu, F.; Liu, X. The wheat TaIQD3D-6 gene encodes a microtubule-associated protein and regulates cell morphogenesis in Arabidopsis. Plant. Sci. 2022, 324, 111420. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tang, Y.; Wang, Q.; Shi, H.; Yin, J.; Li, C. Identification and characterization of an antennae-specific glutathione S-transferase from the Indian Meal Moth. Front. Physiol. 2021, 12, 727619. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, K.; Jing, C.; Wu, R.; Wu, G.; Li, M.; Qing, L. Molecular characterization of a novel Conyza canadensis-infecting begomovirus in China. Phytopathol. Res. 2022, 4, 13. [Google Scholar] [CrossRef]
- Ajit, N.S.; Verma, R.; Shanmugam, V. Extracellular chitinases of fluorescent pseudomonads antifungal to Fusarium oxysporum f. Sp. dianthi causing carnation wilt. Curr. Microbiol. 2006, 52, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Kirubakaran, S.I.; Sakthivel, N. Cloning and overexpression of antifungal barley chitinase gene in Escherichia coli. Protein Expr. Purif. 2007, 52, 159–166. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Growth Inhibition Rate (%) | Corrected Mortality (%) |
|---|---|---|
| Chi60 | 81.99 ± 2.42 | 4.17 ± 2.41 |
| Chi70 | 90.51 ± 1.44 * | 13.89 ± 1.39 * |
| Treatment | LC50 (μg/mL) | 95% CL | Slope ± SE | R2 |
|---|---|---|---|---|
| Tc toxins | 196.11 | 149.30–251.30 | 2.37 ± 0.33 | 0.99 |
| Tc toxins + Chi60 | 185.85 | 151.75–224.32 | 3.00 ± 0.34 | 0.98 |
| Tc toxins + Chi70 | 146.47 | 113.14–175.84 | 2.52 ± 0.31 | 0.99 |
| Treatment | LC50 (μg/mL) | 95% CL | Slope ± SE | R2 |
|---|---|---|---|---|
| WT | 196.11 | 149.30–251.30 | 2.37 ± 0.33 | 0.99 |
| Δchi60 | 885.74 | 650.53–1342.22 | 1.68 ± 0.28 | 0.99 |
| Δchi70 | 757.25 | 551.29–1144.21 | 1.54 ± 0.22 | 0.99 |
| Δchi60-chi70 | 20,049.83 | 10,711.52–64,351.57 | 0.80 ± 0.16 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Bai, H.; Song, P.; Nangong, Z.; Dong, Z.; Li, Z.; Wang, Q. Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex. Toxins 2022, 14, 646. https://doi.org/10.3390/toxins14090646
Liu J, Bai H, Song P, Nangong Z, Dong Z, Li Z, Wang Q. Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex. Toxins. 2022; 14(9):646. https://doi.org/10.3390/toxins14090646
Chicago/Turabian StyleLiu, Jia, Hui Bai, Ping Song, Ziyan Nangong, Zhiping Dong, Zhiyong Li, and Qinying Wang. 2022. "Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex" Toxins 14, no. 9: 646. https://doi.org/10.3390/toxins14090646
APA StyleLiu, J., Bai, H., Song, P., Nangong, Z., Dong, Z., Li, Z., & Wang, Q. (2022). Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex. Toxins, 14(9), 646. https://doi.org/10.3390/toxins14090646