Can Red Yeast (Sporidiobolus pararoseus) Be Used as a Novel Feed Additive for Mycotoxin Binders in Broiler Chickens?

 ,
,  ,
,  ,
,  ,
, 


Abstract
1. Introduction
2. Results
2.1. Growth Performance
2.2. Blood Parameters
2.2.1. Blood Chemistry
2.2.2. Mycotoxin Determination in Blood Samples
2.3. Relative Organ Weight
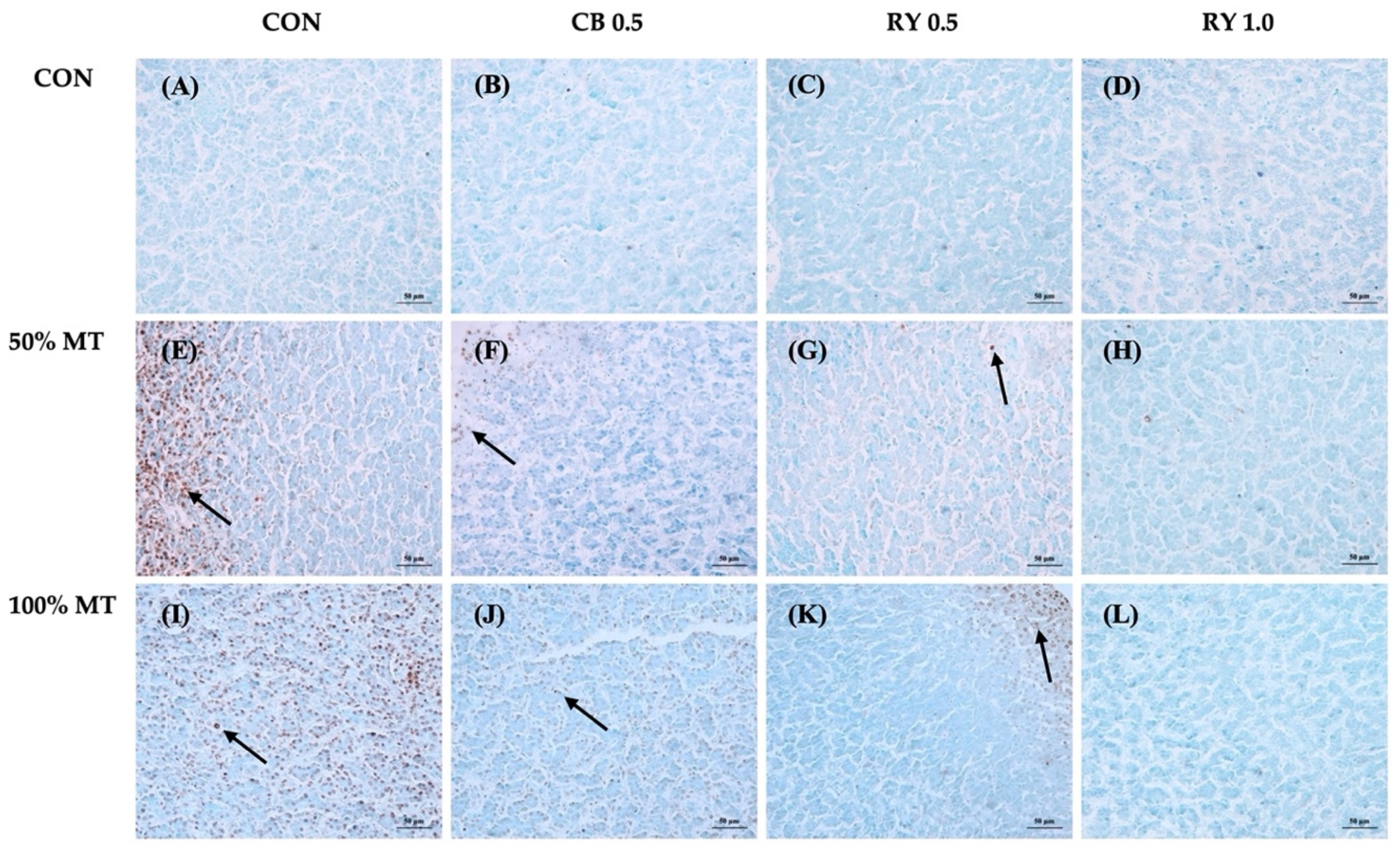
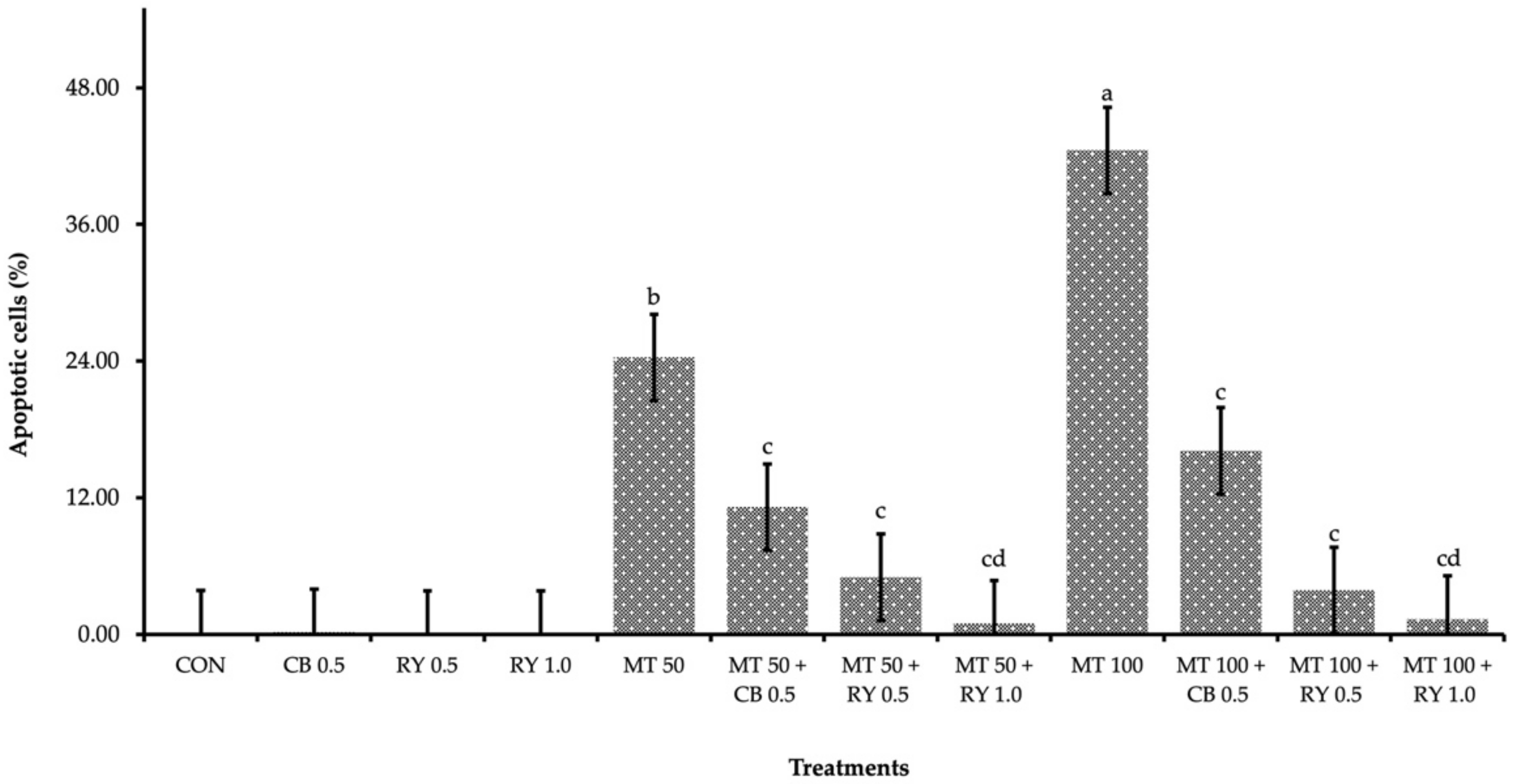
2.4. Apoptotic Cells in Liver Tissue
2.5. Cecal Microbial Populations
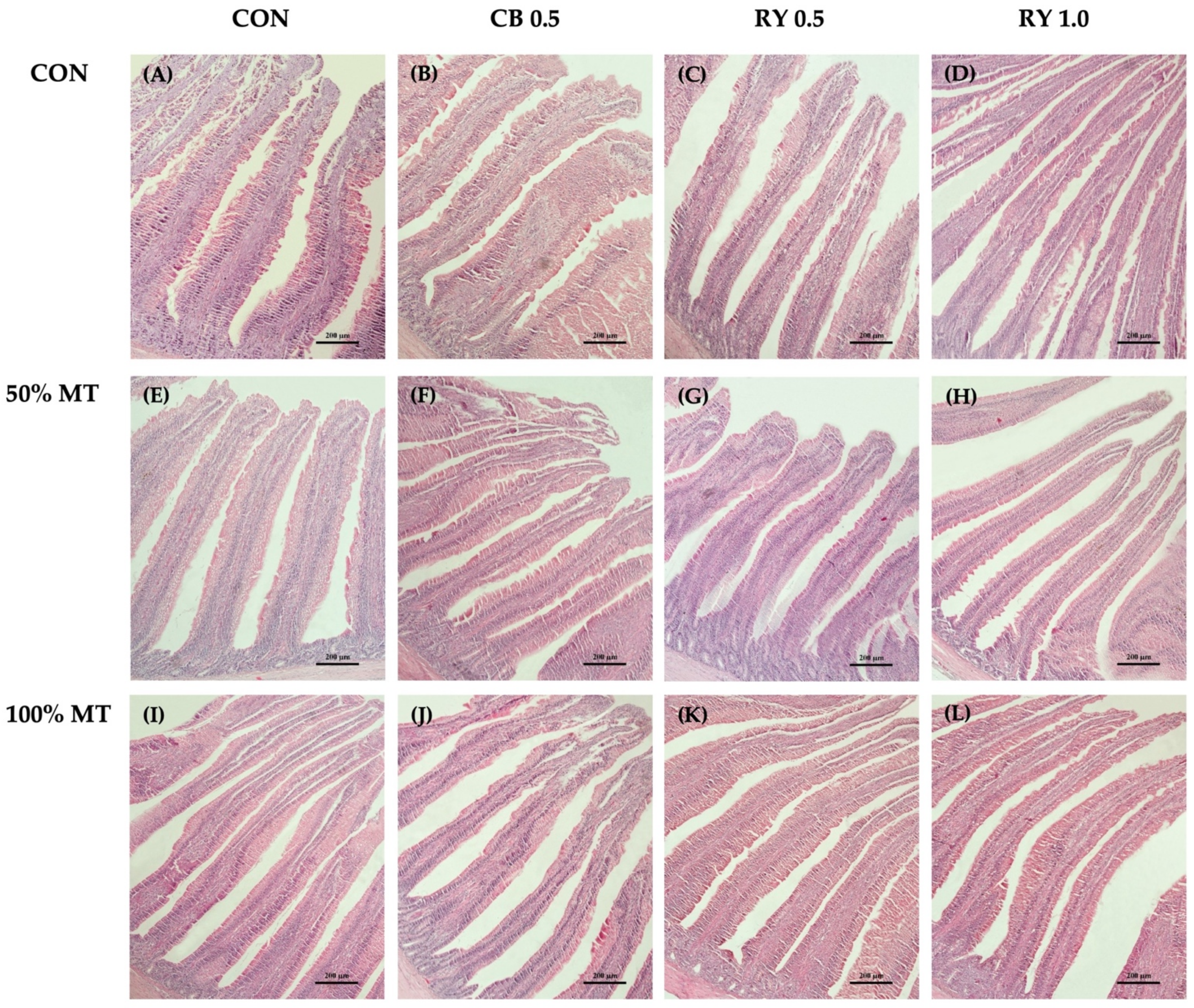
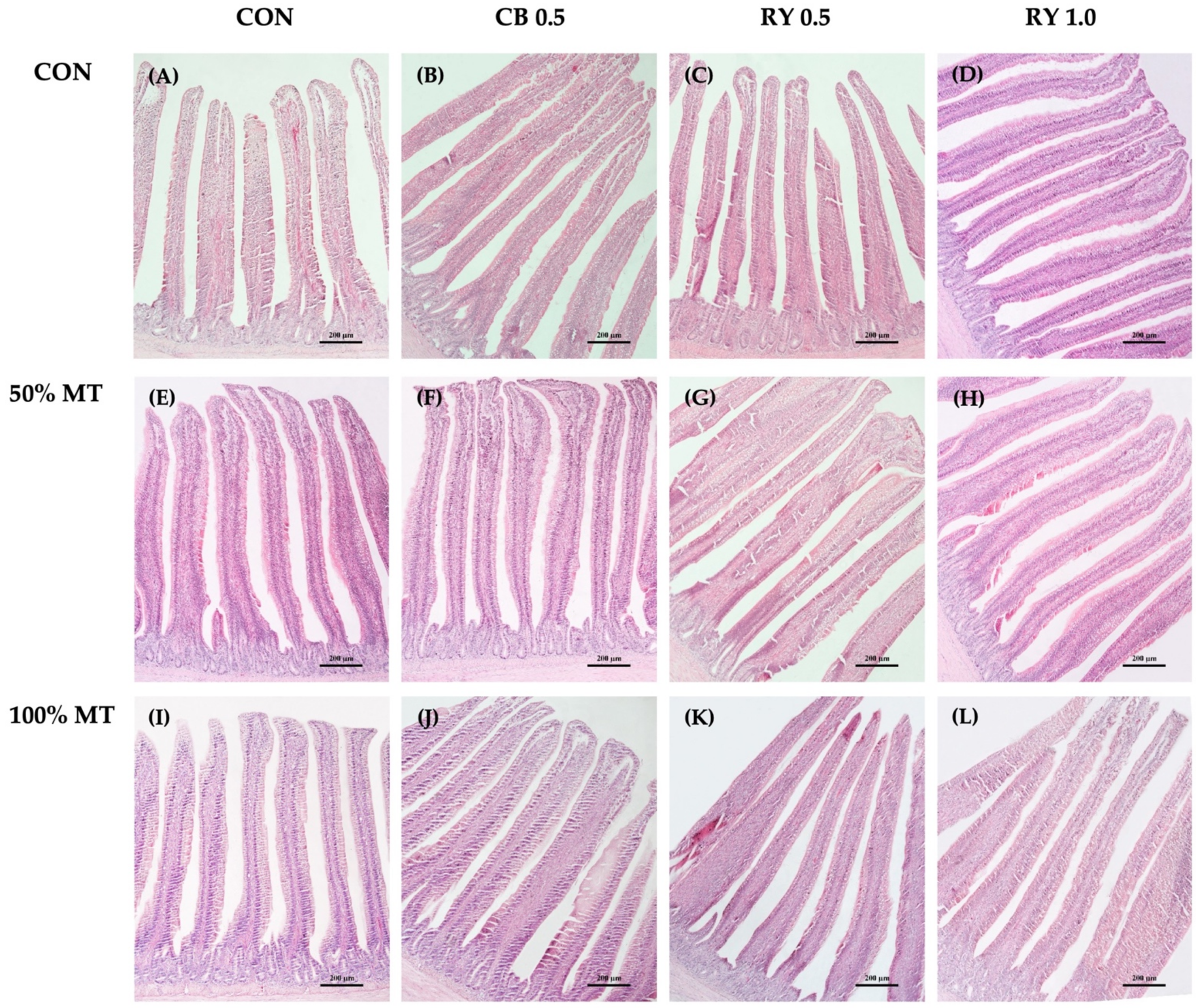
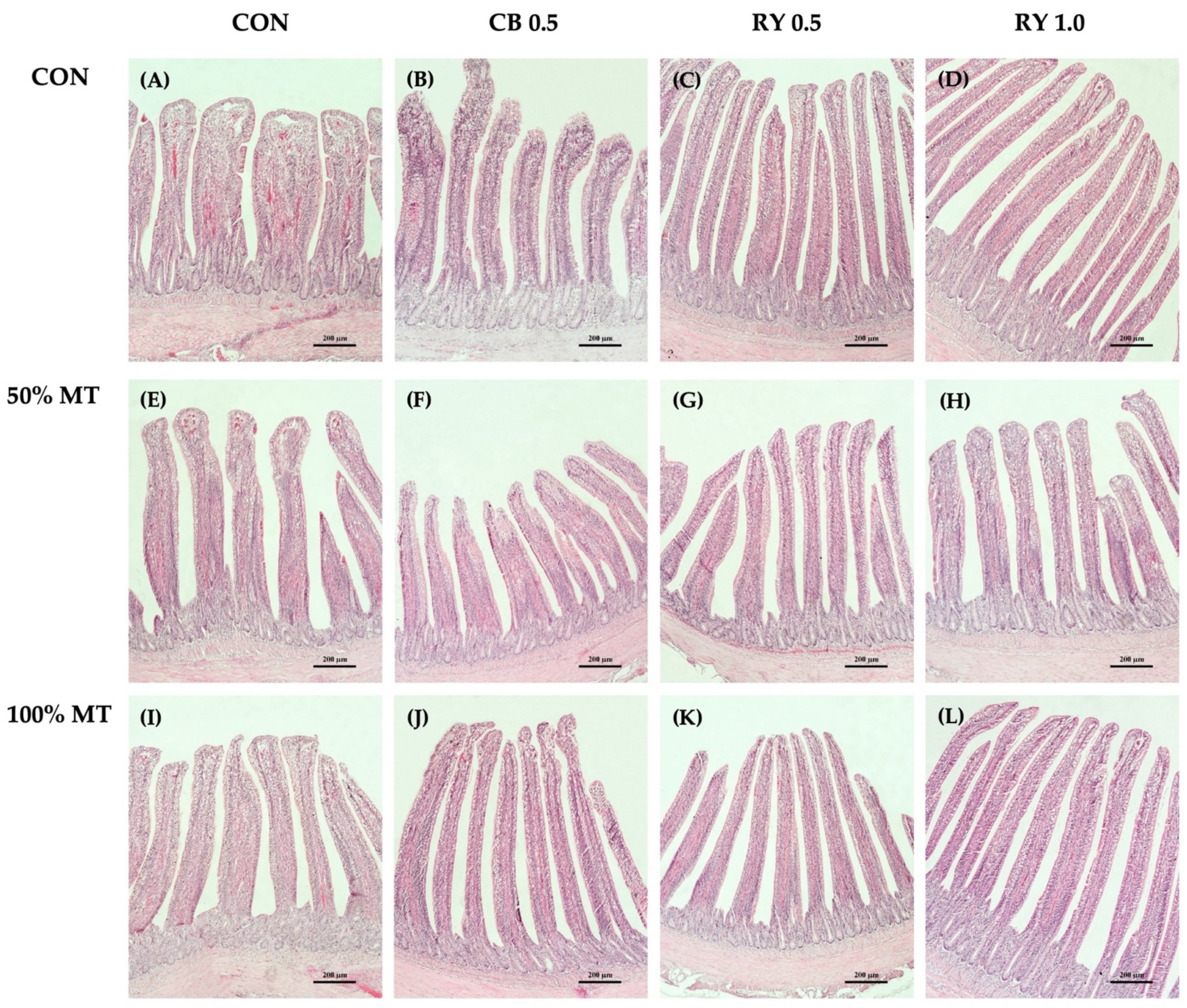
2.6. Intestinal Morphology
2.6.1. Intestinal Morphology in the Duodenum
2.6.2. Intestinal Morphology in the Jejunum
2.6.3. Intestinal Morphology in the Ileum
3. Discussion
3.1. Growth Performance
3.2. Blood Parameters
3.3. Relative Organ Weight and Apoptotic Cells in Liver Tissue
3.4. Cecal Microbial Populations
3.5. Intestinal Morphology
4. Conclusions
5. Materials and Methods
5.1. Preparation of the Red Yeast
5.2. Ethical Approval and Informed Consent
5.3. Animal Management
5.4. Growth Performance
- ADG (g) = BWG/(days of growth period × number of broilers)
- ADFI (g) = feed consumption/(days of growth period × number of broilers)
- FCR = feed consumption (kg)/total weight gain (kg)
5.5. Blood Collection and Analysis
5.6. Mycotoxin Analysis
5.7. The Relative Organ Weights
5.8. Immunohistochemistry of Liver Tissue
5.9. Cecal Microbial Population
5.10. Intestinal Morphology
5.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arif, M.; Iram, A.; Bhutta, M.A.; Naiel, M.A.; El-Hack, A.; Mohamed, E.; Othman, S.I.; Allam, A.A.; Amer, M.S.; Taha, A.E. The biodegradation role of Saccharomyces cerevisiae against harmful effects of mycotoxin contaminated diets on broiler performance, immunity status, and carcass characteristics. Animals 2020, 10, 238. [Google Scholar]
- Osuji, M. Assessment of Factors Affecting Poultry (Broiler) Production in Imo State, Nigeria. Asian J. Agric. Ext. Econ. Sociol. 2019, 35, 1–6. [Google Scholar] [CrossRef]
- Kumar, D.; Kalita, P. Reducing postharvest losses during storage of grain crops to strengthen food security in developing countries. Foods 2017, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [PubMed]
- Eshetu, E.; Habtamu, A.; Gebretensa, A. An overview on major mycotoxin in animal: Its public health implication, economic impact and control strategies. J. Health Med. Nurs. 2016, 25, 64–73. [Google Scholar]
- Yang, C.; Song, G.; Lim, W. Effects of mycotoxin-contaminated feed on farm animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar]
- Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing fungi and mechanisms of phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef]
- Davies, C.R.; Wohlgemuth, F.; Young, T.; Violet, J.; Dickinson, M.; Sanders, J.-W.; Vallieres, C.; Avery, S.V. Evolving challenges and strategies for fungal control in the food supply chain. Fungal Biol. Rev. 2021, 36, 15–26. [Google Scholar] [CrossRef]
- Chen, S.S.; Li, Y.-H.; Lin, M.-F. Chronic exposure to the Fusarium mycotoxin deoxynivalenol: Impact on performance, immune organ, and intestinal integrity of slow-growing chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Sulyok, M.; Krska, R.; Logrieco, A.F.; Vogelgsang, S. Raised concerns about the safety of barley grains and straw: A Swiss survey reveals a high diversity of mycotoxins and other fungal metabolites. Food Control. 2021, 125, 107919. [Google Scholar] [CrossRef]
- Holanda, D.M.; Kim, S.W. Mycotoxin occurrence, toxicity, and detoxifying agents in pig production with an emphasis on deoxynivalenol. Toxins 2021, 13, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Lumsangkul, C.; Chiang, H.-I.; Lo, N.-W.; Fan, Y.-K.; Ju, J.-C. Developmental toxicity of mycotoxin fumonisin b1 in animal embryogenesis: An overview. Toxins 2019, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Lumsangkul, C.; Tso, K.-H.; Fan, Y.-K.; Chiang, H.-I.; Ju, J.-C. Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks. Toxins 2021, 13, 743. [Google Scholar] [CrossRef]
- Hussar, P.; Järveots, T.; Pendovski, L.; Blagoevska, K.; Ristoski, T.; Popovska-Percinic, F. T-2 mycotoxin induced apoptosis in broiler’s liver tissue. Pap. Anthropol. 2018, 27, 9–16. [Google Scholar] [CrossRef][Green Version]
- Wang, H.; Wei, Y.; Xie, Y.; Yan, C.; Du, H.; Li, Z. Ochratoxin A and fumonisin B(1) exhibit synergistic cytotoxic effects by inducing apoptosis on rat liver cells. Toxicon 2020, 181, 19–27. [Google Scholar] [CrossRef]
- Čolović, R.; Puvača, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Đuragić, O.; Kos, J.; Pinotti, L. Decontamination of Mycotoxin-contaminated feedstuffs and compound feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Poughon, L.; Cameleyre, X.; Dussap, C.-G.; François, J.; Bertin, G.; Jouany, J.-P. A novel technique to evaluate interactions between Saccharomyces cerevisiae cell wall and mycotoxins: Application to zearalenone. Biotechnol. Lett. 2003, 25, 783–789. [Google Scholar] [CrossRef]
- Tso, K.-H.; Lumsangkul, C.; Cheng, M.-C.; Ju, J.-C.; Fan, Y.-K.; Chiang, H.-I. Differential Effects of Green Tea Powders on the Protection of Brown Tsaiya and Kaiya Ducklings against Trichothecene T-2 Toxin Toxicity. Animals 2021, 11, 2541. [Google Scholar]
- Tso, K.-H.; Lumsangkul, C.; Ju, J.-C.; Fan, Y.-K.; Chiang, H.-I. The potential of peroxidases extracted from the spent mushroom (Flammulina velutipes) substrate significantly degrade mycotoxin deoxynivalenol. Toxins 2021, 13, 72. [Google Scholar] [CrossRef]
- Kolawole, O.; Graham, A.; Donaldson, C.; Owens, B.; Abia, W.A.; Meneely, J.; Alcorn, M.J.; Connolly, L.; Elliott, C.T. Low doses of mycotoxin mixtures below EU regulatory limits can negatively affect the performance of broiler chickens: A longitudinal study. Toxins 2020, 12, 433. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, C.; Schmidt, J.; Bordignon, H.; Fülber, L.; Layter, J.; Fernandes, J. Efficacy of yeast derived glucomannan or algae-based antioxidant or both as feed additives to ameliorate mycotoxicosis in heat stressed and unstressed broiler chickens. Livest. Sci. 2016, 193, 20–25. [Google Scholar] [CrossRef]
- Kim, S.W.; Holanda, D.M.; Gao, X.; Park, I.; Yiannikouris, A. Efficacy of a yeast cell wall extract to mitigate the effect of naturally co-occurring mycotoxins contaminating feed ingredients fed to young pigs: Impact on gut health, microbiome, and growth. Toxins 2019, 11, 633. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.C.; King, W.; Verax, M.; Fox, U.; Kudupoje, M.B.; Mathis, G.; Lumpkins, B.; Yiannikouris, A. Impact of chronic levels of naturally multi-contaminated feed with Fusarium mycotoxins on broiler chickens and evaluation of the mitigation properties of different titers of yeast cell wall extract. Toxins 2020, 12, 636. [Google Scholar] [CrossRef]
- Tapingkae, W.; Yindee, P.; Moonmanee, T. Effect of dietary red yeast (Sporidiobolus pararoseus) supplementation on small intestinal histomorphometry of laying hens. J. Anim. Plant Sci. 2016, 26, 909–915. [Google Scholar]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Buléon, A.; Jeminet, G.; Canet, I.; François, J.; Bertin, G.; Jouany, J.-P. Comprehensive conformational study of key interactions involved in zearalenone complexation with β-D-glucans. Biomacromolecules 2004, 5, 2176–2185. [Google Scholar]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed. Sci. Technol. 2007, 137, 342–362. [Google Scholar]
- Manowattana, A.; Techapun, C.; Watanabe, M.; Chaiyaso, T. Bioconversion of biodiesel-derived crude glycerol into lipids and carotenoids by an oleaginous red yeast Sporidiobolus pararoseus KM281507 in an airlift bioreactor. J. Biosci. Bioeng. 2018, 125, 59–66. [Google Scholar]
- Ribeiro, D.; Freitas, M.; Silva, A.M.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-A.; Hayden, M.M.; Bannerman, S.; Jansen, J.; Crowe-White, K.M. Anti-apoptotic effects of carotenoids in neurodegeneration. Molecules 2020, 25, 3453. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; Majoka, M.A.; Saha, S.; Garber, A.; Gabarrou, J.-F. Efficacy of dietary antioxidants on broiler oxidative stress, performance and meat quality: Science and market. Avian Biol. Res. 2015, 8, 65–78. [Google Scholar] [CrossRef]
- Fruhauf, S.; Schwartz, H.; Ottner, F.; Krska, R.; Vekiru, E. Yeast cell based feed additives: Studies on aflatoxin B1 and zearalenone. Food Addit. Contam. Part A 2012, 29, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Kanmanee, C.; Srinual, O.; Punyatong, M.; Moonmanee, T.; Lumsangkul, C.; Tangtaweewipat, S.; Van Doan, H.; Yachai, M.; Chaiyaso, T.; Tapingkae, W. Effects of Dietary Supplementation with Red Yeast (Sporidiobolus pararoseus) on Productive Performance, Egg Quality, and Duodenal Cell Proliferation of Laying Hens. Animals 2022, 12, 238. [Google Scholar] [CrossRef]
- Tapingkae, W.; Panyachai, K.; Yachai, M.; Doan, H. Effects of dietary red yeast (Sporidiobolus pararoseus) on production performance and egg quality of laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, e337–e344. [Google Scholar] [CrossRef]
- Tapingkae, W.; Srinual, O.; Lumsangkul, C.; Doan, H.V.; Chiang, H.-I.; Manowattana, A.; Boonchuay, P.; Chaiyaso, T. Industrial-Scale Production of Mycotoxin Binder from the Red Yeast Sporidiobolus pararoseus KM281507. J. Fungi 2022, 8, 353. [Google Scholar]
- Mohaghegh, A.; Chamani, M.; Shivazad, M.; Sadeghi, A.A.; Afzali, N. Effect of esterified glucomannan on broilers exposed to natural mycotoxin-contaminated diets. J. Appl. Anim. Res. 2017, 45, 285–291. [Google Scholar]
- Resanović, R.; Nešić, K.; Nešić, V.; Palić, T.D.; Jaćević, V. Mycotoxins in poultry production. Zb. Matice Srp. Za Prir. Nauk. 2009, 116, 7–14. [Google Scholar]
- Swamy, H.; Smith, T.; Cotter, P.; Boermans, H.; Sefton, A. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on production and metabolism in broilers. Poult. Sci. 2002, 81, 966–975. [Google Scholar]
- Magnoli, A.P.; Poloni, V.L.; Cavaglieri, L. Impact of mycotoxin contamination in the animal feed industry. Curr. Opin. Food Sci. 2019, 29, 99–108. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of zearalenone, aflatoxin B1, and ochratoxin A by yeast-based products: A method for quantification of adsorption performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Matthes, S.; Halle, I.; Ueberschär, K.-H.; Döll, S.; Valenta, H. Effects of graded levels of Fusarium toxin-contaminated wheat and of a detoxifying agent in broiler diets on performance, nutrient digestibility and blood chemical parameters. Br. Poult. Sci. 2003, 44, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, H.; Zhang, K.; Zhang, J.; Hu, P.; Li, Y.; Gu, H.; Liu, H.-Y.; Yang, Z.; Cai, D. Orphan Nuclear Receptor RORγ Modulates the Genome-Wide Binding of the Cholesterol Metabolic Genes during Mycotoxin-Induced Liver Injury. Nutrients 2021, 13, 2539. [Google Scholar] [CrossRef]
- Saminathan, M.; Selamat, J.; Abbasi Pirouz, A.; Abdullah, N.; Zulkifli, I. Effects of nano-composite adsorbents on the growth performance, serum biochemistry, and organ weights of broilers fed with aflatoxin-contaminated feed. Toxins 2018, 10, 345–360. [Google Scholar] [CrossRef]
- Brezina, U.; Rempe, I.; Kersten, S.; Valenta, H.; Humpf, H.U.; Dänicke, S. Diagnosis of intoxications of piglets fed with Fusarium toxin-contaminated maize by the analysis of mycotoxin residues in serum, liquor and urine with LC-MS/MS. Arch. Anim. Nutr. 2014, 68, 425–447. [Google Scholar] [CrossRef]
- Lauwers, M.; De Baere, S.; Letor, B.; Rychlik, M.; Croubels, S.; Devreese, M. Multi LC-MS/MS and LC-HRMS methods for determination of 24 mycotoxins including major phase I and II biomarker metabolites in biological matrices from pigs and broiler chickens. Toxins 2019, 11, 171. [Google Scholar] [CrossRef]
- Mazur-Kuśnirek, M.; Antoszkiewicz, Z.; Lipiński, K.; Fijałkowska, M.; Purwin, C.; Kotlarczyk, S. The effect of polyphenols and vitamin E on the antioxidant status and meat quality of broiler chickens fed diets naturally contaminated with ochratoxin A. Arch. Anim. Nutr. 2019, 73, 431–444. [Google Scholar] [CrossRef]
- Hoerr, F.J. Mycotoxicoses. Dis. Poult. 2020, 31, 1330–1348. [Google Scholar]
- Naseem, M.N.; Saleemi, M.K.; Khan, A.; Khatoon, A.; Gul, S.T.; Rizvi, F.; Ahmad, I.; Fayyaz, A. Pathological effects of concurrent administration of aflatoxin B1 and fowl adenovirus-4 in broiler chicks. Microb. Pathog. 2018, 121, 147–154. [Google Scholar] [CrossRef]
- Shin, J.; Song, M.-H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-oxidant actions of carotenoids in triggering apoptosis of cancer cells: A review of emerging evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Feng, Y.; Wei, J.-T.; Zhu, M.-X.; Zhang, L.; Zhang, J.-C.; Karrow, N.A.; Han, Y.-M.; Wu, Y.-Y.; Guo, Y.-M.; et al. Mitigation effects of bentonite and yeast cell wall binders on AFB1, DON, and OTA induced changes in laying hen performance, egg quality, and health. Toxins 2021, 13, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Kittichaiworakul, R.; Taya, S.; Chariyakornkul, A.; Chaiyaso, T.; Wongpoomchai, R. Antigenotoxic Effects and Possible Mechanism of Red Yeast (Sporidiobolus pararoseus) on Aflatoxin B1-Induced Mutagenesis. Biomolecules 2021, 11, 734. [Google Scholar] [CrossRef] [PubMed]
- Guerre, P. Mycotoxin and gut microbiota interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, E.; Mahdavi, A.; Asgary, S.; Jahanian, R. Effect of dietary supplementation of mannanoligosaccharides on growth performance, ileal microbial counts, and jejunal morphology in broiler chicks exposed to aflatoxins. Livest. Sci. 2016, 190, 123–130. [Google Scholar] [CrossRef]
- Śliżewska, K.; Markowiak-Kopeć, P.; Sip, A.; Lipiński, K.; Mazur-Kuśnirek, M. The effect of using new synbiotics on the turkey performance, the intestinal microbiota and the fecal enzymes activity in turkeys fed ochratoxin a contaminated feed. Toxins 2020, 12, 578. [Google Scholar] [CrossRef]
- Sheth, R.U.; Li, M.; Jiang, W.; Sims, P.A.; Leong, K.W.; Wang, H.H. Spatial metagenomic characterization of microbial biogeography in the gut. Nat. Biotechnol. 2019, 37, 877–883. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Zutter, L.; Houf, K.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Strategies to control Salmonella in the broiler production chain. World’s Poult. Sci. J. 2009, 65, 367–392. [Google Scholar] [CrossRef]
- Wang, A.; Hogan, N.S. Performance effects of feed-borne Fusarium mycotoxins on broiler chickens: Influences of timing and duration of exposure. Anim. Nutr. 2019, 5, 32–40. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, X.; Fang, J.; Cui, H.; Zuo, Z.; Chen, Z. Effects of aflatoxin B 1 exposure and sodium selenite supplementation on the histology, cell proliferation, and cell cycle of jejunum in broilers. Biol. Trace Elem. Res. 2014, 160, 32–40. [Google Scholar] [CrossRef]
- Applegate, T.; Schatzmayr, G.; Pricket, K.; Troche, C.; Jiang, Z. Effect of aflatoxin culture on intestinal function and nutrient loss in laying hens. Poult. Sci. 2009, 88, 1235–1241. [Google Scholar] [PubMed]
- Awad, W.A.; Ghareeb, K.; Böhm, J. The toxicity of Fusarium mycotoxin deoxynivalenol in poultry feeding. World’s Poult. Sci. J. 2012, 68, 651–668. [Google Scholar] [CrossRef]
- Wang, A.; Schwean-Lardner, K.; Hogan, N. Feed preference and feeding behaviours in grower broilers fed diets containing wheat naturally contaminated with fusarium mycotoxins. Br. Poult. Sci. 2019, 60, 309–316. [Google Scholar]
- Ruttanavut, J.; Yamauchi, K. Growth performance and histological alterations of intestinal villi in broilers fed dietary mixed minerals. Asian J. Anim. Sci. 2010, 4, 96–106. [Google Scholar]
- Chaiyaso, T.; Manowattana, A. Enhancement of carotenoids and lipids production by oleaginous red yeast Sporidiobolus pararoseus KM281507. Prep. Biochem. Biotechnol. 2018, 48, 13–23. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Poultry: Ninth Revised Edition; The National Academies Press: Washington, DC, USA, 1994; p. 176.
- Ali Rajput, S.; Sun, L.; Zhang, N.; Mohamed Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.A.; Zhang, J.; Qi, D. Ameliorative effects of grape seed proanthocyanidin extract on growth performance, immune function, antioxidant capacity, biochemical constituents, liver histopathology and aflatoxin residues in broilers exposed to aflatoxin B1. Toxins 2017, 9, 371. [Google Scholar]
- De Castro Ribeiro, M.; Coutinho, L.M.B.; Hilbig, A. The role of apoptosis, cell proliferation index, bcl-2, and p53 in glioblastoma prognosis. Arq. De Neuro-Psiquiatr. 2004, 62, 262–270. [Google Scholar]
- Giannenas, I.; Tontis, D.; Tsalie, E.; Chronis, E.; Doukas, D.; Kyriazakis, I. Influence of dietary mushroom Agaricus bisporus on intestinal morphology and microflora composition in broiler chickens. Res. Vet. Sci. 2010, 89, 78–84. [Google Scholar]
- Srinual, O.; Punyatong, M.; Moonmanee, T.; Intawicha, P.; Yachai, M.; Tapingkae, W. Replacement of fish meal with suckermouth armored catfish and its effect on performance and intestinal morphology of indigenous Thai chicken. J. Anim. Plant Sci. 2020, 30, 803–810. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxins Diets | Adsorbent Level (g kg−1) | Growth Performance | |||||
|---|---|---|---|---|---|---|---|
| Initial Weight (g) | Final Weight (g) | ADG 1 (g) | ADFI (g) | FCR | Mortality (%) | ||
| 8 to 21 d of age | |||||||
| CON 2 | 0 | 184.42 | 550.78 | 26.17 | 38.95 a | 1.49 | 2.50 |
| CB 0.5 | 184.29 | 568.29 | 27.43 | 38.32 a | 1.40 | 0.00 | |
| RY 0.5 | 182.96 | 584.54 | 28.68 | 39.11 a | 1.36 | 0.00 | |
| RY 1.0 | 182.58 | 601.71 | 29.94 | 38.50 a | 1.29 | 0.00 | |
| 50% MT | 0 | 183.63 | 530.83 | 24.80 | 37.96 ab | 1.53 | 5.00 |
| CB 0.5 | 182.92 | 535.00 | 25.14 | 38.12 a | 1.52 | 0.00 | |
| RY 0.5 | 184.29 | 542.58 | 25.59 | 38.38 a | 1.50 | 0.83 | |
| RY 1.0 | 184.79 | 555.04 | 26.45 | 37.89 ab | 1.43 | 0.00 | |
| 100% MT | 0 | 184.42 | 517.19 | 23.77 | 36.41 b | 1.53 | 7.50 |
| CB 0.5 | 183.67 | 557.50 | 26.70 | 38.01 ab | 1.42 | 0.00 | |
| RY 0.5 | 184.08 | 557.29 | 26.66 | 38.13 a | 1.43 | 0.00 | |
| RY 1.0 | 184.63 | 561.16 | 26.90 | 37.94 ab | 1.41 | 0.00 | |
| SEM 3 | 0.74 | 4.06 | 0.28 | 0.11 | 0.05 | 0.33 | |
| Source of variation | p-value | ||||||
| MT | 0.945 | <0.001 | <0.001 | <0.001 | 0.210 | 0.288 | |
| Adsorbent | 0.995 | 0.003 | 0.002 | 0.067 | 0.051 | <0.001 | |
| MT × Adsorbent | 0.995 | 0.783 | 0.703 | 0.040 | 0.890 | 0.282 | |
| 21 to 35 d of age | |||||||
| CON 2 | 0 | 550.78 | 1335.02 | 56.02 | 66.23 | 1.18 | 0.00 c |
| CB 0.5 | 568.29 | 1377.76 | 57.82 | 68.59 | 1.19 | 0.83 c | |
| RY 0.5 | 584.54 | 1409.25 | 58.91 | 67.62 | 1.15 | 0.00 c | |
| RY 1.0 | 601.71 | 1460.74 | 61.36 | 69.43 | 1.13 | 1.67 c | |
| 50% MT | 0 | 530.83 | 1255.96 | 51.79 | 67.81 | 1.31 | 9.47 b |
| CB 0.5 | 535.00 | 1331.36 | 56.88 | 67.79 | 1.19 | 2.50 c | |
| RY 0.5 | 542.58 | 1360.53 | 58.42 | 66.40 | 1.14 | 0.83 c | |
| RY 1.0 | 555.04 | 1390.24 | 59.66 | 67.53 | 1.13 | 0.83 c | |
| 100% MT | 0 | 517.19 | 1138.28 | 44.36 | 65.71 | 1.48 | 14.24 a |
| CB 0.5 | 557.50 | 1330.09 | 55.19 | 68.84 | 1.25 | 0.83 c | |
| RY 0.5 | 557.29 | 1343.07 | 56.13 | 66.92 | 1.19 | 0.83 c | |
| RY 1.0 | 561.16 | 1364.66 | 57.39 | 68.61 | 1.20 | 1.67 c | |
| SEM 3 | 4.06 | 9.64 | 0.53 | 0.31 | 0.02 | 0.41 | |
| Source of variation | p-value | ||||||
| MT | <0.001 | <0.001 | <0.001 | 0.723 | 0.096 | 0.001 | |
| Adsorbent | 0.003 | <0.001 | <0.001 | 0.061 | 0.461 | <0.001 | |
| MT × Adsorbent | 0.783 | 0.089 | 0.221 | 0.655 | 0.979 | <0.001 | |
| 8 to 35 d of age | |||||||
| CON 2 | 0 | 184.42 | 1335.02 b | 41.09 b | 52.59 | 1.28 | 2.50 c |
| CB 0.5 | 184.29 | 1377.76 b | 42.62 ab | 53.45 | 1.25 | 0.83 c | |
| RY 0.5 | 182.96 | 1409.25 ab | 43.79 ab | 53.37 | 1.22 | 0.00 c | |
| RY 1.0 | 182.58 | 1460.74 a | 45.65 a | 53.96 | 1.18 | 1.67 c | |
| 50% MT | 0 | 183.63 | 1255.96 c | 38.30 c | 52.88 | 1.38 | 14.47 b |
| CB 0.5 | 182.92 | 1331.36 bc | 41.02 b | 52.99 | 1.29 | 2.50 c | |
| RY 0.5 | 184.29 | 1360.53 b | 42.01 b | 52.39 | 1.25 | 1.67 c | |
| RY 1.0 | 184.79 | 1390.24 ab | 43.05 ab | 52.71 | 1.22 | 0.83 c | |
| 100% MT | 0 | 184.42 | 1138.28 d | 34.07 d | 51.06 | 1.50 | 21.74 a |
| CB 0.5 | 183.67 | 1330.09 bc | 40.94 b | 53.42 | 1.30 | 0.83 c | |
| RY 0.5 | 184.08 | 1343.07 b | 41.39 b | 52.52 | 1.27 | 0.83 c | |
| RY 1.0 | 184.63 | 1364.66 b | 42.14 b | 53.28 | 1.26 | 1.67 c | |
| SEM 3 | 0.74 | 9.64 | 0.26 | 0.16 | 0.02 | 0.49 | |
| Source of variation | p-value | ||||||
| MT | 0.945 | <0.001 | <0.001 | 0.120 | 0.066 | <0.001 | |
| Adsorbent | 0.995 | <0.001 | <0.001 | 0.041 | 0.008 | <0.001 | |
| MT × Adsorbent | 1.000 | <0.001 | <0.001 | 0.063 | 0.052 | <0.001 | |
| Toxins Diets | Adsorbent level (g kg−1) | Blood Chemistry | ||||||
|---|---|---|---|---|---|---|---|---|
| ALT 1 | Albumin | ALP | CL | TP | BUN | SOD | ||
| CON 2 | 0 | 1.67 | 0.97 | 1364.17 | 0.26 | 2.60 | 1.93 | 0.33 |
| CB 0.5 | 1.67 | 0.87 | 1412.83 | 0.25 | 2.60 | 1.82 | 0.42 | |
| RY 0.5 | 1.00 | 0.97 | 1521.33 | 0.27 | 2.80 | 2.30 | 0.50 | |
| RY 1.0 | 0.67 | 0.93 | 1572.17 | 0.27 | 2.58 | 1.80 | 0.51 | |
| 50% MT | 0 | 1.83 | 0.83 | 1789.83 | 0.26 | 2.43 | 1.95 | 0.10 |
| CB 0.5 | 0.83 | 0.97 | 1612.00 | 0.25 | 2.90 | 2.22 | 0.33 | |
| RY 0.5 | 0.17 | 0.98 | 1607.67 | 0.26 | 2.87 | 2.03 | 0.47 | |
| RY 1.0 | 0.67 | 0.93 | 1639.33 | 0.25 | 2.65 | 1.92 | 0.19 | |
| 100% MT | 0 | 2.17 | 1.02 | 1988.00 | 0.28 | 2.90 | 2.03 | 0.26 |
| CB 0.5 | 0.67 | 1.03 | 1559.83 | 0.27 | 2.92 | 1.78 | 0.22 | |
| RY 0.5 | 0.67 | 0.93 | 1504.67 | 0.26 | 2.62 | 1.92 | 0.47 | |
| RY 1.0 | 0.50 | 1.05 | 1445.50 | 0.26 | 2.90 | 2.03 | 0.37 | |
| SEM 3 | 0.08 | 0.02 | 76.71 | 0.01 | 0.04 | 0.04 | 0.02 | |
| Source of variation | p-value | |||||||
| MT | <0.001 | 0.118 | 0.550 | 0.113 | 0.158 | 0.650 | 0.199 | |
| Adsorbent | 0.451 | 0.922 | 0.811 | 0.596 | 0.523 | 0.471 | 0.202 | |
| MT × Adsorbent | 0.670 | 0.303 | 0.883 | 0.365 | 0.139 | 0.111 | 0.106 | |
| Toxins F8Diets | Adsorbent Level (g kg−1) | Mycotoxin in Blood Samples | ||||
|---|---|---|---|---|---|---|
| AFB1 3 | ZEN | OTA | T-2 | DON | ||
| CON 1 | 0 | ND | ND | ND | ND | ND |
| CB 0.5 | ND | ND | ND | ND | ND | |
| RY 0.5 | ND | ND | ND | ND | ND | |
| RY 1.0 | ND | ND | ND | ND | ND | |
| 50% MT | 0 | 0.45 b | 1.13 b | 1.84 b | 0.62 b | 0.84 b |
| CB 0.5 | 0.05 d | 0.01 c | 0.01 d | 0.01 c | 0.13 c | |
| RY 0.5 | 0.12 c | 0.04 c | 0.05 d | 0.01 c | 0.03 d | |
| RY 1.0 | 0.13 c | 0.01 c | 0.01 d | 0.01 c | 0.12 c | |
| 100% MT | 0 | 1.26 a | 2.74 a | 4.32 a | 1.04 a | 1.08 a |
| CB 0.5 | 0.32 a | 0.04 c | 0.08 d | 0.01 c | 0.12 c | |
| RY 0.5 | 0.13 c | 0.01 c | 0.47 c | 0.01 c | 0.07 d | |
| RY 1.0 | 0.10 c | 0.04 c | 0.18 d | 0.03 c | 0.08 cd | |
| SEM 2 | 0.01 | 0.02 | 0.02 | 0.01 | 0.01 | |
| Source of variation | p-value | |||||
| MT | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Adsorbent | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| MT × Adsorbent | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Toxins Diets | Adsorbent Level (g kg−1) | Relative Organ Weight (%) | ||||
|---|---|---|---|---|---|---|
| Spleen | Kidney | Liver | Heart | Pancreases | ||
| CON 1 | 0 | 0.13 | 0.44 c | 2.33 b | 0.71 | 0.27 c |
| CB 0.5 | 0.16 | 0.70 ab | 2.63 b | 0.80 | 0.36 bc | |
| RY 0.5 | 0.14 | 0.71 ab | 2.49 b | 0.77 | 0.33 c | |
| RY 1.0 | 0.16 | 0.67 ab | 2.53 b | 0.73 | 0.36 bc | |
| 50% MT | 0 | 0.15 | 0.74 ab | 3.01 b | 0.67 | 0.44 b |
| CB 0.5 | 0.15 | 0.72 ab | 2.83 b | 0.77 | 0.33c | |
| RY 0.5 | 0.15 | 0.62 b | 2.64 b | 0.69 | 0.32 c | |
| RY 1.0 | 0.12 | 0.68 ab | 2.58 b | 0.81 | 0.34 c | |
| 100% MT | 0 | 0.12 | 0.81 a | 3.96 a | 0.60 | 0.53 a |
| CB 0.5 | 0.12 | 0.57 bc | 2.99 b | 0.79 | 0.31 c | |
| RY 0.5 | 0.11 | 0.59 bc | 2.67 b | 0.66 | 0.31 c | |
| RY 1.0 | 0.15 | 0.60 bc | 2.50 b | 0.74 | 0.33 c | |
| SEM 2 | 0.01 | 0.01 | 0.07 | 0.01 | 0.01 | |
| Source of variation | p-value | |||||
| MT | 0.240 | 0.265 | 0.011 | 0.191 | 0.106 | |
| Adsorbent | 0.806 | 0.950 | 0.027 | 0.003 | 0.001 | |
| MT × Adsorbent | 0.287 | <0.001 | 0.038 | 0.434 | <0.001 | |
| Toxins Diets | Adsorbent Level (g kg−1) | Bacteria Populations (log10 CFU g−1) | ||
|---|---|---|---|---|
| E. coli | Lactobacillus sp. | Bifidobacterium sp. | ||
| CON 1 | 0 | 5.98 e | 6.47 b | 5.81 b |
| CB 0.5 | 5.81 e | 7.24 a | 5.50 c | |
| RY 0.5 | 7.12 b | 7.13 a | 6.04 ab | |
| RY 1.0 | 6.23 d | 7.58 a | 6.28 a | |
| 50% MT | 0 | 7.04 b | 5.82 d | 4.98 d |
| CB 0.5 | 6.72 c | 6.12 c | 5.47 c | |
| RY 0.5 | 5.96 e | 6.38 b | 5.78 b | |
| RY 1.0 | 5.26 f | 6.56 b | 6.16 a | |
| 100% MT | 0 | 7.69 a | 4.24 g | 4.08 e |
| CB 0.5 | 7.00 b | 5.24 ef | 5.08 d | |
| RY 0.5 | 7.08 b | 5.17 f | 5.11 d | |
| RY 1.0 | 6.41 d | 5.43 e | 5.14 d | |
| SEM 2 | 0.02 | 0.02 | 0.02 | |
| Source of variation | p-value | |||
| MT | <0.001 | <0.001 | <0.001 | |
| Adsorbent | <0.001 | <0.001 | <0.001 | |
| MT × Adsorbent | <0.001 | 0.004 | <0.001 | |
| Toxins Diets | Adsorbent Level (g kg−1) | Duodenum Morphology | |||||
|---|---|---|---|---|---|---|---|
| VH 1(µm) | VW (µm) | CD (µm) | MMT (µm) | VH:CD | VSA (mm2) | ||
| CON 2 | 0 | 4681.71 e | 464.22 d | 603.52 bc | 84.09 cd | 7.85 cd | 6.87 f |
| CB 0.5 | 5239.65 d | 521.72 cd | 556.56 bc | 65.74 ef | 9.77 abc | 8.64 de | |
| RY 0.5 | 5727.52 bc | 488.72 cd | 677.41 abc | 64.00 ef | 9.13 bc | 8.78 de | |
| RY 1.0 | 6109.09 a | 547.70 bc | 535.46 c | 62.06 ef | 11.76 a | 10.51 ab | |
| 50% MT | 0 | 5497.58 d | 476.29 d | 667.92 abc | 120.64 a | 8.34 cd | 8.22 de |
| CB 0.5 | 5735.48 bc | 623.02 a | 614.96 abc | 105.95 b | 10.85 ab | 11.23 a | |
| RY 0.5 | 5646.32 bc | 638.80 a | 711.66 ab | 71.88 de | 8.21 cd | 11.35 a | |
| RY 1.0 | 5892.72 ab | 594.72 ab | 601.35 bc | 56.28 ef | 10.59 ab | 11.01 a | |
| 100% MT | 0 | 5179.44 d | 497.21 cd | 774.36 a | 101.72 b | 6.70 d | 8.08 e |
| CB 0.5 | 5742.89 bc | 508.08 cd | 574.42 bc | 91.56 bc | 10.63 ab | 9.15 cde | |
| RY 0.5 | 5938.88 ab | 549.19 bc | 559.76 bc | 51.92 f | 10.82 ab | 10.28 abc | |
| RY 1.0 | 5885.79 ab | 511.34 cd | 642.44 abc | 65.77 ef | 9.39 bc | 9.44 bcd | |
| SEM 3 | 33.73 | 5.66 | 10.07 | 1.49 | 0.18 | 0.12 | |
| Source of variation | p-value | ||||||
| MT | 0.003 | <0.001 | 0.060 | <0.001 | 0.867 | <0.001 | |
| Adsorbent | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 | <0.001 | |
| MT × Adsorbent | <0.001 | 0.001 | 0.001 | <0.001 | 0.004 | 0.010 | |
| Toxins Diets | Adsorbent Level (g kg−1) | Jejunum Morphology | |||||
|---|---|---|---|---|---|---|---|
| VH 1 (µm) | VW (µm) | CD (µm) | MMT (µm) | VH:CD | VSA (mm2) | ||
| CON 2 | 0 | 4756.42 d | 511.46 | 551.77 c | 80.95 bc | 9.68 a | 7.65 |
| CB 0.5 | 5772.65 a | 420.46 | 669.32 ab | 79.52 bc | 8.95 a | 7.64 | |
| RY 0.5 | 4502.82 de | 447.13 | 551.03 c | 89.41 bc | 8.63 a | 6.46 | |
| RY 1.0 | 5599.24 a | 500.23 | 616.07 bc | 80.98 bc | 9.56 a | 8.74 | |
| 50% MT | 0 | 4148.24 e | 471.166 | 526.90 c | 99.81 b | 7.89 ab | 6.14 |
| CB 0.5 | 4807.77 cd | 447.21 | 537.97 c | 71.87 c | 9.18 a | 6.80 | |
| RY 0.5 | 4519.19 de | 402.66 | 563.37 c | 89.84 bc | 8.40 a | 5.70 | |
| RY 1.0 | 5381.33 ab | 432.62 | 594.32 bc | 94.52 bc | 9.72 a | 7.32 | |
| 100% MT | 0 | 4296.48 e | 462.96 | 724.50 a | 130.66 a | 6.22 b | 6.26 |
| CB 0.5 | 5203.37 c | 455.07 | 625.78 bc | 83.10 bc | 8.49 a | 7.55 | |
| RY 0.5 | 5480.09 b | 419.39 | 609.82 bc | 102.84 b | 9.11 a | 7.19 | |
| RY 1.0 | 5476.95 ab | 440.97 | 572.36 bc | 94.12 bc | 9.88 a | 7.60 | |
| SEM 3 | 41.57 | 5.02 | 9.28 | 1.56 | 0.17 | 0.11 | |
| Source of variation | p-value | ||||||
| MT | <0.001 | 0.028 | 0.004 | <0.001 | 0.195 | <0.001 | |
| Adsorbent | <0.001 | <0.001 | 0.564 | 0.323 | 0.005 | <0.001 | |
| MT × Adsorbent | <0.001 | 0.077 | 0.002 | <0.001 | 0.029 | 0.103 | |
| Toxins Diets | Adsorbent Level (g kg−1) | Ileum Morphology | |||||
|---|---|---|---|---|---|---|---|
| VH 1 (µm) | VW (µm) | CD (µm) | MMT (µm) | VH:CD | VSA (mm2) | ||
| CON 2 | 0 | 2753.69 f | 468.36 a | 436.71 b | 112.02 cd | 6.51 bc | 4.01 |
| CB 0.5 | 3663.17 ab | 394.18 cd | 533.53 ab | 134.44 b | 7.05 ab | 4.57 | |
| RY 0.5 | 3950.62 a | 440.426 ab | 505.99 b | 113.33 bcd | 7.91 a | 5.53 | |
| RY 1.0 | 3548.76 abc | 422.34 bc | 442.93 b | 115.83 bcd | 8.11 a | 4.71 | |
| 50% MT | 0 | 3042.28 ef | 318.40 ef | 527.51 ab | 109.74 cd | 6.03 bc | 3.05 |
| CB 0.5 | 2804.32 ef | 364.01 de | 497.33 b | 112.04 cd | 6.02 bc | 3.21 | |
| RY 0.5 | 3365.45 bcd | 331.80 ef | 520.66 ab | 118.93 bc | 6.75 ab | 3.52 | |
| RY 1.0 | 3192.44 cde | 355.04 de | 488.37 b | 113.66 bcd | 6.88 ab | 3.62 | |
| 100% MT | 0 | 2940.70 ef | 302.63 f | 611.42 a | 178.54 a | 5.33 c | 2.79 |
| CB 0.5 | 3286.89 bcd | 361.03 de | 432.99 b | 105.09 cd | 7.89 a | 3.69 | |
| RY 0.5 | 3502.98 bc | 335.90 ef | 485.68 b | 95.46 de | 7.90 a | 3.70 | |
| RY 1.0 | 3478.54 bc | 325.36 ef | 527.62 ab | 84.72 e | 6.99 ab | 3.54 | |
| SEM 3 | 26.31 | 4.15 | 9.02 | 1.98 | 0.12 | 0.06 | |
| Source of variation | p-value | ||||||
| MT | <0.001 | <0.001 | 0.248 | 0.550 | 0.005 | <0.001 | |
| Adsorbent | <0.001 | 0.863 | 0.392 | <0.001 | <0.001 | <0.001 | |
| MT × Adsorbent | <0.001 | <0.001 | 0.002 | <0.001 | 0.048 | 0.073 | |
| Ingredient (g kg−1 Unless Stated Otherwise) | Starter (1 to 21 Days) | Grower (22 to 35 Days) |
|---|---|---|
| Ground yellow corn (7.8% CP) | 47.70 | 46.00 |
| Soybean meal (44% CP) | 26.12 | 24.95 |
| Full-fat soy bean (36% CP) | 18.00 | 20.00 |
| Dicalcium phosphate | 1.70 | 1.30 |
| Salt (NaCl) | 0.30 | 0.35 |
| Vitamin Premix 1 | 0.05 | 0.05 |
| Mineral Premix 2 | 0.10 | 0.10 |
| Corn oil | 4.40 | 6.00 |
| Limestone | 1.10 | 0.80 |
| Choline | 0.05 | 0.05 |
| L-Lysine | 0.32 | 0.25 |
| DL-methionine | 0.16 | 0.15 |
| Total | 100.00 | 100.00 |
| Calculated chemical analysis | ||
| Crude protein (%) | 22.52 | 20.07 |
| Metabolisable energy (kcal kg−1) | 3121 | 3175 |
| Calcium (%) | 1.12 | 1.13 |
| Available phosphorus (%) | 0.52 | 0.53 |
| Lysine (%) | 1.42 | 1.30 |
| Methionine (%) | 0.67 | 0.56 |
| Methionine and cystine (%) | 1.06 | 0.91 |
| Tryptophan (%) | 0.29 | 0.25 |
| Threonine (%) | 0.90 | 0.80 |
| Mycotoxin Name | Assigned Value | Coefficient | ||||||
|---|---|---|---|---|---|---|---|---|
| 50 µg kg−1 (50% MT) | 100 µg kg−1 (100% MT) | |||||||
| Mean (µg kg−1) ± SD | RSD (%) | CV | Mean (µg kg−1) ± SD | RSD (%) | CV | |||
| AFB1 | 49.86 ± 0.09 | 0.18 | 0.000 | 99.08 | ± 0.09 | 0.09 | 0.000 | 0.988 |
| ZEN | 49.95 ± 2.63 | 5.27 | 0.005 | 99.66 | ± 0.85 | 0.85 | 0.001 | 0.989 |
| OTA | 49.93 ± 0.29 | 0.58 | 0.001 | 99.98 | ± 0.00 | 0.00 | 0.000 | 0.985 |
| T-2 | 49.88 ± 4.72 | 9.46 | 0.009 | 99.97 | ± 0.18 | 0.18 | 0.000 | 0.990 |
| DON | 49.96 ± 1.05 | 2.10 | 0.002 | 99.03 | ± 0.50 | 0.50 | 0.001 | 0.982 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srinual, O.; Moonmanee, T.; Lumsangkul, C.; Doan, H.V.; Punyatong, M.; Yachai, M.; Chaiyaso, T.; Kongtong, K.; Tapingkae, W. Can Red Yeast (Sporidiobolus pararoseus) Be Used as a Novel Feed Additive for Mycotoxin Binders in Broiler Chickens? Toxins 2022, 14, 678. https://doi.org/10.3390/toxins14100678
Srinual O, Moonmanee T, Lumsangkul C, Doan HV, Punyatong M, Yachai M, Chaiyaso T, Kongtong K, Tapingkae W. Can Red Yeast (Sporidiobolus pararoseus) Be Used as a Novel Feed Additive for Mycotoxin Binders in Broiler Chickens? Toxins. 2022; 14(10):678. https://doi.org/10.3390/toxins14100678
Chicago/Turabian StyleSrinual, Orranee, Tossapol Moonmanee, Chompunut Lumsangkul, Hien Van Doan, Montri Punyatong, Mongkol Yachai, Thanongsak Chaiyaso, Kittima Kongtong, and Wanaporn Tapingkae. 2022. "Can Red Yeast (Sporidiobolus pararoseus) Be Used as a Novel Feed Additive for Mycotoxin Binders in Broiler Chickens?" Toxins 14, no. 10: 678. https://doi.org/10.3390/toxins14100678
APA StyleSrinual, O., Moonmanee, T., Lumsangkul, C., Doan, H. V., Punyatong, M., Yachai, M., Chaiyaso, T., Kongtong, K., & Tapingkae, W. (2022). Can Red Yeast (Sporidiobolus pararoseus) Be Used as a Novel Feed Additive for Mycotoxin Binders in Broiler Chickens? Toxins, 14(10), 678. https://doi.org/10.3390/toxins14100678