Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity

 , , ,
, , ,
Abstract

1. Introduction
2. Results
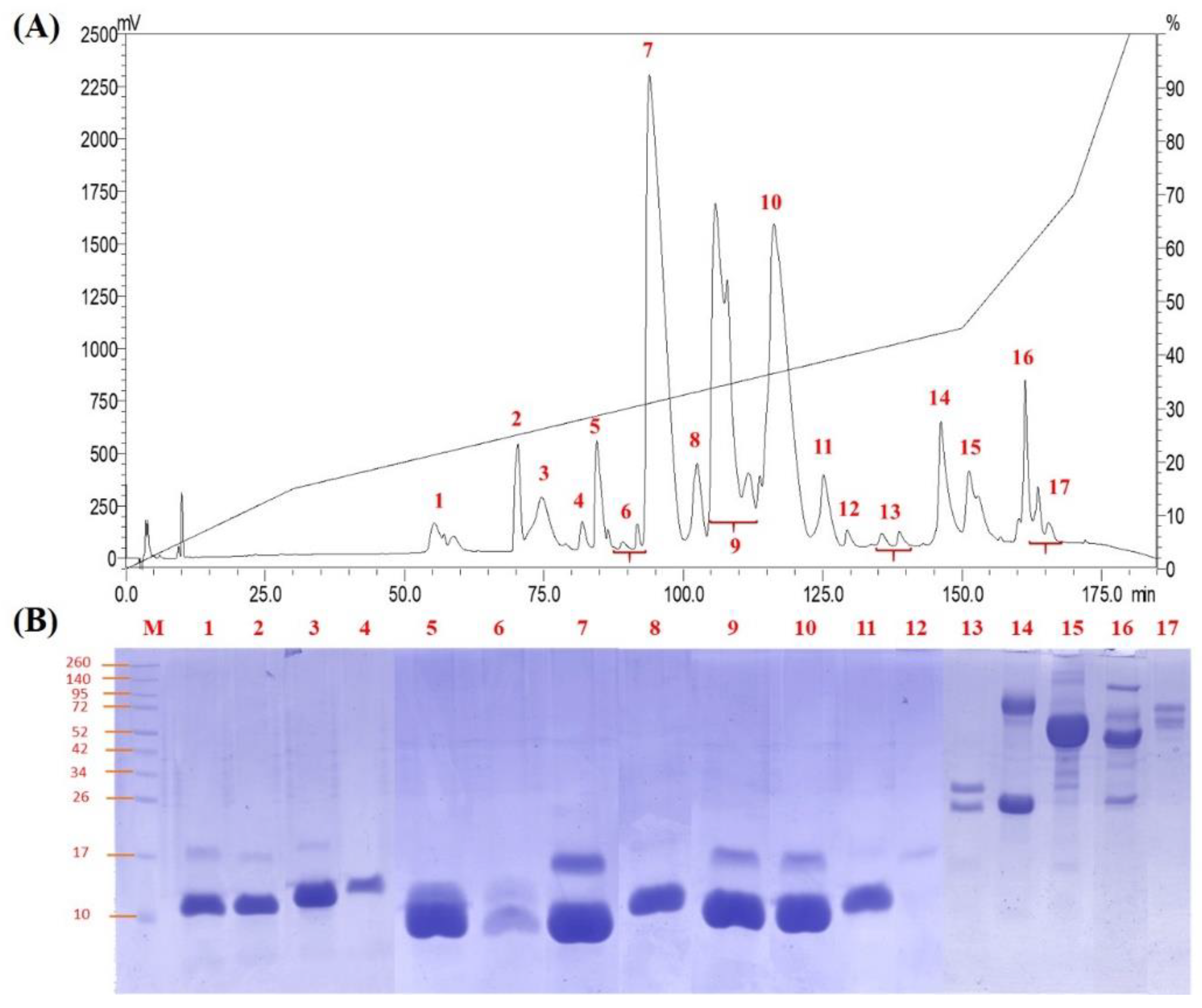
2.1. Decomplexation of N. nivea Venom by Reverse-Phase HPLC
2.2. Naja nivea Venom Proteome
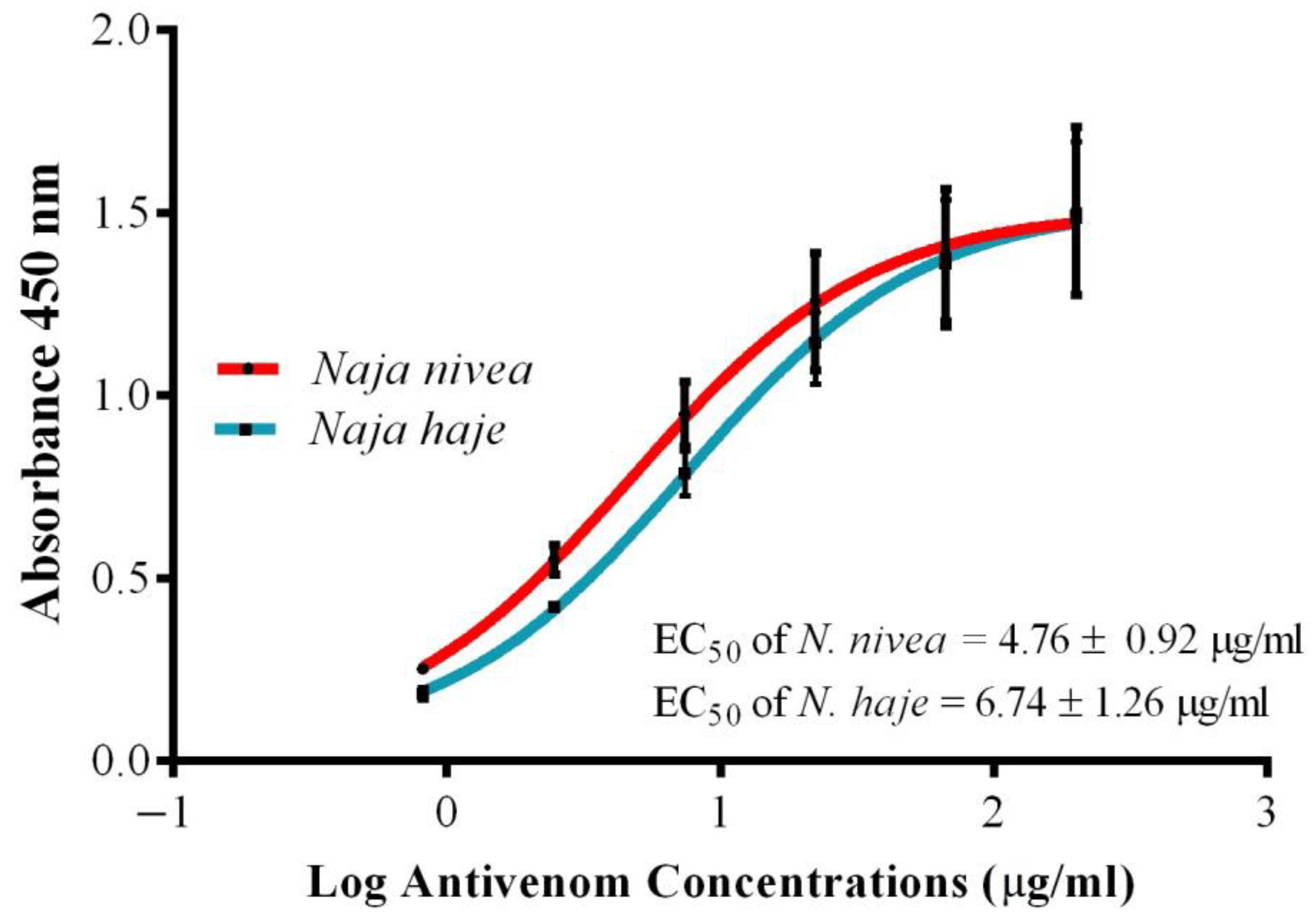
2.3. Immunological Binding Activity of VAPAV toward N. nivea Venom
2.4. Immunorecognition of N. nivea Venom Fractions by VAPAV
2.5. Venom Lethality and Neutralization by Antivenom
2.6. Experimental Envenoming and Rescue with Antivenom
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Materials
5.2. Venom and Antivenom
5.3. Animal Supply
5.4. Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC)
5.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
5.6. In-Solution Tryptic Digestion and Liquid Chromatography-Tandem Mass Spectrometry
5.7. Estimation of Protein Relative Abundance
5.8. Immunoreactivity of Antivenom
5.9. Antivenomics: Immunorecogniztion of N. nivea Venom Fractions by Antivenom
5.10. Venom Lethality and Neutralization by Antivenom
5.11. Experimental Envenoming and Rescue Experiment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Guidelines for the Management of Snakebites, 2nd ed.; WHO Regional Office for Southeast Asia: Dhaka, Bangladesh, 2016. [Google Scholar]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P.; Massougbodji, A.; Habib, A.G. The WHO strategy for prevention and control of snakebite envenoming: A sub-Saharan Africa plan. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e20190083. [Google Scholar] [CrossRef] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Halilu, S.; Iliyasu, G.; Hamza, M.; Chippaux, J.-P.; Kuznik, A.; Habib, A.G. Snakebite burden in Sub-Saharan Africa: Estimates from 41 countries. Toxicon Off. J. Int. Soc. Toxinology 2019, 159, 1–4. [Google Scholar] [CrossRef]
- Benjamin, J.M.; Abo, B.N.; Brandehoff, N. Review Article: Snake Envenomation in Africa. Curr. Trop. Med. Rep. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Wallach, V.; Wüster, W.; Broadley, G.D. In Praise Of Subgenera: Taxonomic Status Of Cobras Of The Genus Naja Laurenti (Serpentes: Elapidae). Zootaxa 2009, 2236, 26–36. [Google Scholar] [CrossRef]
- Broadley, D.G.; Wüster, W. A review of the southern African ‘non-spitting’ cobras (Serpentes: Elapidae: Naja). Afr. J. Herpetol. 2004, 53, 101–122. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Prevention and Clinical Management of Snakebite in Africa; 9290231688; World Health Organization, Regional Office for Africa: Brazzaville, Congo, 2010. [Google Scholar]
- Christensen, P.A. Problems of antivenene standardization revealed by the flocculation reaction. Bull. World Health Organ 1953, 9, 353–370. [Google Scholar] [PubMed]
- Hokama, Y.; Iwanaga, S.; Tatsuki, T.; Suzuki, T. Snake venom proteinase inhibitors. III. Isolation of five polypeptide inhibitors from the venoms of Hemachatus haemachatus (Ringhal’s corbra) and Naja nivea (Cape cobra) and the complete amino acid sequences of two of them. J. Biochem. 1976, 79, 559–578. [Google Scholar] [CrossRef] [PubMed]
- Joubert, F.J.; Taljaard, N. Snake venoms. The amino acid sequences of two Melanoleuca-type toxins. Hoppe. Seylers Z. Physiol. Chem. 1980, 361, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Botes, D.P.; Viljoen, C.C. The amino acid sequence of three non-curarimimetic toxins from Naja nivea venom. Biochim. Et Biophys. Acta (BBA)—Protein Struct. 1976, 446, 1–9. [Google Scholar] [CrossRef]
- Blaylock, R.S.; Lichtman, A.R.; Potgieter, P.D. Clinical manifestations of Cape cobra (Naja nivea) bites. A report of 2 cases. South Afr. Med. J. Suid-Afrik. Tydskr. Vir Geneeskd. 1985, 68, 342–344. [Google Scholar]
- Potet, J.; Smith, J.; McIver, L. Reviewing evidence of the clinical effectiveness of commercially available antivenoms in sub-Saharan Africa identifies the need for a multi-centre, multi-antivenom clinical trial. PLoS Negl. Trop. Dis. 2019, 13, e0007551. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, G.; Casewell, N.R.; Pla, D.; Quesada-Bernat, S.; Logan, R.A.E.; Bolton, F.M.S.; Wagstaff, S.C.; Gutiérrez, J.M.; Calvete, J.J.; Harrison, R.A. Defining the pathogenic threat of envenoming by South African shield-nosed and coral snakes (genus Aspidelaps), and revealing the likely efficacy of available antivenom. J. Proteom. 2019, 198, 186–198. [Google Scholar] [CrossRef]
- Ramos-Cerrillo, B.; de Roodt, A.R.; Chippaux, J.P.; Olguín, L.; Casasola, A.; Guzmán, G.; Paniagua-Solís, J.; Alagón, A.; Stock, R.P. Characterization of a new polyvalent antivenom (Antivipmyn Africa) against African vipers and elapids. Toxicon Off. J. Int. Soc. Toxinology 2008, 52, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Seddik, S.S.; Wanas, S.; Helmy, M.H.; Hashem, M. Cross neutralization of dangerous snake venoms from Africa and the Middle East using the VACSERA polyvalent antivenom. Egyptian Organization for Biological Products & Vaccines. J. Nat. Toxins 2002, 11, 329–335. [Google Scholar]
- Wong, K.Y.; Tan, K.Y.; Tan, N.H.; Tan, C.H. A Neurotoxic Snake Venom without Phospholipase A2: Proteomics and Cross-Neutralization of the Venom from Senegalese Cobra, Naja senegalensis (Subgenus: Uraeus). Toxins 2021, 13, 60. [Google Scholar] [CrossRef]
- Botes, D.P. Snake venom toxins. The amino acid sequences of toxins alpha and beta from Naja nivea venom and the disulfide bonds of toxin alpha. J. Biol. Chem. 1971, 246, 7383–7391. [Google Scholar] [CrossRef]
- Palasuberniam, P.; Chan, Y.W.; Tan, K.Y.; Tan, C.H. Snake Venom Proteomics of Samar Cobra (Naja samarensis) from the Southern Philippines: Short Alpha-Neurotoxins as the Dominant Lethal Component Weakly Cross-Neutralized by the Philippine Cobra Antivenom. Front. Pharm. 2021, 12, 727756. [Google Scholar] [CrossRef]
- Wong, K.Y.; Tan, C.H.; Tan, N.H. Venom and Purified Toxins of the Spectacled Cobra (Naja naja) from Pakistan: Insights into Toxicity and Antivenom Neutralization. Am. J. Trop. Med. Hyg. 2016, 94, 1392–1399. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Neutralization of the Principal Toxins from the Venoms of Thai Naja kaouthia and Malaysian Hydrophis schistosus: Insights into Toxin-Specific Neutralization by Two Different Antivenoms. Toxins 2016, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Loots, J.M.; Meij, H.S.; Meyer, B.J. Effects of Naja nivea venom on nerve, cardiac and skeletal muscle activity of the frog. Br. J. Pharmacol. 1973, 47, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Wong, K.Y.; Chong, H.P.; Tan, N.H.; Tan, K.Y. Proteomic insights into short neurotoxin-driven, highly neurotoxic venom of Philippine cobra (Naja philippinensis) and toxicity correlation of cobra envenomation in Asia. J. Proteom. 2019, 206, 103418. [Google Scholar] [CrossRef]
- Wong, K.Y.; Tan, K.Y.; Tan, N.H.; Gnanathasan, C.A.; Tan, C.H. Elucidating the Venom Diversity in Sri Lankan Spectacled Cobra (Naja naja) through De Novo Venom Gland Transcriptomics, Venom Proteomics and Toxicity Neutralization. Toxins 2021, 13, 558. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Wong, K.Y.; Tan, N.H.; Chong, H.P. Equatorial Spitting Cobra (Naja sumatrana) from Malaysia (Negeri Sembilan and Penang), Southern Thailand, and Sumatra: Comparative Venom Proteomics, Immunoreactivity and Cross-Neutralization by Antivenom. Toxins 2022, 14, 522. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Sim, S.M.; Fung, S.Y.; Tan, N.H. Geographical venom variations of the Southeast Asian monocled cobra (Naja kaouthia): Venom-induced neuromuscular depression and antivenom neutralization. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2016, 185, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, L.P.; Laustsen, A.H.; Lomonte, B.; Gutierrez, J.M. Exploring the venom of the forest cobra snake: Toxicovenomics and antivenom profiling of Naja melanoleuca. J. Proteom. 2017, 150, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; Leon, G.; Warrell, D.A.; Theakston, R.D.; et al. Snake venomics of African spitting cobras: Toxin composition and assessment of congeneric cross-reactivity of the pan-African EchiTAb-Plus-ICP antivenom by antivenomics and neutralization approaches. J. Proteome. Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Wong, K.Y.; Tan, N.H.; Tan, C.H. Quantitative proteomics of Naja annulifera (sub-Saharan snouted cobra) venom and neutralization activities of two antivenoms in Africa. Int. J. Biol. Macromol. 2020, 158, 605–616. [Google Scholar] [CrossRef]
- Marais, J. A Complete Guide to the Snakes of Southern Africa; Struik Nature: Cape Town, South Africa, 2004. [Google Scholar]
- Nirthanan, S.; Gopalakrishnakone, P.; Gwee, M.C.; Khoo, H.E.; Kini, R.M. Non-conventional toxins from Elapid venoms. Toxicon Off. J. Int. Soc. Toxinology 2003, 41, 397–407. [Google Scholar] [CrossRef]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Shulepko, M.A.; Paramonov, A.S.; Chugunov, A.O.; Janickova, H.; Dolejsi, E.; Dolezal, V.; Utkin, Y.N.; Tsetlin, V.I.; et al. Structural Insight into Specificity of Interactions between Nonconventional Three-finger Weak Toxin from Naja kaouthia (WTX) and Muscarinic Acetylcholine Receptors. J. Biol. Chem. 2015, 290, 23616–23630. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.K.; Fung, S.Y.; Tan, C.H.; Sim, S.M.; Tan, N.H. Immunological cross-reactivity and neutralization of the principal toxins of Naja sumatrana and related cobra venoms by a Thai polyvalent antivenom (Neuro Polyvalent Snake Antivenom). Acta Trop. 2015, 149, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Condrea, E.; Devries, A.; Mager, J. Hemolysis and splitting of human erythrocyte phospholipids by snake venoms. Biochim. Et Biophys. Acta 1964, 84, 60–73. [Google Scholar] [CrossRef]
- Gasanov, S.E.; Alsarraj, M.A.; Gasanov, N.E.; Rael, E.D. Cobra venom cytotoxin free of phospholipase A2 and its effect on model membranes and T leukemia cells. J. Membr. Biol. 1997, 155, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Dubovskii, P.V.; Lesovoy, D.M.; Dubinnyi, M.A.; Konshina, A.G.; Utkin, Y.N.; Efremov, R.G.; Arseniev, A.S. Interaction of three-finger toxins with phospholipid membranes: Comparison of S- and P-type cytotoxins. Biochem. J. 2005, 387, 807–815. [Google Scholar] [CrossRef]
- Chien, K.Y.; Chiang, C.M.; Hseu, Y.C.; Vyas, A.A.; Rule, G.S.; Wu, W. Two distinct types of cardiotoxin as revealed by the structure and activity relationship of their interaction with zwitterionic phospholipid dispersions. J. Biol. Chem. 1994, 269, 14473–14483. [Google Scholar] [CrossRef]
- Chong, H.P.; Tan, K.Y.; Tan, C.H. Cytotoxicity of snake venoms and cytotoxins from two Southeast Asian cobras (Naja sumatrana, Naja kaouthia): Exploration of anticancer potential, selectivity, and cell death mechanism. Front. Mol. Biosci. 2020, 7, 583587. [Google Scholar] [CrossRef]
- Epstein, D. The pharmacology of the venom of the cape cobra. Q. J. Exp. Physiol. 1930, 20, 7–19. [Google Scholar] [CrossRef]
- Sudulagunta, S.R.; Sodalagunta, M.B.; Khorram, H.; Sepehrar, M.; Noroozpour, Z. Case Report Cardiotoxicity and respiratory failure due to Cobra bite. Sch. J. Appl. Med. Sci. 2015, 3, 1830–1833. [Google Scholar]
- Senthilkumaran, S.; Meenakshisundaram, R.; Thirumalaikolundusubramanian, P.; Menezes, R.G. Cardiac toxicity following cobra envenomation. Clin. Toxicol. 2012, 50, 862, author reply 863. [Google Scholar] [CrossRef][Green Version]
- Silva de França, F.; Villas-Boas, I.M.; Cogliati, B.; Woodruff, T.M.; Reis, E.d.S.; Lambris, J.D.; Tambourgi, D.V. C5a-C5aR1 axis activation drives envenomation immunopathology by the snake Naja annulifera. Front. Immunol. 2021, 12, 652242. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) From Venomous Snakes: An Overview of the Functional Diversity in A Large and Underappreciated Superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Mackessy, S.P.; Dutta, S. Characterization of a Kunitz-type protease inhibitor peptide (Rusvikunin) purified from Daboia russelii russelii venom. Int. J. Biol. Macromol. 2014, 67, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Earl, S.T.H.; Richards, R.; Johnson, L.A.; Flight, S.; Anderson, S.; Liao, A.; de Jersey, J.; Masci, P.P.; Lavin, M.F. Identification and characterisation of Kunitz-type plasma kallikrein inhibitors unique to Oxyuranus sp. snake venoms. Biochimie 2012, 94, 365–373. [Google Scholar] [CrossRef]
- Wijeyewickrema, L.C.; Gardiner, E.E.; Gladigau, E.L.; Berndt, M.C.; Andrews, R.K. Nerve growth factor inhibits metalloproteinase-disintegrins and blocks ectodomain shedding of platelet glycoprotein VI. J. Biol. Chem. 2010, 285, 11793–11799. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon Off. J. Int. Soc. Toxinology 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Aird, S.D. Taxonomic distribution and quantitative analysis of free purine and pyrimidine nucleosides in snake venoms. Comp. Biochem. Physiology. Part B Biochem. Mol. Biol. 2005, 140, 109–126. [Google Scholar] [CrossRef]
- Vogel, C.-W.; Bredehorst, R.; Fritzinger, D.C.; Grunwald, T.; Ziegelmüller, P.; Kock, M.A. Structure and Function of Cobra Venom Factor, the Complement-Activating Protein in Cobra Venom. In Natural Toxins 2: Structure, Mechanism of Action, and Detection; Singh, B.R., Tu, A.T., Eds.; Springer: Boston, MA, USA, 1996; pp. 97–114. [Google Scholar]
- Vogel, C.-W.; Fritzinger, D.C. Cobra Venom Factor: The Unique Component of Cobra Venom That Activates the Complement System. In Snake Venoms; Inagaki, H., Vogel, C.-W., Mukherjee, A.K., Rahmy, T.R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 345–404. [Google Scholar]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; Rego, C.M.A.; Silva, M.D.S.; Santana, H.M.; Serrath, S.N.; Ikenohuchi, Y.J.; Farias, B.J.C.; Felipin, K.P. Reactive oxygen species-dependent-NLRP3 inflammasome activation in human neutrophils induced by l-amino acid oxidase derived from Calloselasma rhodostoma venom. Life Sci. 2022, 308, 120962. [Google Scholar] [CrossRef]
- Cousin, X.; Bon, C. Acetylcholinesterase from snake venom as a model for its nerve and muscle counterpart. J. Nat. Toxins 1999, 8, 285–294. [Google Scholar]
- Pung, Y.F.; Kumar, S.V.; Rajagopalan, N.; Fry, B.G.; Kumar, P.P.; Kini, R.M. Ohanin, a novel protein from king cobra venom: Its cDNA and genomic organization. Gene 2006, 371, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Wong, K.Y.; Tan, N.H.; Ng, T.S.; Tan, K.Y. Distinctive Distribution of Secretory Phospholipases A2 in the Venoms of Afro-Asian Cobras (Subgenus: Naja, Afronaja, Boulengerina and Uraeus). Toxins 2019, 11, 116. [Google Scholar] [CrossRef]
- Malih, I.; Rusmili, A.M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of Moroccan cobra Naja haje legionis venom using tandem mass spectrometry. J. Proteom. 2014, 96, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics, lethality and neutralization of Naja kaouthia (monocled cobra) venoms from three different geographical regions of Southeast Asia. J Proteom. 2015, 120, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef]
- Habib, A.G.; Musa, B.M.; Iliyasu, G.; Hamza, M.; Kuznik, A.; Chippaux, J.-P. Challenges and prospects of snake antivenom supply in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008374. [Google Scholar] [CrossRef]
- WHO. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins, 2nd ed.; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Ratanabanangkoon, K.; Tan, K.Y.; Pruksaphon, K.; Klinpayom, C.; Gutiérrez, J.M.; Quraishi, N.H.; Tan, C.H. A pan-specific antiserum produced by a novel immunization strategy shows a high spectrum of neutralization against neurotoxic snake venoms. Sci. Rep. 2020, 10, 11261. [Google Scholar] [CrossRef]
- Ratanabanangkoon, K.; Tan, K.Y.; Eursakun, S.; Tan, C.H.; Simsiriwong, P.; Pamornsakda, T.; Wiriyarat, W.; Klinpayom, C.; Tan, N.H. A Simple and Novel Strategy for the Production of a Pan-specific Antiserum against Elapid Snakes of Asia. PLoS Negl. Trop. Dis. 2016, 10, e0004565. [Google Scholar] [CrossRef]
- Howard-Jones, N. A CIOMS ethical code for animal experimentation. WHO Chron. 1985, 39, 51–56. [Google Scholar]
- Tan, C.H.; Wong, K.Y.; Tan, K.Y.; Tan, N.H. Venom proteome of the yellow-lipped sea krait, Laticauda colubrina from Bali: Insights into subvenomic diversity, venom antigenicity and cross-neutralization by antivenom. J. Proteom. 2017, 166, 48–58. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 2nd ed.; Cambridge University Press: New York, NY, USA, 1952; p. 627. [Google Scholar] [CrossRef]
- Morais, V.; Ifran, S.; Berasain, P.; Massaldi, H. Antivenoms: Potency or median effective dose, which to use? J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 191–193. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, N.H.; Tan, C.H. Venom proteomics and antivenom neutralization for the Chinese eastern Russell’s viper, Daboia siamensis from Guangxi and Taiwan. Sci. Rep. 2018, 8, 8545. [Google Scholar] [CrossRef] [PubMed]
- Oh, A.M.F.; Tan, C.H.; Ariaranee, G.C.; Quraishi, N.; Tan, N.H. Venomics of Bungarus caeruleus (Indian krait): Comparable venom profiles, variable immunoreactivities among specimens from Sri Lanka, India and Pakistan. J. Proteom. 2017, 164, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, T.; Wu, S.; Yang, C.; Bai, M.; Shu, K.; Li, K.; Zhang, G.; Jin, Z.; He, F. iProX: An integrated proteome resource. Nucleic Acids Res. 2019, 47, D1211–D1217. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Database Accession a | Species | MS Search Score b | R.A. (%) c |
|---|---|---|---|---|
| Fraction 1 | ||||
| Short neurotoxin 2 | P01422 | Naja annulifera | 38.46 | 1.58 |
| Fraction 2 | ||||
| Short neurotoxin 4 | P01421 | Naja annulifera | 123.04 | 1.88 |
| Long neurotoxin 1 | P01390 | Naja nivea | 89.87 | 0.09 |
| Weak toxin S4C11 | P01400 | Naja melanoleuca | 46.32 | 0.04 |
| Fraction 3 | ||||
| Long neurotoxin 1 | P01390 | Naja nivea | 97.38 | 2.85 |
| Short neurotoxin 4 | P01421 | Naja annulifera | 53.91 | 0.07 |
| Weak toxin CM-11 | P01401 | Naja haje haje | 44.62 | 0.17 |
| Fraction 4 | ||||
| Long neurotoxin 1 | P01390 | Naja nivea | 114.03 | 0.02 |
| Weak toxin CM-13b | P01399 | Naja annulifera | 97.34 | 0.42 |
| Short neurotoxin 4 | P01421 | Naja annulifera | 34.64 | 0.08 |
| Fraction 5 | ||||
| Kunitz-type serine protease inhibitor 2 | P00986 | Naja nivea | 141.56 | 1.16 |
| Long neurotoxin 1 | P01390 | Naja nivea | 114.03 | 0.34 |
| Weak toxin CM-10 | P25680 | Naja nivea | 57.78 | 0.91 |
| Fraction 6 | ||||
| Long neurotoxin 1 | P01390 | Naja nivea | 102.63 | 0.32 |
| Kunitz-type serine protease inhibitor 2 | P00986 | Naja nivea | 82.24 | 0.07 |
| Weak toxin CM-13b | P01399 | Naja annulifera | 34.6 | 0.04 |
| Fraction 7 | ||||
| Cytotoxin 1 | P01456 | Naja nivea | 210.7 | 25.63 |
| Cytotoxin 2 | P01462 | Naja annulifera | 65.61 | 0.74 |
| Long neurotoxin 1 | P01390 | Naja nivea | 58.24 | 0.16 |
| Fraction 8 | ||||
| Cytotoxin 1 | P01456 | Naja nivea | 102.98 | 0.67 |
| Cytotoxin 8 | P01460 | Naja annulifera | 74.54 | 1.35 |
| Cytotoxin 2 | P01463 | Naja nivea | 68.3 | 0.26 |
| Cytotoxin 3 | P01458 | Naja nivea | 51.41 | 0.40 |
| Fraction 9 | ||||
| Cytotoxin 2 | P01462 | Naja annulifera | 131.23 | 4.41 |
| Cytotoxin 7 | P01466 | Naja annulifera | 116.39 | 5.03 |
| Cytotoxin 8 | P01460 | Naja annulifera | 99.86 | 6.97 |
| Cytotoxin 10 | P01453 | Naja annulifera | 88.36 | 2.10 |
| Cytotoxin 3 | P01458 | Naja nivea | 66.72 | 0.25 |
| Cytotoxin 1 | P01456 | Naja nivea | 61.69 | 3.04 |
| Venom nerve growth factor 2 | Q5YF89 | Naja sputatrix | 34.62 | 0.09 |
| Fraction 10 | ||||
| Cytotoxin 2 | P01463 | Naja nivea | 161.32 | 1.91 |
| Cytotoxin 3 | P01458 | Naja nivea | 164.72 | 9.68 |
| Cytotoxin 7 | P01466 | Naja annulifera | 97.35 | 2.92 |
| Cytotoxin 10 | P01453 | Naja annulifera | 64.96 | 2.41 |
| Cytotoxin 8 | P01460 | Naja annulifera | 52.47 | 4.93 |
| Venom nerve growth factor 2 | CL429.Contig1_NnSL | Naja naja | 42.38 | 1.12 |
| Fraction 11 | ||||
| Cytotoxin homolog | P14541 | Naja kaouthia | 98.04 | 1.78 |
| Cytotoxin 2 | P01463 | Naja nivea | 72.38 | 0.43 |
| Cytotoxin 1 | P01456 | Naja nivea | 42.36 | 0.07 |
| Cytotoxin 3 | P01459 | Naja annulifera | 40.31 | 0.21 |
| Cytotoxin 1 | P01468 | Naja pallida | 34.49 | 0.16 |
| Venom nerve growth factor 2 | Q5YF89 | Naja sputatrix | 32.84 | 0.01 |
| Fraction 12 | ||||
| Thaicobrin | P82885 | Naja kaouthia | 89.06 | 0.04 |
| Cytotoxin 3 | P01458 | Naja nivea | 87.75 | 0.03 |
| Cytotoxin 2 | P01463 | Naja nivea | 77.44 | 0.05 |
| Cytotoxin 1 | P01456 | Naja nivea | 74.18 | 0.02 |
| Cytotoxin 1 | P01468 | Naja pallida | 61.79 | 0.03 |
| Cytotoxin 8 | P01460 | Naja annulifera | 61.72 | 0.12 |
| Cytotoxin 10 | P01453 | Naja annulifera | 58.41 | 0.02 |
| Venom nerve growth factor 2 | Q5YF89 | Naja sputatrix | 81.11 | <0.01 |
| Cytotoxin homolog | P14541 | Naja kaouthia | 61.23 | 0.01 |
| Cytotoxin 11 | P62394 | Naja haje haje | 44.22 | 0.01 |
| Fraction 13 | ||||
| Cysteine-rich venom protein natrin-2 | Q7ZZN8 | Naja atra | 88.14 | 0.44 |
| Cysteine-rich venom protein latisemin | Q8JI38 | Laticauda semifasciata | 41.66 | 0.16 |
| Venom nerve growth factor 2 | Q5YF89 | Naja sputatrix | 72.47 | 0.01 |
| Zinc metalloproteinase-disintegrin atragin | CL626.Contig4_NsM | Naja sumatrana | 35.12 | 0.08 |
| Fraction 14 | ||||
| Natrin-1 | CL85.Contig1_NnSL | Naja naja | 139.79 | 1.41 |
| Cysteine-rich venom protein natrin-1 | Q7T1K6 | Naja atra | 134.04 | 1.41 |
| Zinc metalloproteinase-disintegrin cobrin | CL2966.Contig3_NnSL | Naja naja | 67.86 | 0.85 |
| Scutellatease-1 | CL2215.Contig1_HsM | Enhydrina schistosa | 38.33 | 0.31 |
| Cysteine-rich secretory protein Pg-CRP | F2Q6F6 | Cerrophidion godmani | 34.78 | 0.04 |
| SVMP-Aca-4 | R4G2D3 | Acanthophis wellsi | 29.96 | 0.03 |
| Fraction 15 | ||||
| Natrin-1 | CL85.Contig1_NnSL | Naja naja | 72.73 | 0.10 |
| Microlepidotease-1 | B5KFV6 | Oxyuranus microlepidotus | 52.23 | 0.15 |
| Metalloproteinase (Type III) 1 | U3EPC7 | Micrurus fulvius | 41.16 | 0.15 |
| Zinc metalloproteinase-disintegrin-like atragin | CL444.Contig1_NsM | Naja sumatrana | 53.63 | 0.03 |
| Hemorrhagic metalloproteinase-disintegrin-like kaouthiagin | P82942 | Naja kaouthia | 49.13 | 0.21 |
| SVMP-Aca-4 | R4G2D3 | Acanthophis wellsi | 44.77 | 0.01 |
| Zinc metalloproteinase-disintegrin-like atrase-A | D5LMJ3 | Naja atra | 48.07 | 1.49 |
| Zinc metalloproteinase-disintegrin cobrin | CL2966.Contig3_NnSL | Naja naja | 47.8 | 1.52 |
| Zinc metalloproteinase-disintegrin cobrin | CL7366.Contig1_OhM | Ophiophagus hannah | 30.86 | 0.03 |
| Fraction 16 | ||||
| Phosphodiesterase 1 | Unigene5869_NsM | Naja sumatrana | 167.77 | 0.15 |
| Phosphodiesterase | A0A194ARD7 | Micrurus tener | 129.25 | 0.30 |
| Phosphodiesterase 1 | CL4383.Contig2_OhM | Ophiophagus hannah | 89.99 | 0.54 |
| Snake venom 5′-nucleotidase | CL3600.Contig1_NsM2 | Naja sumatrana | 119.17 | 0.38 |
| Snake venom 5′-nucleotidase | CL4180.Contig1_OhM | Ophiophagus hannah | 108.51 | 0.36 |
| Zinc metalloproteinase-disintegrin-like atragin | CL444.Contig1_NsM | Naja sumatrana | 78.8 | 0.62 |
| Zinc metalloproteinase-disintegrin-like atragin | D3TTC2 | Naja atra | 45.45 | 0.21 |
| Zinc metalloproteinase-disintegrin atragin | CL2051.Contig1_NkM | Naja kaouthia | 29.61 | 0.33 |
| Fraction 17 | ||||
| Cobra venom factor | Unigene370_NsM2 | Naja sumatrana | 268.75 | 0.03 |
| Cobra venom factor | CL4560.Contig1_NsM | Naja sumatrana | 267 | 0.04 |
| Cobra venom factor | Q91132 | Naja kaouthia | 136.91 | 0.13 |
| L-amino-acid oxidase | CL4047.Contig1_NsM2 | Naja sumatrana | 202.09 | 0.07 |
| L-amino-acid oxidase | A8QL58 | Naja atra | 187.13 | 0.06 |
| L-amino acid oxidase | A8QL51 | Bungarus multicinctus | 67.97 | 0.15 |
| L-amino-acid oxidase | CL2322.Contig1_HhSL | Hypnale hypnale | 36.77 | 0.20 |
| Acetylcholinesterase | CL4231.Contig1_NsM | Naja sumatrana | 114.16 | 0.05 |
| Acetylcholinesterase | Unigene16279_OhM | Ophiophagus hannah | 85.55 | 0.05 |
| Snake venom 5′-nucleotidase | CL3600.Contig1_NsM2 | Naja sumatrana | 77.5 | 0.01 |
| Snake venom metalloproteinase-disintegrin-like mocarhagin | Q10749 | Naja mossambica | 65.43 | 0.24 |
| SVMP-Hop-45 | R4G2Y9 | Hoplocephalus bungaroides | 36.28 | 0.38 |
| Zinc metalloproteinase-disintegrin cobrin | CL115.Contig9_NkT | Naja kaouthia | 30.06 | 0.01 |
| Zinc metalloproteinase-disintegrin atragin | CL626.Contig4_NsM2 | Naja sumatrana | 54.02 | 0.02 |
| Zinc metalloproteinase mocarhagin | Unigene25077_NnSL | Naja naja | 32.68 | 0.04 |
| Zinc metalloproteinase-disintegrin-like VLAIP-A | Q4VM08 | Macrovipera lebetina | 30.22 | 0.11 |
| Phosphodiesterase 1 | unigene5869_NsM | Naja sumatrana | 27.96 | <0.01 |
| Protein Family | Protein Name | Accession a | Species | R.A. (%) b |
|---|---|---|---|---|
| Three-finger toxins (3FTX) | 84.62 | |||
| Short neurotoxin (SNTX) | 3.61 | |||
| Short neurotoxin 2 | P01422 | Naja annulifera | 1.58 | |
| Short neurotoxin 4 | P01421 | Naja annulifera | 2.03 | |
| Long neurotoxin (LNTX) | 3.78 | |||
| Long neurotoxin 1 | P01390 | Naja nivea | 3.78 | |
| Weak neurotoxin (WTX) | 1.58 | |||
| Weak toxin CM-13b | P01399 | Naja annulifera | 0.45 | |
| Weak toxin S4C11 | P01400 | Naja melanoleuca | 0.04 | |
| Weak toxin CM-11 | P01401 | Naja haje haje | 0.17 | |
| Weak toxin CM-10 | P25680 | Naja nivea | 0.91 | |
| Cytotoxin/cardiotoxin (CTX) | 75.65 | |||
| Cytotoxin 10 | P01453 | Naja annulifera | 4.53 | |
| Cytotoxin 1 | P01456 | Naja nivea | 29.43 | |
| Cytotoxin 3 | P01458 | Naja nivea | 10.36 | |
| Cytotoxin 3 | P01459 | Naja annulifera | 0.21 | |
| Cytotoxin 8 | P01460 | Naja annulifera | 13.37 | |
| Cytotoxin 2 | P01462 | Naja annulifera | 5.15 | |
| Cytotoxin 2 | P01463 | Naja nivea | 2.65 | |
| Cytotoxin 7 | P01466 | Naja annulifera | 7.95 | |
| Cytotoxin 1 | P01468 | Naja pallida | 0.20 | |
| Cytotoxin homolog | P14541 | Naja kaouthia | 1.79 | |
| Cytotoxin 11 | P62394 | Naja haje haje | <0.01 | |
| Snake venom metalloproteinase (SVMP) | 6.79 | |||
| microlepidotease-1 | B5KFV6 | Oxyuranus microlepidotus | 0.15 | |
| Zinc metalloproteinase-disintegrin cobrin | CL115.Contig9_NkT | Naja kaouthia | 0.01 | |
| Zinc metalloproteinase-disintegrin atragin | CL2051.Contig1_NkM | Naja kaouthia | 0.33 | |
| Scutellatease-1 | CL2215.Contig1_HsM | Hydrophis schistosus | 0.30 | |
| Zinc metalloproteinase-disintegrin cobrin | CL2966.Contig3_NnSL | Naja naja | 2.37 | |
| Zinc metalloproteinase-disintegrin-like atragin | CL444.Contig1_NsM | Naja sumatrana | 0.64 | |
| Zinc metalloproteinase-disintegrin atragin | CL626.Contig4_NsM2 | Naja sumatrana | 0.10 | |
| Zinc metalloproteinase-disintegrin cobrin | CL7366.Contig1_OhM | Ophiophagus hannah | 0.03 | |
| Zinc metalloproteinase-disintegrin-like atragin | D3TTC2 | Naja atra | 0.21 | |
| Zinc metalloproteinase-disintegrin-like atrase-A | D5LMJ3 | Naja atra | 1.49 | |
| Hemorrhagic metalloproteinase-disintegrin-like kaouthiagin | P82942 | Naja kaouthia | 0.21 | |
| Snake venom metalloproteinase-disintegrin-like mocarhagin | Q10749 | Naja mossambica | 0.24 | |
| Zinc metalloproteinase-disintegrin-like VLAIP-A | Q4VM08 | Macrovipera lebetina | 0.11 | |
| SVMP-Aca-4 | R4G2D3 | Acanthophis wellsi | 0.04 | |
| SVMP-Hop-45 | R4G2Y9 | Hoplocephalus bungaroides | 0.38 | |
| Metalloproteinase (Type III) 1 | U3EPC7 | Micrurus fulvius | 0.15 | |
| Zinc metalloproteinase mocarhagin | unigene25077_NnSL | Naja naja | 0.04 | |
| Cysteine-rich secretory protein (CRISP) | 3.56 | |||
| Natrin-1 | CL85.Contig1_NnSL | Naja naja | 1.51 | |
| Cysteine-rich venom protein ophanin | Q7ZT98 | Ophiophagus hannah | 0.04 | |
| Cysteine-rich venom protein natrin-1 | Q7T1K6 | Naja atra | 1.41 | |
| Cysteine-rich venom protein natrin-2 | Q7ZZN8 | Naja atra | 0.44 | |
| Cysteine-rich venom protein latisemin | Q8JI38 | Laticauda semifasciata | 0.16 | |
| Kunitz-type serine protease inhibitor (KSPI) | 1.24 | |||
| Kunitz-type serine protease inhibitor 2 | P00986 | Naja nivea | 1.24 | |
| Nerve growth factor (NGF) | 1.22 | |||
| Venom nerve growth factor 2 | CL429.Contig1_NnSL | Naja naja | 1.12 | |
| Venom nerve growth factor 2 | Q5YF89 | Naja sputatrix | 0.10 | |
| Phosphodiesterase (PDE) | 1.00 | |||
| Phosphodiesterase | A0A194ARD7 | Micrurus tener | 0.30 | |
| Phosphodiesterase 1 | CL4383.Contig2_OhM | Ophiophagus hannah | 0.54 | |
| Phosphodiesterase 1 | unigene5869_NsM | Naja sumatrana | 0.15 | |
| 5′-nucleotidase (5′-NUC) | 0.75 | |||
| Snake venom 5′-nucleotidase | CL3600.Contig1_NsM2 | Naja sumatrana | 0.39 | |
| Snake venom 5′-nucleotidase | CL4180.Contig1_OhM | Ophiophagus hannah | 0.36 | |
| L-amino acid oxidase (LAAO) | 0.48 | |||
| L-amino-acid oxidase | CL4047.Contig1_NsM2 | Naja sumatrana | 0.07 | |
| L-amino-acid oxidase | A8QL58 | Naja atra | 0.06 | |
| L-amino acid oxidase | A8QL51 | Bungarus multicinctus | 0.15 | |
| L-amino-acid oxidase-like | A0A6J1W8Y3 | Notechis scutatus | 0.20 | |
| Cobra venom factor (CVF) | 0.20 | |||
| Cobra venom factor | unigene370_NsM2 | Naja sumatrana | 0.03 | |
| Cobra venom factor | CL4560.Contig1_NsM | Naja sumatrana | 0.04 | |
| Cobra venom factor | Q91132 | Naja kaouthia | 0.13 | |
| Acetylcholinesterase (AChE) | 0.10 | |||
| Acetylcholinesterase NS2 | CL4231.Contig1_NsM | Naja sumatrana | 0.05 | |
| Acetylcholinesterase | unigene16279_OhM | Ophiophagus hannah | 0.05 | |
| Vespryn (VES) | 0.04 | |||
| Thaicobrin | P82885 | Naja kaouthia | 0.04 | |
| Number of Protein Fractions a | Immunorecognition Capacity of VAPAV b (%) |
|---|---|
| 1 | 67.0 |
| 2, 3 | 70.2 |
| 4 | 24.8 |
| 5 | 22.3 |
| 6 | 22.9 |
| 7 | 25.5 |
| 8 | 24.4 |
| 9, 10, 11 | 31.3 |
| 12 | 58.8 |
| i.v. LD50 a (µg/g) | s.c. LD50 a (µg/g) | Antivenom | Antivenom Protein Concentration (mg/mL) | Challenge Dose | ED50 b (µL) | ER50 c (mg/mL) | Potency d (mg/mL) | Normalized Potency e (n-P) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1.11 (0.73–1.69) | 2.42 (2.10–2.79) | VAPAV | 130.68 ± 2.83 # | 2.5 | 74.61 | 0.86 (0.61–1.20) | 0.51 | 3.90 | Current study |
| 0.84 (0.57–1.04) | - | SAIMR | 118.4 | 2.0 | 17.81 | 1.70 (1.49–1.94) | 0.94 | 7.94 | [16] |
| 0.45 (0.43–0.47) | - | Antivipmyn-Africa | 32 | 3.0 | 57.11 | 0.47 (0.47–0.48) | 0.31 | 9.69 | [17] |
| 3.55 (95% CI: NA) | - | VACSERA | NA | NA | Not effective | [18] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, C.H.; Wong, K.Y.; Huang, L.-K.; Tan, K.Y.; Tan, N.H.; Wu, W.-G. Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity. Toxins 2022, 14, 860. https://doi.org/10.3390/toxins14120860
Tan CH, Wong KY, Huang L-K, Tan KY, Tan NH, Wu W-G. Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity. Toxins. 2022; 14(12):860. https://doi.org/10.3390/toxins14120860
Chicago/Turabian StyleTan, Choo Hock, Kin Ying Wong, Li-Kun Huang, Kae Yi Tan, Nget Hong Tan, and Wen-Guey Wu. 2022. "Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity" Toxins 14, no. 12: 860. https://doi.org/10.3390/toxins14120860
APA StyleTan, C. H., Wong, K. Y., Huang, L.-K., Tan, K. Y., Tan, N. H., & Wu, W.-G. (2022). Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity. Toxins, 14(12), 860. https://doi.org/10.3390/toxins14120860