An Ecotoxicological Evaluation of Four Fungal Metabolites with Potential Application as Biocides for the Conservation of Cultural Heritage

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Instruments and Chemicals
4.2. Production, Isolation, and Identification of Selected Compounds
4.3. Stability Studies on the Selected Compounds
4.3.1. Qualitative Analysis
4.3.2. Quantitative Analysis
4.4. Ecotoxicity Analysis
4.4.1. Algal Growth Inhibition
4.4.2. Luminescence Bacteria Inhibition
4.4.3. Crustacean Immobility
4.4.4. Nematocidal Activity
4.4.5. Root Growth Inhibition
4.5. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Romani, M.; Carrion, C.; Fernandez, F.; Intertaglia, L.; Pecqueur, D.; Lebaron, P.; Lami, R. High bacterial diversity in pioneer biofilms colonizing ceramic roof tiles. Int. Biodeterm. Biodegrad. 2019, 144, 104745. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, P. What about biofilms on the surface of stone monuments. Open Conf. Proc. J. 2016, 7, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, M.L.; Miller, A.Z.; Macedo, M.F. Biological colonization and biodeterioration of architectural ceramic materials: An overview. J. Cult. Herit. 2015, 16, 759–777. [Google Scholar] [CrossRef] [Green Version]
- Caneva, G.; Nugari, M.P.; Salvadori, O. Plant Biology for Cultural Heritage: Biodeterioration and Conservation; Getty Publications: Los Angeles, CA, USA, 2008; ISBN 978-0-89236-939-3. [Google Scholar]
- Lo Schiavo, S.; De Leo, F.; Urzì, C. Present and future perspectives for biocides and antifouling products for stone-built cultural heritage: Ionic liquids as a challenging alternative. Appl. Sci. 2020, 10, 6568. [Google Scholar] [CrossRef]
- Pinna, D. Coping with Biological Growth on Stone Heritage Objects: Methods, Products, Applications, and Perspectives; Apple Academic Press: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- AA.VV. Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 Concerning the Making Available on the Market and Use of Biocidal Products. 2012. Available online: http://data.europa.eu/eli/reg/2012/528/2019-11-20 (accessed on 18 September 2020).
- Garcia-Hidalgo, E.; Sottas, V.; Goetz, N.; Von Hauri, U.; Bogdal, C.; Hungerbühler, K. Occurrence and concentrations of isothiazolinones in detergents and cosmetics in Switzerland. Contact Derm. 2016, 76, 96–106. [Google Scholar] [CrossRef]
- Wieck, S.; Olsson, O.; Kümmerer, K. Not only biocidal products: Washing and cleaning agents and personal care products can act as further sources of biocidal active substances in wastewater. Environ. Int. 2018, 115, 247–256. [Google Scholar] [CrossRef]
- Wieck, S.; Olsson, O.; Kümmerer, K. Possible underestimations of risks for the environment due to unregulated emissions of biocides from households to wastewater. Environ. Int. 2016, 94, 695–705. [Google Scholar] [CrossRef]
- Bollmann, U.E.; Fernández-Calviño, D.; Brandt, K.K.; Storgaard, M.S.; Sanderson, H.; Bester, K. Biocide runoff from building facades: Degradation kinetics in soil. Environ. Sci. Technol. 2017, 51, 3694–3702. [Google Scholar] [CrossRef]
- Chand, R.; Tulucan, T.; Aburlacitei, M. Investigation of biocide biodegradation in wastewater under laboratory set-up in anaerobic, aerobic and aerobic with substrate conditions. J. Civ. Environ. Eng. 2018, 8, 2. [Google Scholar] [CrossRef]
- Palla, F.; Bruno, M.; Mercurio, F.; Tantillo, A.; Rotolo, V. Essential oils as natural biocides in conservation of cultural heritage. Molecules 2020, 25, 730. [Google Scholar] [CrossRef] [Green Version]
- Bruno, L.; Rugnini, L.; Spizzichino, V.; Caneve, L.; Canini, A.; Ellwood, N.T.W. Biodeterioration of Roman hypogea: The case study of the Catacombs of SS. Marcellino and Pietro (Rome, Italy). Ann. Microbiol. 2019, 69, 1023–1032. [Google Scholar] [CrossRef]
- Arreche, R.; Vázquez, P. Green biocides to control biodeterioration in materials science and the example of preserving World Heritage Monuments. Curr. Opin. Green Sustain. Chem. 2020, 25, 100359. [Google Scholar] [CrossRef]
- Kakakhel, M.A.; Wu, F.; Gu, J.D.; Feng, H.; Shah, K.; Wang, W. Controlling biodeterioration of cultural heritage objects with biocides: A review. Int. Biodeterer. Biodegrad. 2019, 143, 104721. [Google Scholar] [CrossRef]
- Fidanza, M.R.; Caneva, G. Natural biocides for the conservation of stone cultural heritage: A review. J. Cult. Herit. 2019, 38, 271–286. [Google Scholar] [CrossRef]
- Silva, M.; Rosado, T.; da Silva, Z.L.; Nóbrega, Y.; Silveira, D.; Candeias, A.; Caldeira, A.T. Green bioactive compounds: Mitigation strategies for cultural heritage. Conserv. Sci. Cult. Herit. 2019, 19, 133–142. [Google Scholar]
- Petraretti, M.; Duffy, K.J.; Del Mondo, A.; Pollio, A.; De Natale, A. Community composition and ex situ cultivation of fungi associated with UNESCO heritage monuments in the bay of Naples. Appl. Sci. 2021, 11, 4327. [Google Scholar] [CrossRef]
- Masi, M.; Petraretti, M.; De Natale, A.; Pollio, A.; Evidente, A. Fungal metabolites with antagonistic activity against fungi of lithic substrata. Biomolecules 2021, 11, 295. [Google Scholar] [CrossRef]
- Manual of Decisions for BPD-98/EC. 2005. Manual of Decisions for Implementations of Directive 98/8/EC Concerning the Placing on the Market of Biocidal Products. Last Modified 12.12.2005. Available online: http://ec.europa.eu/environment/biocides/pdf/mod.pdf (accessed on 13 June 2022).
- Evidente, A.; Sparapano, L.; Fierro, O.; Bruno, G.; Giordano, F.; Motta, A. Sphaeropsidone and episphaeropsidone, phytotoxic dimedone methylethers produced by Sphaeropsis sapinea f. sp. cupressi grown in liquid culture. Phytochemistry 1998, 48, 1139–1143. [Google Scholar] [CrossRef]
- Evidente, A.; Randazzo, G.; Ballio, A. Structure determination of seiridin and isoseiridin, phytotoxic butenolides from culture filtrate of Seiridium cardinale. J. Nat. Prod. 1986, 49, 593–601. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Scanu, B.; Evidente, A.; Cimmino, A. Bioactive metabolites from pathogenic and endophytic fungi of forest trees. Curr. Med. Chem. 2018, 25, 208–252. [Google Scholar] [CrossRef]
- Evidente, A.; Randazzo, G.; Iacobellis, N.S.; Bottalico, A. Structure of cavoxin, a new phytotoxin from Phoma cava and cavoxone, its related chroman-4-one. J. Nat. Prod. 1985, 48, 916–923. [Google Scholar] [CrossRef]
- Andolfi, A.; Maddau, L.; Basso, S.; Linaldeddu, B.T.; Cimmino, A.; Scanu, B.; Evidente, A. Diplopimarane, a 20-nor-ent-pimarane produced by the oak pathogen Diplodia quercivora. J. Nat. Prod. 2014, 77, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Maddau, L.; Scanu, B.; Andolfi, A.; Masi, M.; Motta, A.; Tuzi, A. Sphaeropsidones, phytotoxic dimedone methyl ethers produced by Diplodia cupressi: A structure−activity relationship study. J. Nat. Prod. 2011, 74, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Cala, A.; Masi, M.; Cimmino, A.; Molinillo, J.M.; Macias, F.A.; Evidente, A. (+)-epi-Epoformin, a phytotoxic fungal cyclohexenepoxide: Structure activity relationships. Molecules 2018, 23, 1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barilli, E.; González-Bernal, M.J.; Cimmino, A.; Agudo-Jurado, F.J.; Masi, M.; Rubiales, D.; Evidente, A. Impact of fungal and plant metabolites application on early development stages of pea powdery mildew. Pest Manag. Sci. 2019, 75, 2464–2473. [Google Scholar] [CrossRef] [PubMed]
- Schrader, K.K.; Andolfi, A.; Cantrell, C.L.; Cimmino, A.; Duke, S.O.; Osbrink, W.; Wedge, D.E.; Evidente, A. A survey of phytotoxic microbial and plant metabolites as potential natural products for pest management. Chem. Biodivers. 2010, 7, 2261–2280. [Google Scholar] [CrossRef]
- Barilli, E.; Cimmino, A.; Masi, M.; Evidente, M.; Rubiales, D.; Evidente, A. Inhibition of spore germination and appressorium formation of rust species by plant and fungal metabolites. Nat. Prod. Comm. 2016, 11, 1934578X1601100940. [Google Scholar] [CrossRef] [Green Version]
- Barilli, E.; Cimmino, A.; Masi, M.; Evidente, M.; Rubiales, D.; Evidente, A. Inhibition of early development stages of rust fungi by the two fungal metabolites cyclopaldic acid and epi-epoformin. Pest Manag. Sci. 2017, 73, 1161–1168. [Google Scholar] [CrossRef]
- Sparapano, L.; Evidente, A. Studies on structure-activity relationship of seiridins, phytotoxins produced by three species of Seiridium. Nat. Toxins 1995, 3, 166–173. [Google Scholar] [CrossRef]
- Sangermano, F.; Masi, M.; Kumar, A.; Peravali, R.; Tuzi, A.; Cimmino, A.; Vallone, D.; Giamundo, D.; Conte, I.; Evidente, A.; et al. In vitro and in vivo toxicity evaluation of natural products with potential applications as biopesticides. Toxins 2021, 13, 805. [Google Scholar] [CrossRef]
- Bellas, J. Comparative toxicity of alternative antifouling biocides on embryos and larvae of marine invertebrates. Sci. Total Environ. 2006, 367, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, M. Globally harmonized system of classification and labelling of chemicals (GHS) and its implementation in Japan. Nihon Eiseigaku zasshi. Jpn. J. Hyg. 2010, 65, 5–13. [Google Scholar] [CrossRef] [PubMed]
- ISO 8692; Water Quality–Fresh Water Algal Growth Inhibition Test with Unicellular Green Algae. ISO: Geneva, Switzerland, 2012.
- Masi, M.; Meyer, S.; Clement, S.; Cimmino, A.; Evidente, A. Effect of cultural conditions on the production of radicinin, a specific fungal phytotoxin for buffelgrass (Cenchrus ciliaris) biocontrol, by different Cochlioboulus australiensis strains. Nat. Prod. Res. 2021, 35, 99–107. [Google Scholar] [CrossRef]
- ISO 11348-3:2007; Water Quality–Determination of the Inhibitory Effect of Water Samples on the Light Emission of Aliivibrio fischeri (Luminescent Bacteria Test)–Part 3: Method Using Freeze-Dried Bacteria. ISO: Geneva, Switzerland, 2007.
- ISO 6341:2012; Water Quality-Determination of the Inhibition of the Mobility of Daphnia Magna Straus (Cladocera, Crustacea)–Acute Toxicity Test. International Organisation for Standardisation: Geneva, Switzerland, 2012. Available online: https://www.iso.org/standard/54614.html (accessed on 21 April 2021).
- ISO 11269-1:2012; Soil Quality—Determination of the Effects of Pollutants on Soil Flora—Part 1: Method for the Measurement of Inhibition of Root Growth. ISO: Geneva, Switzerland, 2012.
- Libralato, G.; Costa Devoti, A.; Zanella, M.; Sabbioni, E.; Mičetić, I.; Manodori, L. Phytotoxicity of ionic, micro- and nano-sized iron in three plant species Ecotoxicol. Environ. Saf. 2016, 123, 81–88. [Google Scholar] [CrossRef] [PubMed]
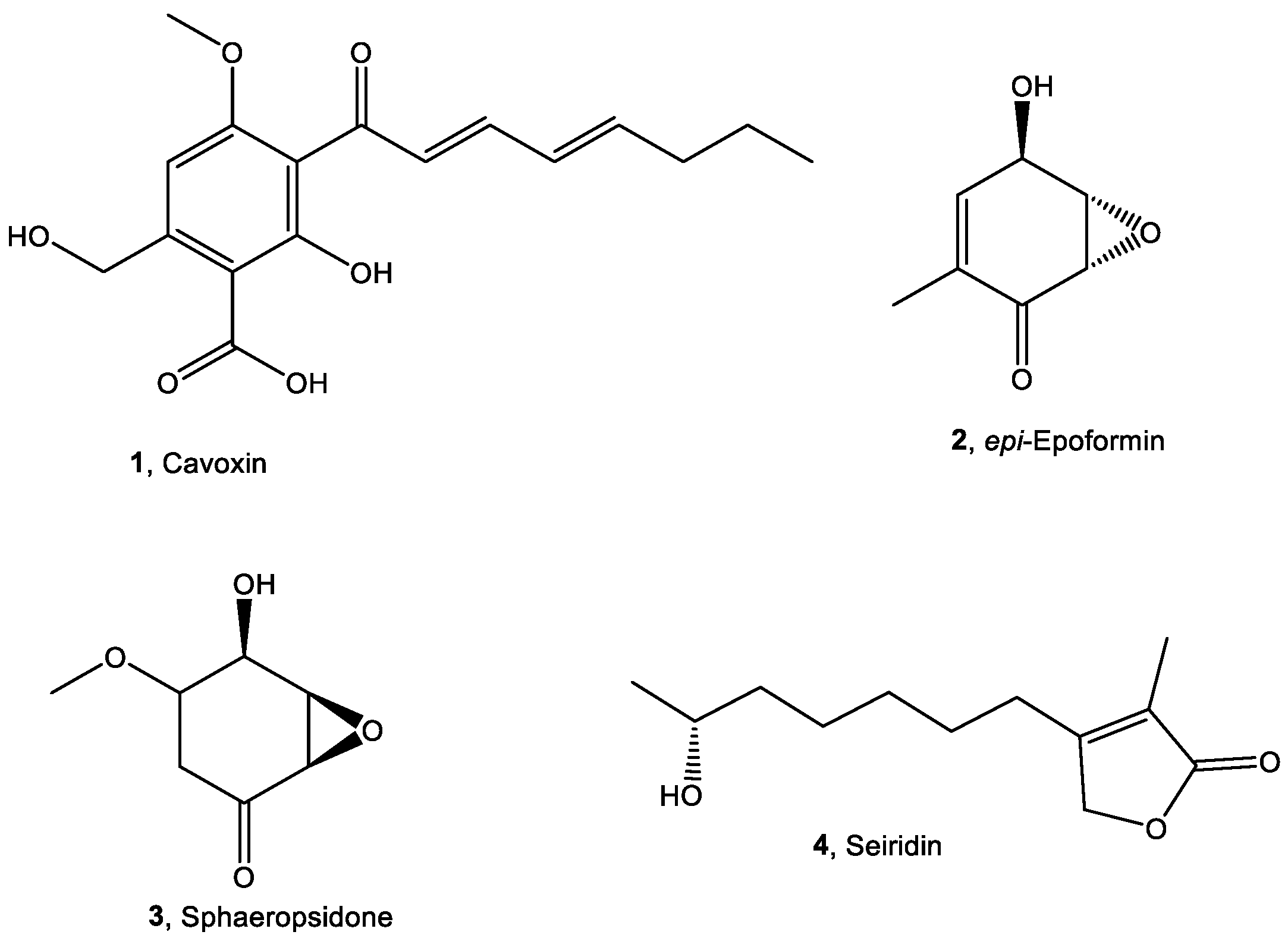
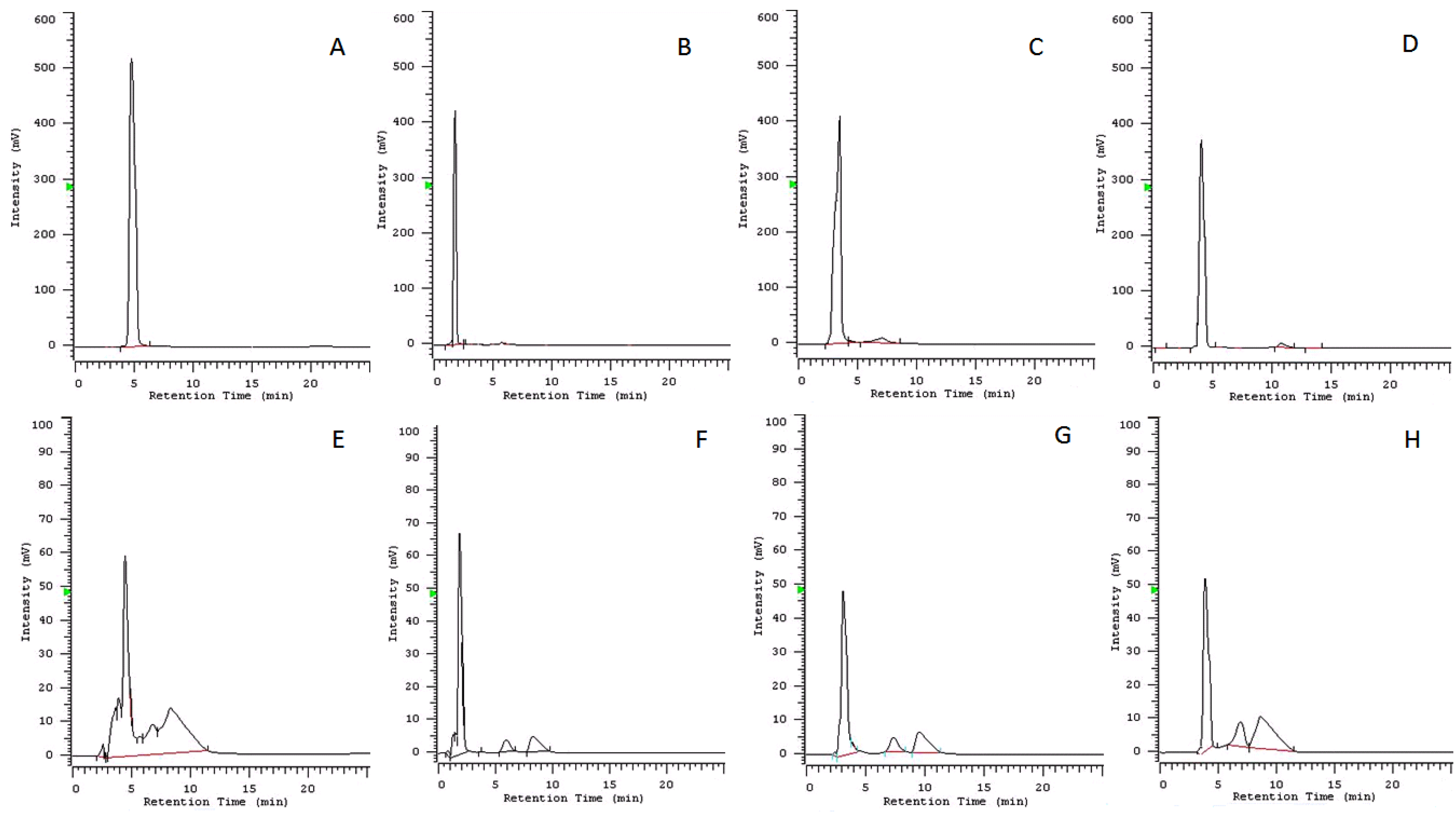

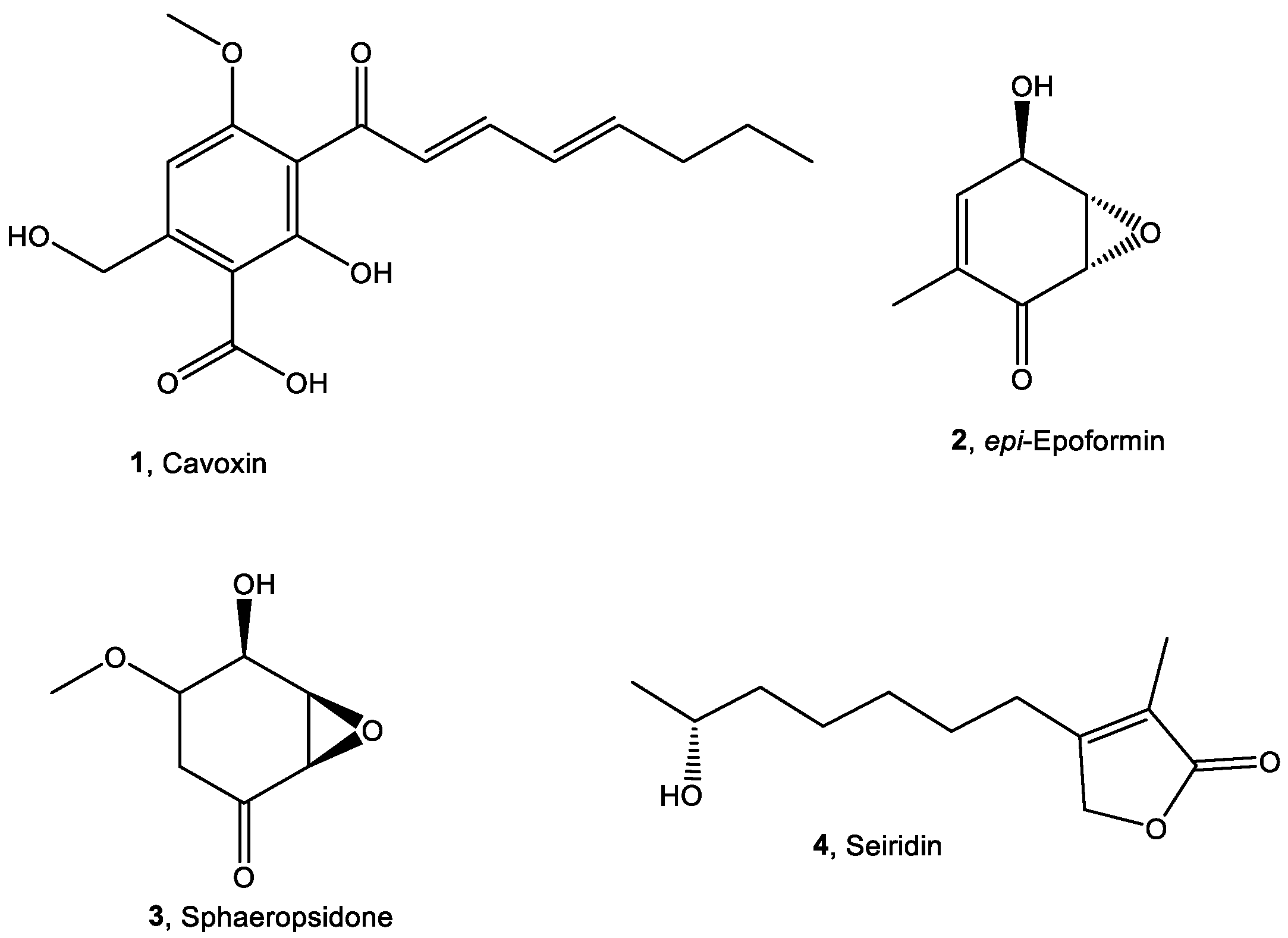{kind=link}
{kind=link}
| Compound | Rt (min) | R2 | Detection Limit (μg) | Compound Detected (μg) in 10 μL | % of Compound Present in the Culture Medium after 72 h |
|---|---|---|---|---|---|
| cavoxin (1) | 4.790 | 0.9998 | 0.0001 | n.d. b | 0 |
| epi-epoformin (2) | 1.830 | 0.9997 | 0.0001 | 0.00095 ± 0.0001 | 95 |
| sphaeropsidone (3) | 3.200 | 0.9998 | 0.0001 | 0.00092 ± 0.0002 | 92 |
| seiridin (4) | 4.080 | 0.9996 | 0.0003 | 0.00090 ± 0.0003 | 90 |
| Organism | Compound | EC5 | EC20 | EC50 |
|---|---|---|---|---|
| cavoxin (1) | 15.97 (10.11–25.13) | 20.93 (13.25–32.96) | 35.98 (22.77–56.67) | |
| Raphidocelis subcapitata | epi-epoformin (2) | 1.91 (1.28–2.99) | 2.58 (1.70–4.10) | 4.67 (2.98–7.70) |
| sphaeropsidone (3) | 1.78 (1.32–2.45) | 3.43 (2.49–4.82) | 12.78 (8.95–18.64) | |
| seiridin (4) | n.d. | n.d. | n.d. | |
| cavoxin (1) | 1.42 (0.93–2.24) | 2.59 (1.66–4.16) | 8.57 (5.24–14.42) | |
| Aliivibrio fischeri | epi-epoformin (2) | 0.59 (0.33–1.15) | 1.20 (0.64–2.48) | 5.12 (2.50–11.58) |
| sphaeropsidone (3) | 4.12 (3.29–5.15) | 10.49 (8.38–13.14) | 68.14 (54.43–85.29) | |
| seiridin (4) | 5.69 (2.95–11.00) | 10.01 (5.18–19.35) | 30.96 (16.02–59.84) | |
| cavoxin (1) | 2.64 (0.22–1.37) | 4.36 (0.38–2.20) | 1.91 (1.11–5.66) | |
| epi-epoformin (2) | 1.89 (1.13–3.36) | 2.68 (1.57–4.84) | 5.36 (3.03–10.04) | |
| Daphnia magna | sphaeropsidone (3) | 4.26 (1.20–5.06) | 6.49 (1.83–12.96) | 15.07 (4.26–16.34) |
| seiridin (4) | n.d. | n.d. | n.d. | |
| cavoxin (1) | 1.98 (1.08–3.91) | 3.34 (1.77–6.80) | 9.44 (4.73–20.52) | |
| epi-epoformin (2) | 1.10 (0.63–2.05) | 1.56 (0.88–2.97) | 3.12 (1.70–6.19) | |
| Caenorhabditis elegans | sphaeropsidone (3) | 1.12 (0.58–2.54) | 2.05 (1.00–5.02) | 6.88 (2.97–19.69) |
| seiridin (4) | n.d. | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petraretti, M.; Siciliano, A.; Carraturo, F.; Cimmino, A.; De Natale, A.; Guida, M.; Pollio, A.; Evidente, A.; Masi, M. An Ecotoxicological Evaluation of Four Fungal Metabolites with Potential Application as Biocides for the Conservation of Cultural Heritage. Toxins 2022, 14, 407. https://doi.org/10.3390/toxins14060407
Petraretti M, Siciliano A, Carraturo F, Cimmino A, De Natale A, Guida M, Pollio A, Evidente A, Masi M. An Ecotoxicological Evaluation of Four Fungal Metabolites with Potential Application as Biocides for the Conservation of Cultural Heritage. Toxins. 2022; 14(6):407. https://doi.org/10.3390/toxins14060407
Chicago/Turabian StylePetraretti, Mariagioia, Antonietta Siciliano, Federica Carraturo, Alessio Cimmino, Antonino De Natale, Marco Guida, Antonino Pollio, Antonio Evidente, and Marco Masi. 2022. "An Ecotoxicological Evaluation of Four Fungal Metabolites with Potential Application as Biocides for the Conservation of Cultural Heritage" Toxins 14, no. 6: 407. https://doi.org/10.3390/toxins14060407
APA StylePetraretti, M., Siciliano, A., Carraturo, F., Cimmino, A., De Natale, A., Guida, M., Pollio, A., Evidente, A., & Masi, M. (2022). An Ecotoxicological Evaluation of Four Fungal Metabolites with Potential Application as Biocides for the Conservation of Cultural Heritage. Toxins, 14(6), 407. https://doi.org/10.3390/toxins14060407