Collagen-Derived Peptides in CKD: A Link to Fibrosis

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Patient Characteristics
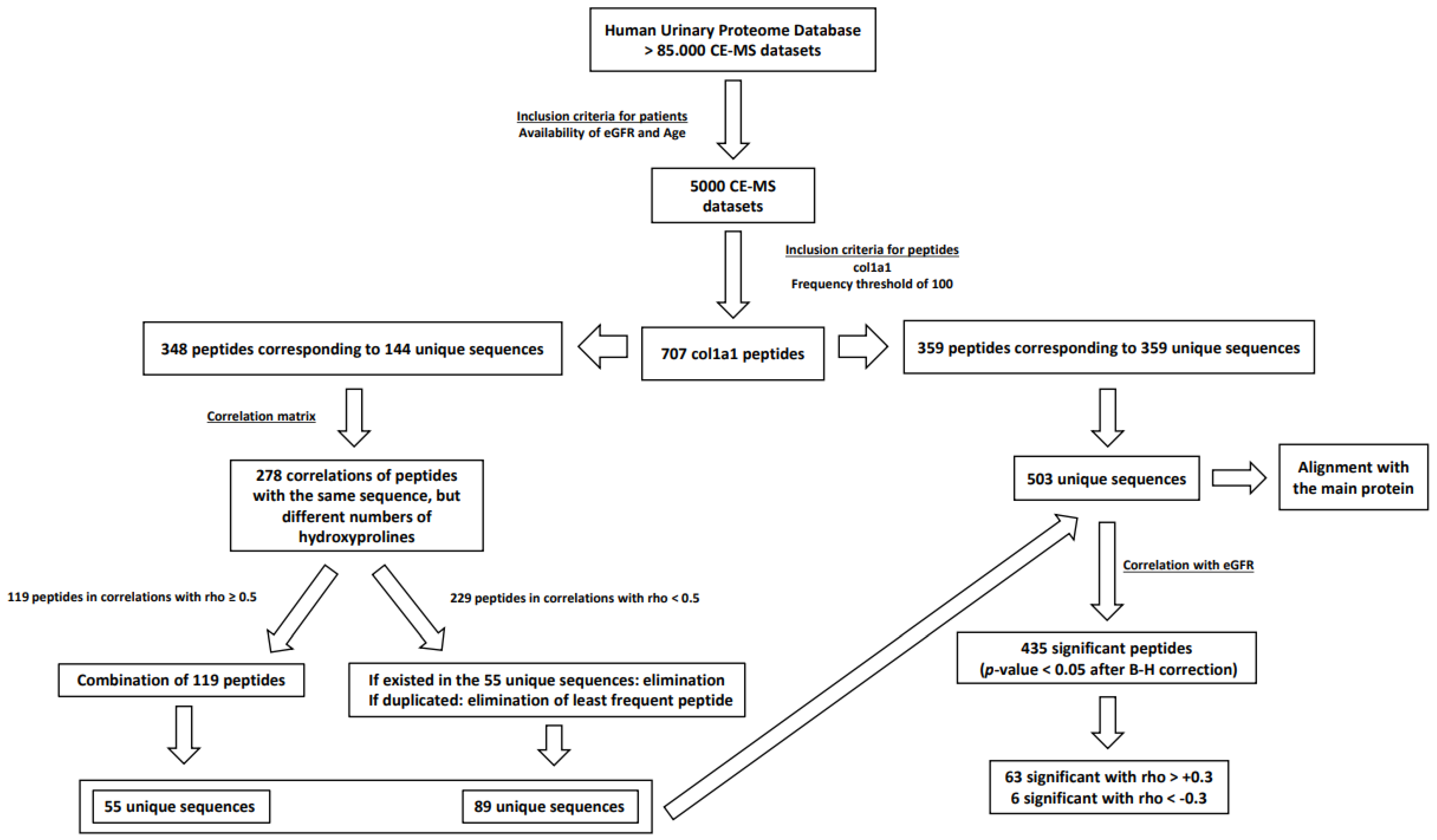
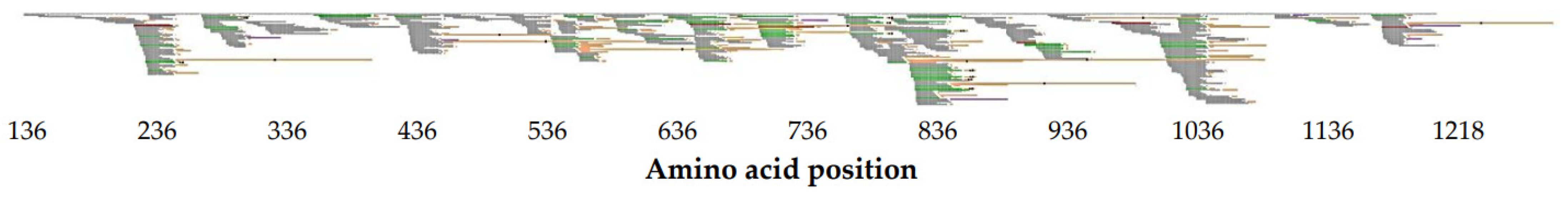
2.2. Identification of Unique col1a1 Protein Fragments
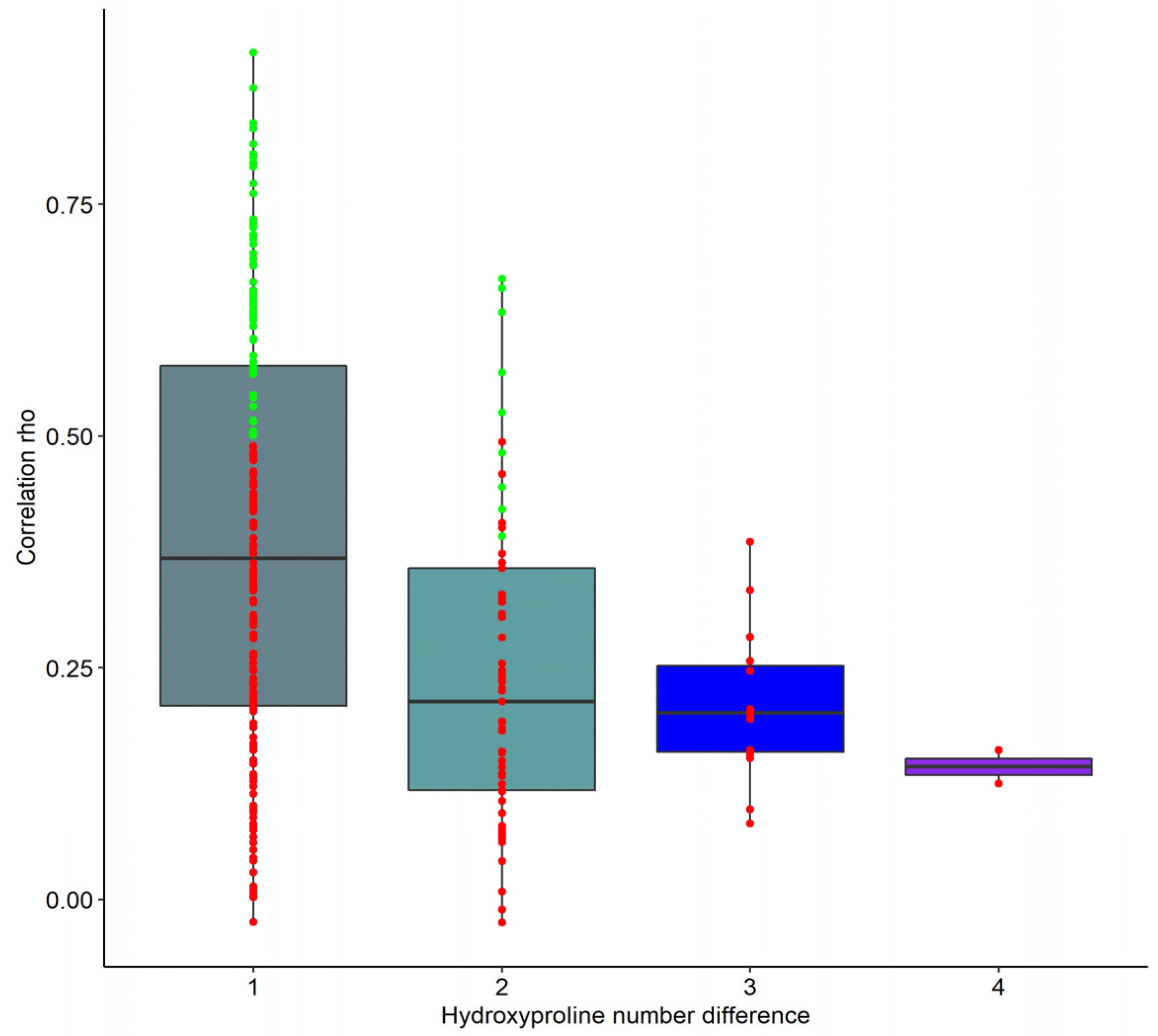
2.3. Correlation of Unique col1a1 Protein Fragments with eGFR and Age
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patients
5.2. Data Curation
5.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Huizen, N.A.; Ijzermans, J.N.M.; Burgers, P.C.; Luider, T.M. Collagen analysis with mass spectrometry. Mass Spectrom. Rev. 2020, 39, 309–335. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Friess, W. Collagen—Biomaterial for drug delivery. Eur. J. Pharm. Biopharm. 1998, 45, 113–136. [Google Scholar] [CrossRef]
- Di Lullo, G.A.; Sweeney, S.M.; Körkkö, J.; Ala-Kokko, L.; Antonio, J.D.S. Mapping the ligand-binding sites and disease-associated mutations on the most abundant protein in the human, type I collagen. J. Biol. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kwan, J.Y.Y.; Yip, K.; Liu, P.P.; Liu, F.-F. Targeting metabolic dysregulation for fibrosis therapy. Nat. Rev. Drug Discov. 2019, 19, 57–75. [Google Scholar] [CrossRef] [PubMed]
- Distler, J.H.W.; Györfi, A.-H.; Ramanujam, M.; Whitfield, M.L.; Königshoff, M.; Lafyatis, R. Shared and distinct mechanisms of fibrosis. Nat. Rev. Rheumatol. 2019, 15, 705–730. [Google Scholar] [CrossRef] [PubMed]
- Bosman, F.T.; Stamenkovic, I. Functional structure and composition of the extracellular matrix. J. Pathol. 2003, 200, 423–428. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, P.; Pejchinovski, M.; Markoska, K.; Banasik, M.; Klinger, M.; Švec-Billá, D.; Rychlík, I.; Rroji, M.; Restivo, A.; Capasso, G.; et al. Association of kidney fibrosis with urinary peptides: A path towards non-invasive liquid biopsies? Sci. Rep. 2017, 7, 16915. [Google Scholar] [CrossRef]
- He, T.; Melgarejo, J.D.; Clark, A.L.; Yu, Y.; Thijs, L.; Díez, J.; López, B.; González, A.; Cleland, J.G.; Schanstra, J.P.; et al. Serum and urinary biomarkers of collagen type-I turnover predict prognosis in patients with heart failure. Clin. Transl. Med. 2021, 11, 2–5. [Google Scholar] [CrossRef]
- Wendt, R.; He, T.; Latosinska, A.; Siwy, J.; Mischak, H.; Beige, J. Proteomic characterization of obesity-related nephropathy. Clin. Kidney J. 2020, 13, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Schanstra, J.P.; Zürbig, P.; Alkhalaf, A.; Argiles, A.; Bakker, S.J.L.; Beige, J.; Bilo, H.J.G.; Chatzikyrkou, C.; Dakna, M.; Dawson, J.; et al. Diagnosis and prediction of CKD progression by assessment of urinary peptides. J. Am. Soc. Nephrol. 2015, 26, 1999–2010. [Google Scholar] [CrossRef]
- Mavrogeorgis, E.; Mischak, H.; Beige, J.; Latosinska, A.; Siwy, J. Understanding glomerular diseases through proteomics. Expert Rev. Proteom. 2021, 18, 137–157. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Bascands, J.L.; Mischak, H.; Schanstra, J.P. The role of urinary peptidomics in kidney disease research. Kidney Int. 2016, 89, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Latosinska, A.; Frantzi, M.; Vlahou, A.; Mischak, H. Clinical applications of capillary electrophoresis coupled to mass spectrometry in biomarker discovery: Focus on bladder cancer. Proteom. Clin. Appl. 2013, 7, 779–793. [Google Scholar] [CrossRef]
- Latosinska, A.; Siwy, J.; Mischak, H.; Frantzi, M. Peptidomics and proteomics based on CE-MS as a robust tool in clinical application: The past, the present, and the future. Electrophoresis 2019, 40, 2294–2308. [Google Scholar] [CrossRef]
- Klein, J.; Papadopoulos, T.; Mischak, H.; Mullen, W. Comparison of CE-MS/MS and LC-MS/MS sequencing demonstrates significant complementarity in natural peptide identification in human urine. Electrophoresis 2014, 35, 1060–1064. [Google Scholar] [CrossRef]
- Mischak, H.; Vlahou, A.; Ioannidis, J.P.A. Technical aspects and inter-laboratory variability in native peptide profiling: The CE-MS experience. Clin. Biochem. 2013, 46, 432–443. [Google Scholar] [CrossRef]
- Martens, D.S.; Thijs, L.; Latosinska, A.; Trenson, S.; Siwy, J.; Zhang, Z.-Y.; Wang, C.; Beige, J.; Vlahou, A.; Janssens, S.; et al. Urinary peptidomic profiles to address age-related disabilities: A prospective population study. Lancet Health Longev. 2021, 2, e690–e703. [Google Scholar] [CrossRef]
- Nkuipou-Kenfack, E.; Bhat, A.; Klein, J.; Jankowski, V.; Mullen, W.; Vlahou, A.; Dakna, M.; Koeck, T.; Schanstra, J.P.; Zürbig, P.; et al. Identification of ageing-associated naturally occurring peptides in human urine. Oncotarget 2015, 6, 34106–34117. [Google Scholar] [CrossRef]
- Zürbig, P.; Decramer, S.; Dakna, M.; Jantos, J.; Good, D.M.; Coon, J.J.; Bandin, F.; Mischak, H.; Bascands, J.-L.; Schanstra, J.P. The human urinary proteome reveals high similarity between kidney aging and chronic kidney disease. Proteomics 2009, 9, 2108–2117. [Google Scholar] [CrossRef]
- Latosinska, A.; Siwy, J.; Faguer, S.; Beige, J.; Mischak, H.; Schanstra, J.P. Value of Urine Peptides in Assessing Kidney and Cardiovascular Disease. Proteom. Clin. Appl. 2020, 15, 2000027. [Google Scholar] [CrossRef] [PubMed]
- McKleroy, W.; Lee, T.-H.; Atabai, K. Always cleave up your mess: Targeting collagen degradation to treat tissue fibrosis. Am. J. Physiol. Cell. Mol. Physiol. 2013, 304, L709–L721. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Sakakibara, S.; Inouye, K.; Shudo, K.; Kishida, Y.; Kobayashi, Y.; Prockop, D.J. Synthesis of (Pro-Hyp-Gly)n of defined molecular weights Evidence for the stabilization of collagen triple helix by hydroxypyroline. Biochim. Biophys. Acta Protein Struct. 1973, 303, 198–202. [Google Scholar] [CrossRef]
- Berg, R.A.; Prockop, D.J. The thermal transition of a non-hydroxylated form of collagen. Evidence for a role for hydroxyproline in stabilizing the triple-helix of collagen. Biochem. Biophys. Res. Commun. 1973, 52, 115–120. [Google Scholar] [CrossRef]
- Jiravanichanun, N.; Nishino, N.; Okuyama, K. Conformation of alloHyp in the Y position in the host–guest peptide with the pro–pro–gly sequence: Implication of the destabilization of (Pro–alloHyp–Gly)10. Biopolymers 2006, 81, 225–233. [Google Scholar] [CrossRef]
- Inouy, K.; Sakakibara, S.; Prockop, D.J. Effects of the stereo-configuration of the hydroxyl group in 4-hydroxyproline on the triple-helical structures formed by homogeneous peptides resembling collagen. Biochim. Biophys. Acta Protein Struct. 1976, 420, 133–141. [Google Scholar] [CrossRef]
- He, T.; Pejchinovski, M.; Mullen, W.; Beige, J.; Mischak, H.; Jankowski, V. Peptides in Plasma, Urine, and Dialysate: Toward Unravelling Renal Peptide Handling. Proteom. Clin. Appl. 2020, 15, e2000029. [Google Scholar] [CrossRef]
- Gkogkolou, P.; Böhm, M. Advanced glycation end products: Keyplayers in skin aging? Derm. Endocrinol. 2012, 4, 259–270. [Google Scholar] [CrossRef]
- Ahmed, N. Advanced glycation endproducts—Role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef]
- Verzijl, N.; DeGroot, J.; Thorpe, S.R.; Bank, R.A.; Shaw, J.N.; Lyons, T.J.; Bijlsma, J.W.J.; Lafeber, F.P.J.G.; Baynes, J.W.; TeKoppele, J.M. Effect of collagen turnover on the accumulation of advanced glycation end products. J. Biol. Chem. 2000, 275, 39027–39031. [Google Scholar] [CrossRef] [PubMed]
- Avery, N.C.; Bailey, A.J. The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathol. Biol. 2006, 54, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Haitoglou, C.S.; Tsilibary, E.C.; Brownlee, M.; Charonis, A.S. Altered cellular interactions between endothelial cells and nonenzymatically glucosylated laminin/type IV collagen. J. Biol. Chem. 1992, 267, 12404–12407. [Google Scholar] [CrossRef]
- Fessel, G.; Li, Y.; Diederich, V.; Guizar-Sicairos, M.; Schneider, P.; Sell, D.R.; Monnier, V.M.; Snedeker, J.G. Advanced Glycation End-Products Reduce Collagen Molecular Sliding to Affect Collagen Fibril Damage Mechanisms but Not Stiffness. PLoS ONE 2014, 9, e110948. [Google Scholar] [CrossRef]
- Kuzuya, M.; Asai, T.; Kanda, S.; Maeda, K.; Cheng, X.W.; Iguchi, A. Glycation cross-links inhibit matrix metalloproteinase-2 activation in vascular smooth muscle cells cultured on collagen lattice. Diabetologia 2001, 44, 433–436. [Google Scholar] [CrossRef][Green Version]
- McLennan, S.V.; Kelly, D.J.; Schache, M.; Waltham, M.; Dy, V.; Langham, R.G.; Yue, D.K.; Gilbert, R.E. Advanced glycation end products decrease mesangial cell MMP-7: A role in matrix accumulation in diabetic nephropathy? Kidney Int. 2007, 72, 481–488. [Google Scholar] [CrossRef]
- Degroot, J.; Verzijl, N.; Wenting-Van Wijk, M.J.G.; Bank, R.A.; Lafeber, F.P.J.G.; Bijlsma, J.W.J.; Tekoppele, J.M. Age-Related Decrease in Susceptibility of Human Articular Cartilage to Matrix Metalloproteinase-Mediated Degradation The Role of Advanced Glycation End Products. Arthritis Rheum. 2001, 44, 2562–2571. [Google Scholar] [CrossRef]
- Robins, S.P. Biochemistry and functional significance of collagen cross-linking. Biochem. Soc. Trans. 2007, 35, 849–852. [Google Scholar] [CrossRef]
- McKay, T.B.; Priyadarsini, S.; Karamichos, D. Mechanisms of Collagen Crosslinking in Diabetes and Keratoconus. Cells 2019, 8, 1239. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zhang, Z.; Staessen, J.A.; Mischak, H.; Latosinska, A.; Beige, J. Proteomic Biomarkers in the Cardiorenal Syndrome: Toward Deciphering Molecular Pathophysiology. Am. J. Hypertens. 2021, 34, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Catanese, L.; Siwy, J.; Mavrogeorgis, E.; Amann, K.; Mischak, H.; Beige, J.; Rupprecht, H. A novel urinary proteomics classifier for non-invasive evaluation of interstitial fibrosis and tubular atrophy in chronic kidney disease. Proteomes 2021, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Good, D.M.; Zürbig, P.; Argilés, À.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol. Cell. Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef] [PubMed]
- Wendt, R.; Thijs, L.; Kalbitz, S.; Mischak, H.; Siwy, J.; Raad, J.; Metzger, J.; Neuhaus, B.; von der Leyen, H.; Dudoignon, E.; et al. A urinary peptidomic profile predicts outcome in SARS-CoV-2-infected patients. EClinicalMedicine 2021, 36, 100883. [Google Scholar] [CrossRef]
- Jantos-Siwy, J.; Schiffer, E.; Brand, K.; Brand, K.; Schumann, G.; Schumann, G.; Rossing, K.; Delles, C.; Mischak, H.; Metzger, J. Quantitative urinary proteome analysis for biomarker evaluation in chronic kidney disease. J. Proteome Res. 2009, 8, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Mavrogeorgis, E.; Mischak, H.; Latosinska, A.; Siwy, J.; Jankowski, V.; Jankowski, J. Reproducibility Evaluation of Urinary Peptide Detection Using CE-MS. Molecules 2021, 26, 7260. [Google Scholar] [CrossRef] [PubMed]
- Siwy, J.; Schanstra, J.P.; Argiles, A.; Bakker, S.J.L.; Beige, J.; Boucek, P.; Brand, K.; Delles, C.; Duranton, F.; Fernandez-Fernandez, B.; et al. Multicentre prospective validation of a urinary peptidome-based classifier for the diagnosis of type 2 diabetic nephropathy. Nephrol. Dial. Transplant. 2014, 29, 1563–1570. [Google Scholar] [CrossRef]
- Pontillo, C.; Zhang, Z.Y.; Schanstra, J.P.; Jacobs, L.; Zürbig, P.; Thijs, L.; Ramírez-Torres, A.; Heerspink, H.J.L.; Lindhardt, M.; Klein, R.; et al. Prediction of Chronic Kidney Disease Stage 3 by CKD273, a Urinary Proteomic Biomarker. Kidney Int. Rep. 2017, 2, 1066–1075. [Google Scholar] [CrossRef]
- Rudnicki, M.; Siwy, J.; Wendt, R.; Lipphardt, M.; Koziolek, M.J.; Maixnerova, D.; Peters, B.; Kerschbaum, J.; Leierer, J.; Neprasova, M.; et al. Urine proteomics for prediction of disease progression in patients with IgA nephropathy. Nephrol. Dial. Transplant. 2020, 35, gfaa140-MO041. [Google Scholar] [CrossRef]
- Siwy, J.; Zürbig, P.; Argiles, A.; Beige, J.; Haubitz, M.; Jankowski, J.; Julian, B.A.; Linde, P.G.; Marx, D.; Mischak, H.; et al. Noninvasive diagnosis of chronic kidney diseases using urinary proteome analysis. Nephrol. Dial. Transplant. 2016, 32, 2079–2089. [Google Scholar] [CrossRef]
- He, T.; Mischak, M.; Clark, A.L.; Campbell, R.T.; Delles, C.; Díez, J.; Filippatos, G.; Mebazaa, A.; McMurray, J.J.V.; González, A.; et al. Urinary peptides in heart failure: A link to molecular pathophysiology. Eur. J. Heart Fail. 2021, 2, 1875–1887. [Google Scholar] [CrossRef] [PubMed]
- Rickham, P.P. Human experimentation. Code of ethics of the world medical association. Declaration of Helsinki. Br. Med. J. 1964, 2, 177. [Google Scholar] [CrossRef]
- Levey, A.S.; Stevens, L.A.; Schmid, C.H.; Zhang, Y.; Castro, A.F., III; Feldman, H.I.; Kusek, J.W.; Eggers, P.; Lente, F.V.; Greene, T.; et al. A New Equation to Estimate Glomerular Filtration Rate. Ann. Intern. Med. 2009, 150, 604. [Google Scholar] [CrossRef]
- Xiao, N.; Cao, D.-S.; Zhu, M.-F.; Xu, Q.-S. protr/ProtrWeb: R package and web server for generating various numerical representation schemes of protein sequences. Bioinformatics 2015, 31, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.; Imai, K.; King, G.; Stuart, E.A. MatchIt: Nonparametric Preprocessing for Parametric Causal Inference. J. Stat. Softw. 2011, 42, 1–28. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Disease Aetiology | Number |
|---|---|
| Controls | 1717 |
| Amyloidosis | 3 |
| Diabetes mellitus | 2756 |
| Focal segmental glomerulosclerosis | 27 |
| IgA nephropathy | 247 |
| Minimal change disease | 16 |
| Membranous glomerulopathy | 28 |
| Membranoproliferative glomerulopathy | 2 |
| Nephritis | 3 |
| Nephrosclerosis | 135 |
| Systemic lupus erythematosus | 22 |
| Steroid-Resistant Nephrotic Syndrome | 4 |
| Vasculitis | 40 |
| Sequence | Start AA | Stop AA | rho eGFR | eGFR p-Value (B-H) | rho Age | Age p-Value (B-H) |
|---|---|---|---|---|---|---|
| ADGQpGAKGEpGDAGAKGDAGpPGPAGPAGPpGPIG | 819 | 854 | 0.61 | 0.00 | −0.39 | 1.14 × 10−180 |
| IGPpGPAGApGDKGESGPSGPAGPTG | 769 | 794 | 0.59 | 0.00 | −0.38 | 7.36 × 10−172 |
| LTGPIGppGPAGAPGDKGESGPSGPAGPTG | 765 | 794 | 0.57 | 0.00 | −0.36 | 1.19 × 10−153 |
| pPGADGQPGAKGEpGDAGAKGDAGppGPAGPAGPPGPIG | 816 | 854 | 0.55 | 0.00 | −0.34 | 2.69 × 10−132 |
| PpGPAGFAGPPGADGQPGAKGEpGDAGAKGDAGPPGPAGP | 807 | 846 | 0.54 | 0.00 | −0.31 | 7.21 × 10−110 |
| LDGAKGDAGPAGPKGEpGSpGENGApG | 273 | 299 | 0.50 | 0.00 | −0.38 | 5.05 × 10−169 |
| TGPIGpPGPAGAPGDKGESGpSGPAGPTG | 766 | 794 | 0.50 | 0.00 | −0.29 | 3.64 × 10−94 |
| GPpGADGQPGAKGEpGDAGAKGDAGPPGpAGPAGPPGpIG | 815 | 854 | 0.50 | 1.02 × 10−305 | −0.35 | 3.06 × 10−142 |
| GADGQpGAKGEpGDAGAKGDAGPPGPAGPAGPpGPIG | 818 | 854 | 0.48 | 1.42 × 10−291 | −0.27 | 6.08 × 10−81 |
| NGDDGEAGKPGRpGERGPpGPQG | 229 | 251 | 0.48 | 3.41 × 10−279 | −0.25 | 3.26 × 10−69 |
| Sequence | Start AA | Stop AA | rho Age-corrected eGFR | Age-corrected eGFR p-value (B-H) | rho Age-matched | Age-matched p-value (B-H) |
| ADGQpGAKGEpGDAGAKGDAGpPGPAGPAGPpGPIG | 819 | 854 | 0.44 | 2.84 × 10−232 | −0.27 | 4.28 × 10−16 |
| PpGPAGFAGPPGADGQPGAKGEpGDAGAKGDAGPPGPAGP | 807 | 846 | 0.41 | 1.64 × 10−202 | −0.15 | 1.92 × 10−05 |
| IGPpGPAGApGDKGESGPSGPAGPTG | 769 | 794 | 0.41 | 1.05 × 10−201 | −0.28 | 4.70 × 10−17 |
| PGPAGPPGEAGKPGEQGVPGDLGAPGPSGARG | 646 | 677 | −0.41 | 9.43 × 10−197 | −0.06 | 1.47 × 10−01 |
| LTGPIGppGPAGAPGDKGESGPSGPAGPTG | 765 | 794 | 0.40 | 1.24 × 10−191 | −0.27 | 5.95 × 10−16 |
| pPGADGQPGAKGEpGDAGAKGDAGppGPAGPAGPPGPIG | 816 | 854 | 0.40 | 4.80 × 10−186 | −0.29 | 6.76 × 10−19 |
| TGPIGpPGPAGAPGDKGESGpSGPAGPTG | 766 | 794 | 0.38 | 1.26 × 10−168 | −0.24 | 1.22 × 10−12 |
| NGDDGEAGKPGRpGERGPpGPQG | 229 | 251 | 0.37 | 4.25 × 10−163 | −0.22 | 1.15 × 10−10 |
| KEGGKGPRGETGPAGRpGEVGPpGPpGPAG | 903 | 932 | 0.37 | 8.82 × 10−160 | −0.10 | 6.96 × 10−03 |
| GADGQpGAKGEpGDAGAKGDAGPPGPAGPAGPpGPIG | 818 | 854 | 0.37 | 1.68 × 10−158 | −0.21 | 6.90 × 10−10 |
| Sequence | Start AA | Stop AA | rho Age-corrected eGFR | Age-corrected eGFR p-value (B-H) | rho Age-matched | Age-matched p-value (B-H) |
| DAGPAGPKGEpGSpGENGApG | 279 | 299 | 0.21 | 2.87 × 10−52 | −0.39 | 2.15 × 10−33 |
| LDGAKGDAGPAGPKGEpGSpGENGApG | 273 | 299 | 0.31 | 9.95 × 10−111 | −0.36 | 5.16 × 10−29 |
| pGpAGEKGSpGADGPAGAP | 928 | 946 | 0.02 | 2.58 × 10−01 | 0.36 | 1.10 × 10−28 |
| GLPGpAGppGEAGKPGEQGVPGDLGApGP | 644 | 672 | 0.16 | 2.92 × 10−30 | −0.36 | 1.68 × 10−28 |
| ADGQpGAKGEpGDAGAKGDAGPPGPAGP | 819 | 846 | 0.30 | 1.83 × 10−101 | −0.35 | 5.81 × 10−27 |
| GSpGSpGPDGKTGPpGPAG | 542 | 560 | 0.21 | 4.83 × 10−48 | −0.35 | 2.43 × 10−26 |
| EpGSpGENGAPGQmGPR | 288 | 304 | 0.09 | 3.28 × 10−11 | 0.32 | 3.01 × 10−22 |
| pPGADGQPGAKGEpGDAGAKGDAGppGPAGPAGPPGPIG | 816 | 854 | 0.40 | 4.80 × 10−186 | −0.29 | 6.76 × 10−19 |
| EGSPGRDGSPGAK | 1021 | 1033 | 0.15 | 4.99 × 10−26 | 0.28 | 2.41 × 10−17 |
| IGPpGPAGApGDKGESGPSGPAGPTG | 769 | 794 | 0.41 | 1.05 × 10−201 | −0.28 | 4.70 × 10−17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavrogeorgis, E.; Mischak, H.; Latosinska, A.; Vlahou, A.; Schanstra, J.P.; Siwy, J.; Jankowski, V.; Beige, J.; Jankowski, J. Collagen-Derived Peptides in CKD: A Link to Fibrosis. Toxins 2022, 14, 10. https://doi.org/10.3390/toxins14010010
Mavrogeorgis E, Mischak H, Latosinska A, Vlahou A, Schanstra JP, Siwy J, Jankowski V, Beige J, Jankowski J. Collagen-Derived Peptides in CKD: A Link to Fibrosis. Toxins. 2022; 14(1):10. https://doi.org/10.3390/toxins14010010
Chicago/Turabian StyleMavrogeorgis, Emmanouil, Harald Mischak, Agnieszka Latosinska, Antonia Vlahou, Joost P. Schanstra, Justyna Siwy, Vera Jankowski, Joachim Beige, and Joachim Jankowski. 2022. "Collagen-Derived Peptides in CKD: A Link to Fibrosis" Toxins 14, no. 1: 10. https://doi.org/10.3390/toxins14010010
APA StyleMavrogeorgis, E., Mischak, H., Latosinska, A., Vlahou, A., Schanstra, J. P., Siwy, J., Jankowski, V., Beige, J., & Jankowski, J. (2022). Collagen-Derived Peptides in CKD: A Link to Fibrosis. Toxins, 14(1), 10. https://doi.org/10.3390/toxins14010010