Staphylococcus aureus Alpha-Toxin in Deep Tracheal Aspirates—Preliminary Evidence for Its Presence in the Lungs of Sepsis Patients


 ,
,
Abstract
1. Introduction
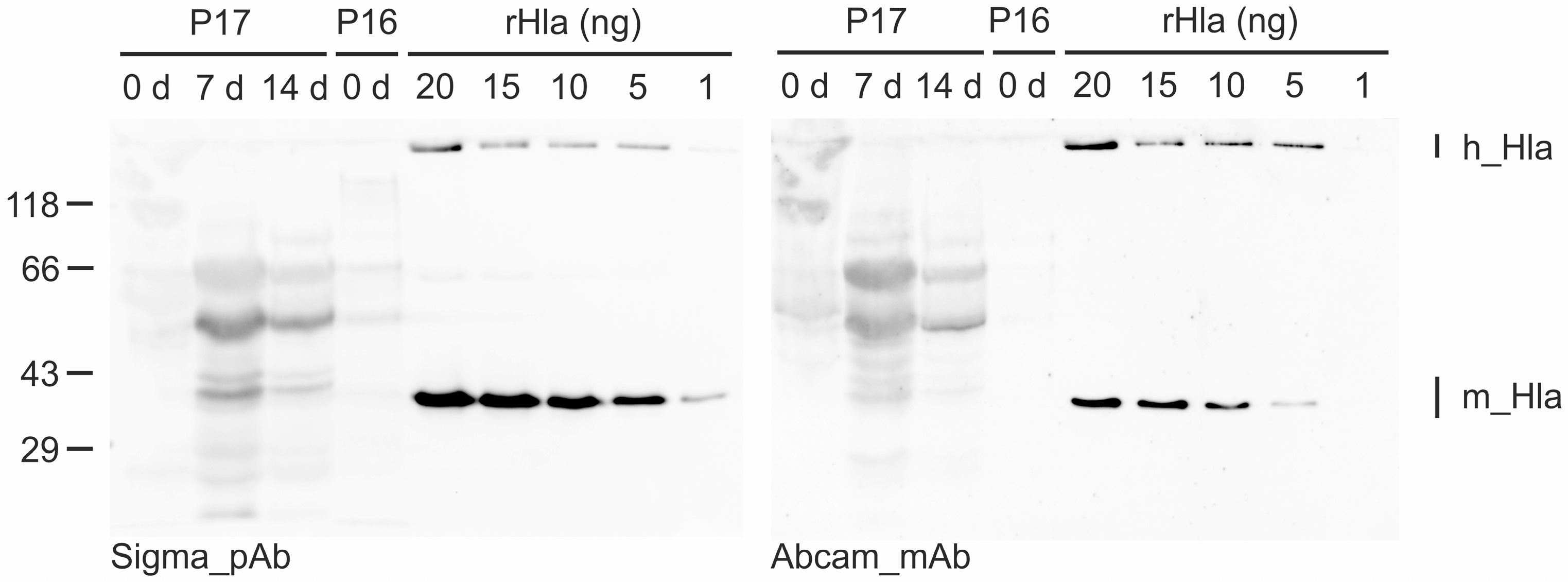
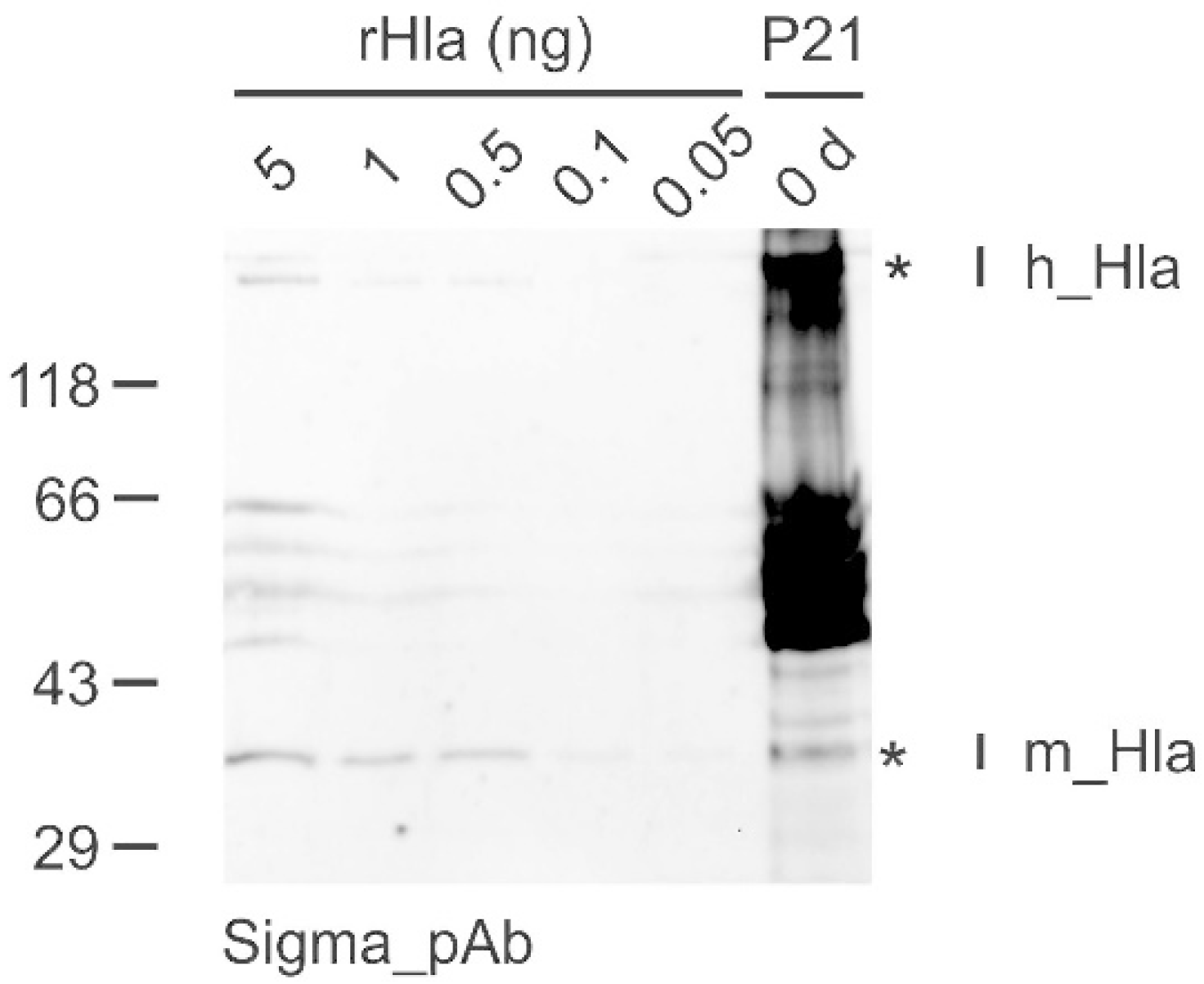
2. Results
3. Discussion
4. Materials and Methods
4.1. Deep Tracheal Aspirates (DTA)
4.2. Sample Preparation for the Blood Culture of Microorganisms
4.3. Antibodies
4.4. Recombinant Hla (rHla)
4.5. Western Blot Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- McCarthy, H.; Rudkin, J.K.; Black, N.S.; Gallagher, L.; O’Neill, E.; O’Gara, J.P. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2015, 5, 1. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef]
- da Silva, M.C.; Zahm, J.M.; Gras, D.; Bajolet, O.; Abely, M.; Hinnrasky, J.; Milliot, M.; De Assis, M.C.; Hologne, C.; Bonnet, N.; et al. Dynamic interaction between airway epithelial cells and Staphylococcus aureus. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2004, 287, L543–L551. [Google Scholar] [CrossRef]
- Gouaux, E. alpha-Hemolysin from Staphylococcus aureus: An archetype of beta-barrel, channel-forming toxins. J. Struct. Biol. 1998, 121, 110–122. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P. Pore-forming virulence factors of Staphylococcus aureus destabilize epithelial barriers—Effects of alpha-toxin in the early phases of airway infection. AIMS Microbiol. 2015, 1, 11–36. [Google Scholar] [CrossRef]
- von Hoven, G.; Qin, Q.; Neukirch, C.; Husmann, M.; Hellmann, N. Staphylococcus aureus alpha-toxin: Small pore, large consequences. Biol. Chem. 2019, 400, 1261–1276. [Google Scholar] [CrossRef]
- Walev, I.; Martin, E.; Jonas, D.; Mohamadzadeh, M.; Muller-Klieser, W.; Kunz, L.; Bhakdi, S. Staphylococcal alpha-toxin kills human keratinocytes by permeabilizing the plasma membrane for monovalent ions. Infect. Immun. 1993, 61, 4972–4979. [Google Scholar] [CrossRef]
- Rose, F.; Dahlem, G.; Guthmann, B.; Grimminger, F.; Maus, U.; Hanze, J.; Duemmer, N.; Grandel, U.; Seeger, W.; Ghofrani, H.A. Mediator generation and signaling events in alveolar epithelial cells attacked by S. aureus alpha-toxin. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L207–L214. [Google Scholar] [CrossRef]
- Eiffler, I.; Behnke, J.; Ziesemer, S.; Müller, C.; Hildebrandt, J.-P. Staphylococcus aureus alpha-toxin-mediated cation entry depolarizes membrane potential and activates p38 MAP kinase in airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L676–L685. [Google Scholar] [CrossRef]
- Räth, S.; Ziesemer, S.; Witte, A.; Konkel, A.; Müller, C.; Hildebrandt, P.; Völker, U.; Hildebrandt, J.-P. S. aureus hemolysin A-induced IL-8 and IL-6 release from human airway epithelial cells is mediated by activation of p38- and Erk-MAP kinases and additional, cell-type specific signalling mechanisms. Cell. Microbiol. 2013, 15, 1253–1265. [Google Scholar] [CrossRef]
- Hermann, I.; Räth, S.; Ziesemer, S.; Volksdorf, T.; Dress, R.J.; Gutjahr, M.; Müller, C.; Beule, A.G.; Hildebrandt, J.-P. Staphylococcus aureus-hemolysin A disrupts cell-matrix adhesions in human airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2015, 52, 14–24. [Google Scholar] [CrossRef]
- Ziesemer, S.; Eiffler, I.; Schönberg, A.; Müller, C.; Hochgräfe, F.; Beule, A.G.; Hildebrandt, J.-P. Staphylococcus aureus alpha-toxin induces actin filament remodeling in human airway epithelial model cells. Am. J. Respir. Cell Mol. Biol. 2018, 58, 482–491. [Google Scholar] [CrossRef]
- Berube, B.J.; BubeckWardenburg, J. Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Ajao, A.O.; Aman, M.J.; Karauzum, H.; Sarwar, J.; Lydecker, A.D.; Johnson, J.K.; Nguyen, C.; Chen, W.H.; Roghmann, M.-C. Lower antibody levels to Staphylococcus aureus exotoxins are associated with sepsis in hospitalized adults with invasive S. aureus infections. J. Infect. Dis. 2012, 206, 915–923. [Google Scholar] [CrossRef]
- Monecke, S.; Müller, E.; Büchler, J.; Stieber, B.; Ehricht, R. Staphylococcus aureus in vitro secretion of alpha toxin (hla) correlates with the affiliation to clonal complexes. PLoS ONE 2014, 9, e100427. [Google Scholar] [CrossRef]
- Tavares, A.; Nielsen, J.B.; Boye, K.; Rohde, S.; Paulo, A.C.; Westh, H.; Schønning, K.; De Lencastre, H.; Miragaia, M. Insights into alpha-hemolysin (Hla) evolution and expression among Staphylococcus aureus clones with hospital and community origin. PLoS ONE 2014, 9, e98634. [Google Scholar] [CrossRef]
- BubeckWardenburg, J.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar] [CrossRef]
- Foletti, D.; Strop, P.; Shaughnessy, L.; Hasa-Moreno, A.; Casas, M.G.; Russell, M.; Bee, C.; Wu, S.; Pham, A.; Zeng, Z.; et al. Mechanism of action and in vivo efficacy of a human-derived antibody against Staphylococcus aureus alpha-hemolysin. J. Mol. Biol. 2013, 425, 1641–1654. [Google Scholar] [CrossRef]
- Fritz, S.A.; Tiemann, K.M.; Hogan, P.G.; Epplin, E.K.; Rodriguez, M.; Al-Zubeidi, D.N.; Bubeck Wardenburg, J.; Hunstad, D.A. A serologic correlate of protective immunity against community-onset Staphylococcus aureus infection. Clin. Infect. Dis. 2013, 56, 1554–1561. [Google Scholar] [CrossRef]
- Fischer, A.J.; Singh, S.B.; LaMarche, M.M.; Maakestad, L.J.; Kienenberger, Z.E.; Peña, T.A.; Stoltz, D.A.; Limoli, D.H. Sustained coinfections with Staphylococcus aureus and Pseudomonas aeruginosa in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2020, 203, 328–338. [Google Scholar] [CrossRef]
- Hughes, D.A.; Price, H.; Rosenthal, M.; Davies, J.C. Pseudomonas aeruginosa in the CF airway: Does it deserve its reputation as a predatory ‘bully’? Am. J. Respir. Crit. Care Med. 2021, 203, 1027–1030. [Google Scholar] [CrossRef]
- Metersky, M.L.; Aksamit, T.R.; Barker, A.; Choate, R.; Daley, C.L.; Daniels, L.A.; DiMango, A.; Eden, E.; Griffith, D.; Johnson, M.; et al. The prevalence and significance of Staphylococcus aureus in patients with non-cystic fibrosis bronchiectasis. Ann. Am. Thoracic Soc. 2018, 15, 365–370. [Google Scholar] [CrossRef]
- Montoya, M.; Gouaux, E. Beta-barrel membrane protein folding and structure viewed through the lens of alpha-hemolysin. Biochim. Biophys. Acta 2003, 1609, 19–27. [Google Scholar] [CrossRef]
- Wilke, G.A.; BubeckWardenburg, J. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus alpha-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13578. [Google Scholar] [CrossRef]
- Möller, N.; Ziesemer, S.; Hildebrandt, P.; Assenheimer, N.; Völker, U.; Hildebrandt, J.-P. S. aureus alpha-toxin monomer binding and heptamer formation in host cell membranes—Do they determine sensitivity of airway epithelial cells toward the toxin? PLoS ONE 2020, 15, e0233854. [Google Scholar] [CrossRef]
- Ziesemer, S.; Möller, N.; Nitsch, A.; Müller, C.; Beule, A.G.; Hildebrandt, J.-P. Sphingomyelin depletion from plasma membranes of human airway epithelial cells completely abrogates the deleterious actions of S. aureus alpha-toxin. Toxins 2019, 11, 126. [Google Scholar] [CrossRef]
- Scheer, C.S.; Fuchs, C.; Gründling, M.; Vollmer, M.; Bast, J.; Bohnert, J.A.; Zimmermann, K.; Hahnenkamp, K.; Rehberg, S.; Kuhn, S.O. Impact of antibiotic administration on blood culture positivity at the beginning of sepsis: A prospective clinical cohort study. Clin. Microbiol. Infect. 2019, 25, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Below, S.; Konkel, A.; Zeeck, C.; Müller, C.; Kohler, C.; Engelmann, S.; Hildebrandt, J.-P. Virulence factors of Staphylococcus aureus induce Erk-MAP kinase activation and c-Fos expression in S9 and 16HBE14o- human airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L470–L479. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Patient # | Day of Sampling | Detection of S. aureus in Patient | Hla Detected | Monomeric Hla (ng/mL DTA) |
|---|---|---|---|---|
| 1 | 7 | no | yes | 3173 |
| 5 | 0 | no | yes | n.d. |
| 16 | 0 | yes | yes | 16 |
| 17 | 7 | yes | yes | 128 |
| 14 | yes | yes | 56 | |
| 21 | 0 | yes | yes | 384 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziesemer, S.; Kuhn, S.-O.; Hahnenkamp, A.; Gerber, M.; Lutjanov, E.; Gruendling, M.; Hildebrandt, J.-P. Staphylococcus aureus Alpha-Toxin in Deep Tracheal Aspirates—Preliminary Evidence for Its Presence in the Lungs of Sepsis Patients. Toxins 2022, 14, 450. https://doi.org/10.3390/toxins14070450
Ziesemer S, Kuhn S-O, Hahnenkamp A, Gerber M, Lutjanov E, Gruendling M, Hildebrandt J-P. Staphylococcus aureus Alpha-Toxin in Deep Tracheal Aspirates—Preliminary Evidence for Its Presence in the Lungs of Sepsis Patients. Toxins. 2022; 14(7):450. https://doi.org/10.3390/toxins14070450
Chicago/Turabian StyleZiesemer, Sabine, Sven-Olaf Kuhn, Anke Hahnenkamp, Manuela Gerber, Elvira Lutjanov, Matthias Gruendling, and Jan-Peter Hildebrandt. 2022. "Staphylococcus aureus Alpha-Toxin in Deep Tracheal Aspirates—Preliminary Evidence for Its Presence in the Lungs of Sepsis Patients" Toxins 14, no. 7: 450. https://doi.org/10.3390/toxins14070450
APA StyleZiesemer, S., Kuhn, S.-O., Hahnenkamp, A., Gerber, M., Lutjanov, E., Gruendling, M., & Hildebrandt, J.-P. (2022). Staphylococcus aureus Alpha-Toxin in Deep Tracheal Aspirates—Preliminary Evidence for Its Presence in the Lungs of Sepsis Patients. Toxins, 14(7), 450. https://doi.org/10.3390/toxins14070450