MC-LR Aggravates Liver Lipid Metabolism Disorders in Obese Mice Fed a High-Fat Diet via PI3K/AKT/mTOR/SREBP1 Signaling Pathway

 and
and 
Abstract
1. Introduction
2. Results
2.1. Effects of MC-LR on Body Weight, Food Intake, and Liver Index in HFD-Induced Obese Mice
2.2. Biochemical Indexes
2.3. Pathological Observation of Liver
2.4. Effects of MC-LR on Lipid Synthesis and Fatty Acid β-Oxidation in Liver
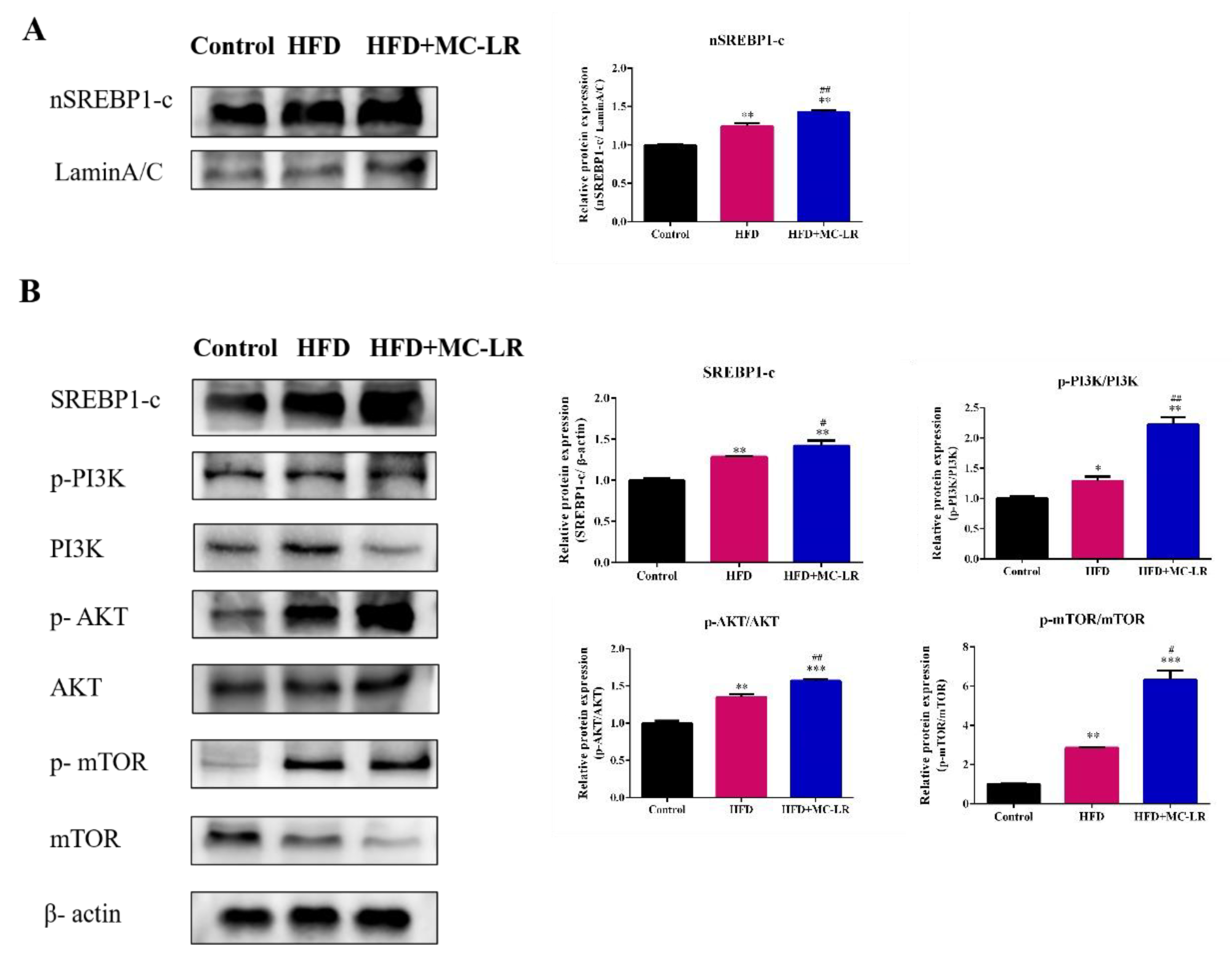
2.5. Effect of MC-LR on PI3K/AKT/mTOR/SREBP1 Pathway in Liver
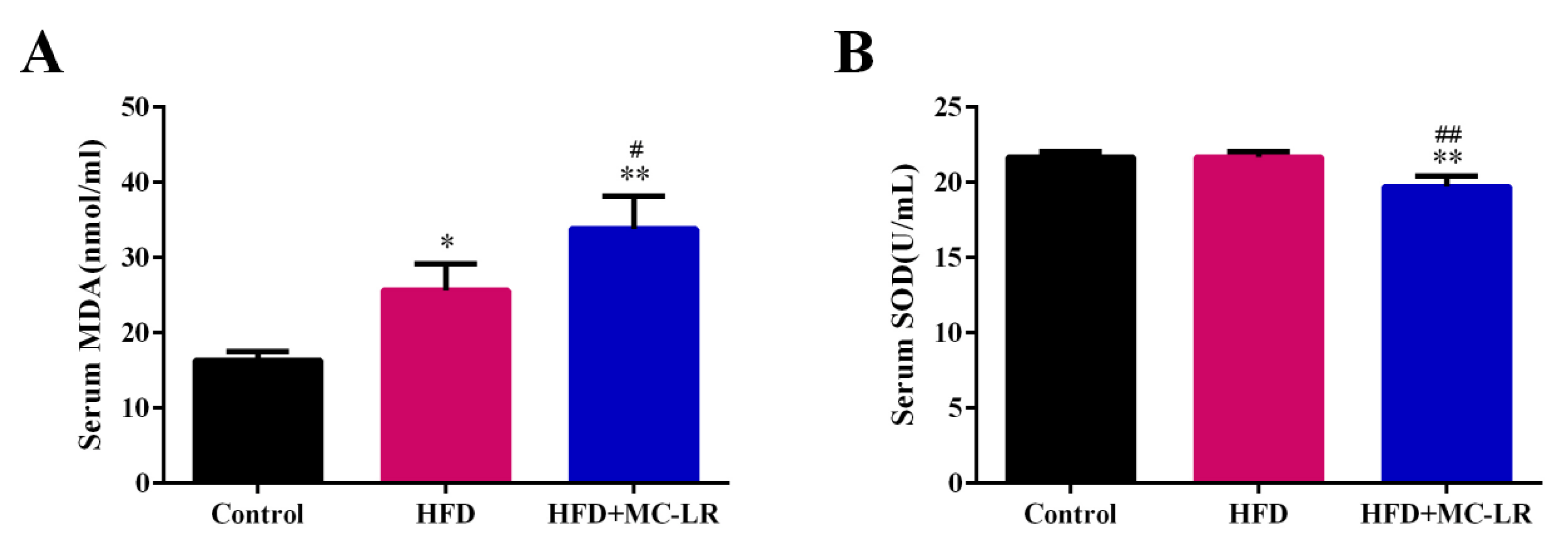
2.6. Effects of MC-LR on Serum Oxidative Stress Status in HFD-Induced Obese Mice
3. Discussion
3.1. Effects of MC-LR on General Conditions in Obese Mice
3.2. MC-LR Aggravates Liver Damage in Obese Mice
3.3. MC-LR Aggravates Liver Lipid Metabolism in Obese Mice
3.4. MC-LR Induced and Aggravated the Serum Oxidative Stress Status
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Animals and Diet
5.3. Detection of Biochemical Indexes
5.4. Hematoxylin and Eosin and Oil Red O Staining
5.5. Quantitative RT-PCR
5.6. Western Blot
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baxter, J.; Armijo, P.R.; Flores, L.; Krause, C.; Samreen, S.; Tanner, T. Updates on Monogenic Obesity in a Multifactorial Disease. Obes. Surg. 2019, 29, 4077–4083. [Google Scholar] [CrossRef] [PubMed]
- Abarca-Gómez, L.; Abdeen, Z.A.; Hamid, Z.A.; Abu-Rmeileh, N.M.; Acosta-Cazares, B.; Acuin, C.; Adams, R.J.; Aekplakorn, W.; Afsana, K.; Aguilar-Salinas, C.A.; et al. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [PubMed]
- Tianyang, H.; Weikang, Z.; Xiangguo, M.; Jianhui, J.; Fuan, Z.; Wanmeng, Z.; Hao, H.; Guozhen, C. Oral administration of Camellia oil ameliorates obesity and modifies the gut microbiota composition in mice fed a high-fat diet. FEMS Microbiol. Lett. 2021, 368, fnab063. [Google Scholar]
- Francesco, V.; Gerdes, J.M. The role of primary cilia in obesity and diabetes. Ann. New York Acad. Sci. 2017, 1391, 71–84. [Google Scholar]
- Parto, P.; Lavie, C.J. Obesity and CardiovascularDiseases. Curr Probl Cardiol 2017, 42, 376–394. [Google Scholar] [CrossRef]
- Tianhua, X.; Zitong, S.; Li, Y. Obesity-related glomerulopathy: Pathogenesis, pathologic, clinical characteristics and treatment. Front. Med. 2017, 11, 340–348. [Google Scholar]
- Piyawan, K.; Sripaiboonkij, T.N.; Pisut, K.; Paweena, S.; Nattachai, S.; Kearkiat, P.; Kriang, T.; Somchai, E.-O. The magnitude of obesity and metabolic syndrome among diabetic chronic kidney disease population: A nationwide study. PLoS ONE 2018, 13, e0196332. [Google Scholar]
- Carbone, S.; Canada, J.M.; Billingsley, H.E.; Siddiqui, M.S.; Elagizi, A.; Lavie, C.J. Obesity paradox in cardiovascular disease: Where do we stand? Vasc. Health Risk Manag. 2019, 15, 89–100. [Google Scholar] [CrossRef]
- Stephanie, A.; Gillian, M.; Tracy, R. Obesity and Cancer Metastasis: Molecular and Translational Perspectives. Cancers 2020, 12, 3798. [Google Scholar]
- Yan, X.; Xu, X.; Wang, M.; Wang, G.; Wu, S.; Li, Z.; Sun, H.; Shi, A.; Yang, Y. Climate warming and cyanobacteria blooms: Looks at their relationships from a new perspective. Water Res. 2017, 125, 449–457. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat Rev Microbiol 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X.; et al. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res 2020, 174, 115638. [Google Scholar] [CrossRef] [PubMed]
- Sangolkar, L.N.; Maske, S.S.; Chakrabarti, T. Methods for determining microcystins (peptide hepatotoxins) and microcystin-producing cyanobacteria. Water Res. 2006, 40, 3485–3496. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Environmental science. Blooms bite the hand that feeds them. Science 2013, 342, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Celine, S.; Kasun, R.; Kanishka, S.; Ajith, K. Microcystins: Biogenesis, toxicity, analysis and control. Chem. Res. Toxicol. 2020, 33, 2225–2246. [Google Scholar]
- Danping, C.; Jia, W.; Feiyu, H.; Hai, F.; Tangjian, P.; Jiayou, L.; Fei, Y. The detoxification activities and mechanisms of microcystinase towards MC-LR. Ecotoxicol. Environ. Saf. 2022, 236, 2225–2246. [Google Scholar]
- Can, D.; Shuilin, Z.; Yue, Y.; Xiangling, F.; Jihua, C.; Yan, T.; Hui, W.; Fei, Y. Chronic exposure to low concentration of MC-LR caused hepatic lipid metabolism disorder. Ecotoxicol. Environ. Saf. 2022, 239, 113649. [Google Scholar]
- Xiping, Y.; Shuaishuai, X.; Feiyu, H.; Cong, W.; Shuilin, Z.; Hai, F.; Jian, G.; Jihua, C.; Xiangling, F.; Yang, F.; et al. Effects of Chronic Exposure to Microcystin-LR on Kidney in Mice. Int. J. Environ. Res. Public Health 2019, 16, 5030. [Google Scholar]
- Yue, Y.; Hui, W.; Xiaoyan, W.; Ling, C.; Wenya, L.; Danping, C.; Shuxiang, D.; Hanyu, C.; Ying, L.; Xiangling, F.; et al. Long-term environmental levels of microcystin-LR exposure induces colorectal chronic inflammation, fibrosis and barrier disruption via CSF1R/Rap1b signaling pathway. J. Hazard. Mater. 2022, 440, 129793. [Google Scholar]
- Lan, G.; Jing, C.; Jian, L.; An-Qi, C.; Wei-Wei, Z.; Xiu-Liang, L.; Jing, W.; Xiao-Yi, Z.; Ye, Z.; Yuan-Hua, C.; et al. Microcystin-LR inhibits testosterone synthesis via reactive oxygen species-mediated GCN2/eIF2α pathway in mouse testes. Sci. Total Environ. 2021, 781, 146730. [Google Scholar]
- Haohao, L.; Xin, Z.; Yueqin, W.; Losiewicz, M.D.; Xinghai, C.; Xingde, D.; Yongshui, W.; Bingyu, Z.; Xing, G.; Shumeng, Y.; et al. Chronic Exposure to Environmentally Relevant Concentrations of Microcystin-Leucine Arginine Causes Lung Barrier Damage through PP2A Activity Inhibition and Claudin1 Ubiquitination. J. Agric. Food Chem. 2022, 70, 10907–10918. [Google Scholar]
- Yaw, M.I.; Xian, Z.; Fei, Y. Importance of bacterial biodegradation and detoxification processes of microcystins for environmental health. J. Toxicol. Environ. Health. Part B Crit. Rev. 2018, 21, 357–369. [Google Scholar]
- Shuidong, F.; Shuxiang, D.; Yan, T.; Ying, L.; Yue, Y.; Shuaishuai, X.; Peng, T.; Yao, L.; Yanying, D.; Jia, W.; et al. Microcystin-LR Combined with Cadmium Exposures and the Risk of Chronic Kidney Disease: A Case-Control Study in Central China. Environ. Sci. Technol. 2022, previous. [Google Scholar]
- Apurva, L.; Breidenbach, J.D.; Su, R.C.; Jordan, M.; Rebecca, K.; Alison, M.; John, N.; Shivani, P.; Prajwal, H.; Mirella, Y.; et al. As We Drink and Breathe: Adverse Health Effects of Microcystins and Other Harmful Algal Bloom Toxins in the Liver, Gut, Lungs and Beyond. Life 2022, 12, 418. [Google Scholar]
- Leticia, D.; Monte, B.M.d.; María, P.; Angeles, J. Immunotoxic Effects Induced by Microcystins and Cylindrospermopsin: A Review. Toxins 2021, 13, 711. [Google Scholar]
- Jiajia, Z.; Jia, W.; Yaw, M.I.; Tangjian, P.; Fei, Y. Immobilization of Microbes for Biodegradation of Microcystins: A Mini Review. Toxins 2022, 14, 573. [Google Scholar]
- Rodríguez, E.M.; Acero, J.L.; Spoof, L.; Meriluoto, J. Oxidation of MC-LR and -RR with chlorine and potassium permanganate: Toxicity of the reaction products. Water Res. 2007, 42, 1744–1752. [Google Scholar] [CrossRef]
- Feiyu, H.; Xiaoyu, L.; Jian, G.; Hai, F.; Fei, Y. Aromatic hydrocarbon compound degradation of phenylacetic acid by indigenous bacterial Sphingopyxis isolated from Lake Taihu. J. Toxicol. Environ. Health. Part A 2019, 82, 1164–1171. [Google Scholar]
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of Microcystin-LR by a Novel Native Effective Bacterial Community Designated as YFMCD4 Isolated from Lake Taihu. Toxins 2018, 10, 363. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Drinking-Water Quality, 2nd ed.; Volume 2-Health Criteria and Other Supporting Information; World Health Organization: Geneva, Switzerland, 2020.
- Grosse, Y.; Baan, R.; Straif, K.; Secretan, B.; Ghissassi, F.E.; Cogliano, V. Carcinogenicity of nitrate, nitrite, and cyanobacterial peptide toxins. Lancet Oncol. 2006, 7, 628–629. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M. Tumor promoters--microcystin-LR, nodularin and TNF-alpha and human cancer development. Anti-Cancer Agents Med. Chem. 2011, 11, 628–629. [Google Scholar] [CrossRef]
- Liang, C.; Ping, X. Mechanisms of Microcystin-induced Cytotoxicity and Apoptosis. Mini Rev. Med. Chem. 2016, 16, 1018–1031. [Google Scholar]
- Clark, S.P.; Davis, M.A.; Ryan, T.P.; Searfoss, G.H.; Hooser, S.B. Hepatic Gene Expression Changes in Mice Associated with Prolonged Sublethal Microcystin Exposure. Toxicol. Pathol. 2007, 35, 594–605. [Google Scholar] [CrossRef]
- Lad, A.; Su, R.C.; Breidenbach, J.D.; Stemmer, P.M.; Carruthers, N.J.; Sanchez, N.K.; Khalaf, F.K.; Zhang, S.; Kleinhenz, A.L.; Dube, P.; et al. Chronic Low Dose Oral Exposure to Microcystin-LR Exacerbates Hepatic Injury in a Murine Model of Non-Alcoholic Fatty Liver Disease. Toxins 2019, 11, 486. [Google Scholar] [CrossRef]
- Arman, T.; Lynch, K.D.; Montonye, M.L.; Goedken, M.; Clarke, J.D. Sub-Chronic Microcystin-LR Liver Toxicity in Preexisting Diet-Induced Nonalcoholic Steatohepatitis in Rats. Toxins 2019, 11, 398. [Google Scholar] [CrossRef]
- Zhao, Y.; Yan, Y.; Xie, L.; Wang, L.; He, Y.; Wan, X.; Xue, Q. Long-term environmental exposure to microcystins increases the risk of nonalcoholic fatty liver disease in humans: A combined fisher-based investigation and murine model study. Environ. Int. 2020, 138, 105648. [Google Scholar] [CrossRef]
- He, J.; Li, G.; Chen, J.; Lin, J.; Zeng, C.; Chen, J.; Deng, J.; Xie, P. Prolonged exposure to low-dose microcystin induces nonalcoholic steatohepatitis in mice: A systems toxicology study. Arch. Toxicol. 2017, 91, 465–480. [Google Scholar] [CrossRef]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Giannuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of Microcystin-LR. Toxicon 2015, 104, 26–33. [Google Scholar] [CrossRef]
- Liang, C.; Yufei, H.; Jun, H.; Jun, C.; Giesy, J.P.; Ping, X. Responses of the Proteome and Metabolome in Livers of Zebrafish Exposed Chronically to Environmentally Relevant Concentrations of Microcystin-LR. Environ. Sci. Technol. 2017, 51, 596–607. [Google Scholar]
- Zhang, Z.; Zhang, X.-X.; Wu, B.; Yin, J.; Yu, Y.; Yang, L. Comprehensive insights into microcystin-LR effects on hepatic lipid metabolism using cross-omics technologies. J. Hazard. Mater. 2016, 315, 126–134. [Google Scholar] [CrossRef]
- Tai, C.-J.; Choong, C.-Y.; Shi, Y.-C.; Lin, Y.-C.; Wang, C.-W.; Lee, B.-H.; Tai, C.-J. Solanum nigrum Protects against Hepatic Fibrosis via Suppression of Hyperglycemia in High-Fat/Ethanol Diet-Induced Rats. Molecules 2016, 21, 269. [Google Scholar] [CrossRef]
- Fawell, J.K. The toxicity of cyanobacterial toxins in the mouse: I Microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef]
- Jensen-Urstad, A.P.L.; Semenkovich, C.F. Fatty acid synthase and liver triglyceride metabolism: Housekeeper or messenger? BBA Mol. Cell Biol. Lipids 2012, 1821, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Højland, I.D.; Jens, L.; Pernille, T.-N. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. CMLS 2018, 75, 3313–3327. [Google Scholar]
- Berlanga, A.; Guiu-Jurado, E.; Porras, J.A.; Auguet, T. Molecular pathways in non-alcoholic fatty liver disease. Clin. Exp. Gastroenterol. 2014, 2014, 221–239. [Google Scholar]
- Iizuka, K.; Horikawa, Y. Chrebp: A Glucose-activated Transcription Factor Involved in the Development of Metabolic Syndrome. Endocr. J. 2008, 55, 221–239. [Google Scholar] [CrossRef]
- Goedeke, L.; Bates, J.; Vatner, D.F.; Perry, R.J.; Wang, T.; Ramirez, R.; Shulman, G.I. Acetyl-CoA Carboxylase Inhibition Reverses NAFLD and Hepatic Insulin Resistance but Promotes Hypertriglyceridemia in Rodents. Hepatology 2018, 68, 221–239. [Google Scholar] [CrossRef]
- Salie, M.J.; Thelen, J.J. Regulation and structure of the heteromeric acetyl-CoA carboxylase. BBA Mol. Cell Biol. Lipids 2016, 1861, 1207–1213. [Google Scholar] [CrossRef]
- Yuki, K.; Cohen, D.E. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J. Gastroenterol. 2013, 48, 1207–1213. [Google Scholar]
- Ricoult, S.J.; Manning, B.D. The multifaceted role of mTORC1 in the control of lipid metabolism. EMBO Rep. 2013, 14, 242–251. [Google Scholar] [CrossRef]
- Du, C.; Wu, M.; Liu, H.; Ren, Y.; Du, Y.; Wu, H.; Wei, J.; Liu, C.; Yao, F.; Wang, H.; et al. Thioredoxin-interacting protein regulates lipid metabolism via Akt/mTOR pathway in diabetic kidney disease. Int J Biochem Cell Biol 2016, 79, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Krycer, J.; Sharpe, L.; Luu, W.; Brown, A. The Akt-SREBP nexus: Cell signaling meets lipid metabolism. Trends Endocrinol. Metab. TEM 2010, 21, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Zhu, J.; Wu, J.; Thompson, C.B.; Jiang, X. Oncogenic activation of PI3K-AKT-mTOR signaling suppresses ferroptosis via SREBP-mediated lipogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 31189–31197. [Google Scholar] [CrossRef] [PubMed]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.R.; Chung, Y.-L.; Schulze, A. SREBP Activity Is Regulated by mTORC1 and Contributes to Akt-Dependent Cell Growth. Cell Metab. 2008, 8, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Wang, Q.; Wei, Y.; Yao, N.; Wu, Z.; Ma, Y.; Lin, Z.; Zhao, M.; Che, C.; Yao, X.; et al. Cholesterol-lowering effects and potential mechanisms of different polar extracts from Cyclocarya paliurus leave in hyperlipidemic mice. J. Ethnopharmacol. 2015, 176, 17–26. [Google Scholar] [CrossRef]
- Gu, Y.; Gao, L.; Han, Q.; Li, A.; Yu, H.; Liu, D.; Pang, Q. GSK-3beta at the Crossroads in Regulating Protein Synthesis and Lipid Deposition in Zebrafish. Cells 2019, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, Q.; Long, X.; Zhang, J.; Huang, X.; Aa, J.; Yang, H.; Chen, Z.; Xing, J. CD147 reprograms fatty acid metabolism in hepatocellular carcinoma cells through Akt/mTOR/SREBP1c and P38/PPARalpha pathways. J Hepatol 2015, 63, 1378–1389. [Google Scholar] [CrossRef]
- Pepino, M.Y.; Kuda, O.; Samovski, D.; Abumrad, N.A. Structure-Function of CD36 and Importance of Fatty Acid Signal Transduction in Fat Metabolism. Annu. Rev. Nutr. 2014, 34, 281–303. [Google Scholar] [CrossRef]
- Coleman, R.A.; Mashek, D.G. Mammalian triacylglycerol metabolism: Synthesis, lipolysis, and signaling. Chem. Rev. 2011, 111, 6359–6386. [Google Scholar] [CrossRef]
- Yen, C.L.E.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V. Thematic review series: Glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J. Lipid Res. 2008, 49, 6359–6386. [Google Scholar] [CrossRef]
- Yi-Hao, Y.; Ginsberg, H. The role of acyl-CoA:diacylglycerol acyltransferase (DGAT) in energy metabolism. Ann. Med. 2004, 36, 252–261. [Google Scholar]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Zhengyan, G.; Rui, J.; Qin, H.; Liping, C.; Jinliang, D.; Wenrong, F.; Galina, J.; Pao, X.; Guojun, Y. Alteration of lipid metabolism, autophagy, apoptosis and immune response in the liver of common carp (Cyprinus carpio) after long-term exposure to bisphenol A. Ecotoxicol. Environ. Saf. 2021, 211, 111923. [Google Scholar]
- Glade, M.J.; Meguid, M.M. A Glance At... ethanol consumption, GSH suppression, and oxidative liver damage. Nutrition 2017, 33, 199–203. [Google Scholar] [CrossRef]
- Bao, W.; Li, K.; Rong, S.; Yao, P.; Hao, L.; Ying, C.; Zhang, X.; Nussler, A.; Liu, L. Curcumin alleviates ethanol-induced hepatocytes oxidative damage involving heme oxygenase-1 induction. J. Ethnopharmacol. 2010, 128, 549–553. [Google Scholar] [CrossRef]
- Fatma, U.; Sevilay, S.; Serpil, E.; Sumeyya, A.; Ferah, A.; Omer, A. The relationship between oxidative stress and nonalcoholic fatty liver disease: Its effects on the development of nonalcoholic steatohepatitis. Redox Rep. Commun. Free Radic. Res. 2013, 18, 127–133. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| SREBP-1c | TGGAGACATCGCAAACAAG | GGTAGACAACAGCCGCATC |
| DGAT2 | GGCTGGCATTTGACTGG | TGGTGGTCAGCAGGTTGT |
| FAS | GCCTCCGTGGACCTTATC | ACAGACACCTTCCCGTCA |
| ACC1 | AAGGGACAGTAGAAATCAAA | CAGCCTCCAGTAGAAGAAG |
| SCD1 | GGGAATAGTCAAGAGGCT | ACGAGGACGACAATACAA |
| DGAT1 | GTGGGTTCCGTGTTTGC | CTCGGTAGGTCAGGTTGTCT |
| PPARγ | TTCGCTGATGCACTGCCTAT | TGATCGCACTTTGGTATTCTTGG |
| CD36 | GGCAGGAGTGCTGGATTA | GAGGCGGGCATAGTATCA |
| CPT1A | ATGTTTCGACAGGTGGTT | TGCGTTTATGCCTATCTT |
| PPARα | AGGGCCTCCCTCCTACGCTTG | GGGTGGCAGGAAGGGAACAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, H.; Du, C.; Yang, Y.; Feng, X.; Zhu, L.; Chen, J.; Yang, F. MC-LR Aggravates Liver Lipid Metabolism Disorders in Obese Mice Fed a High-Fat Diet via PI3K/AKT/mTOR/SREBP1 Signaling Pathway. Toxins 2022, 14, 833. https://doi.org/10.3390/toxins14120833
Chu H, Du C, Yang Y, Feng X, Zhu L, Chen J, Yang F. MC-LR Aggravates Liver Lipid Metabolism Disorders in Obese Mice Fed a High-Fat Diet via PI3K/AKT/mTOR/SREBP1 Signaling Pathway. Toxins. 2022; 14(12):833. https://doi.org/10.3390/toxins14120833
Chicago/Turabian StyleChu, Hanyu, Can Du, Yue Yang, Xiangling Feng, Lemei Zhu, Jihua Chen, and Fei Yang. 2022. "MC-LR Aggravates Liver Lipid Metabolism Disorders in Obese Mice Fed a High-Fat Diet via PI3K/AKT/mTOR/SREBP1 Signaling Pathway" Toxins 14, no. 12: 833. https://doi.org/10.3390/toxins14120833
APA StyleChu, H., Du, C., Yang, Y., Feng, X., Zhu, L., Chen, J., & Yang, F. (2022). MC-LR Aggravates Liver Lipid Metabolism Disorders in Obese Mice Fed a High-Fat Diet via PI3K/AKT/mTOR/SREBP1 Signaling Pathway. Toxins, 14(12), 833. https://doi.org/10.3390/toxins14120833