Mycotoxin Biomarkers in Pigs—Current State of Knowledge and Analytics



Abstract
:1. Introduction
2. Impact of Mycotoxins on Pig Health
2.1. Aflatoxins (AFs)
2.2. Deoxynivalenol (DON)
2.3. Fumonisin B1 (FB1) and Fumonisin B2 (FB2)
2.4. Ochratoxin A (OTA)
2.5. T-2 Toxin (T-2)
2.6. Zearalenone (ZEN)
2.7. Modified Mycotoxins
3. In Vivo Mycotoxin Absorption, Distribution, Metabolism and Excretion (ADME) and Toxicokinetic Studies on Pigs
3.1. ZEN Administration
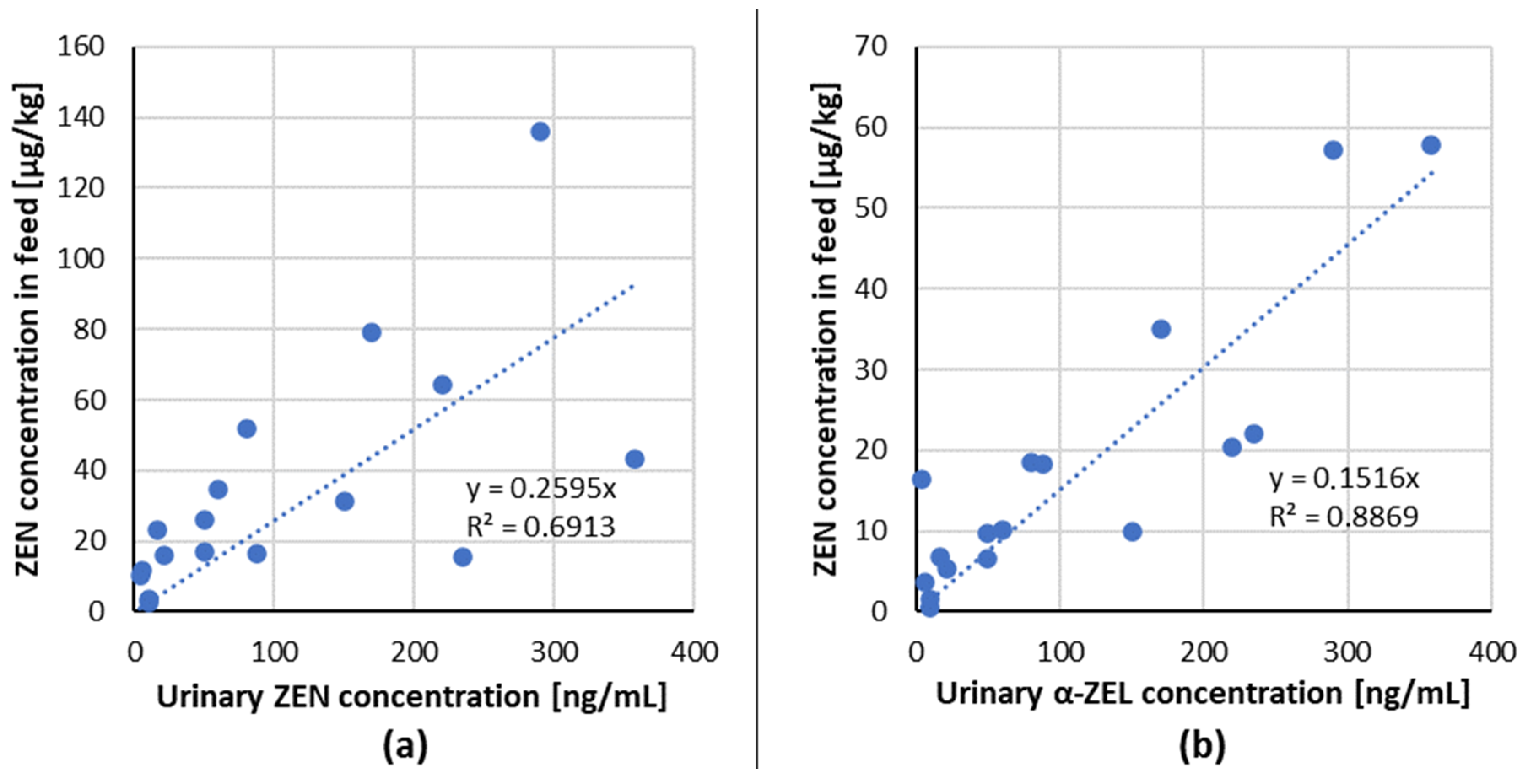
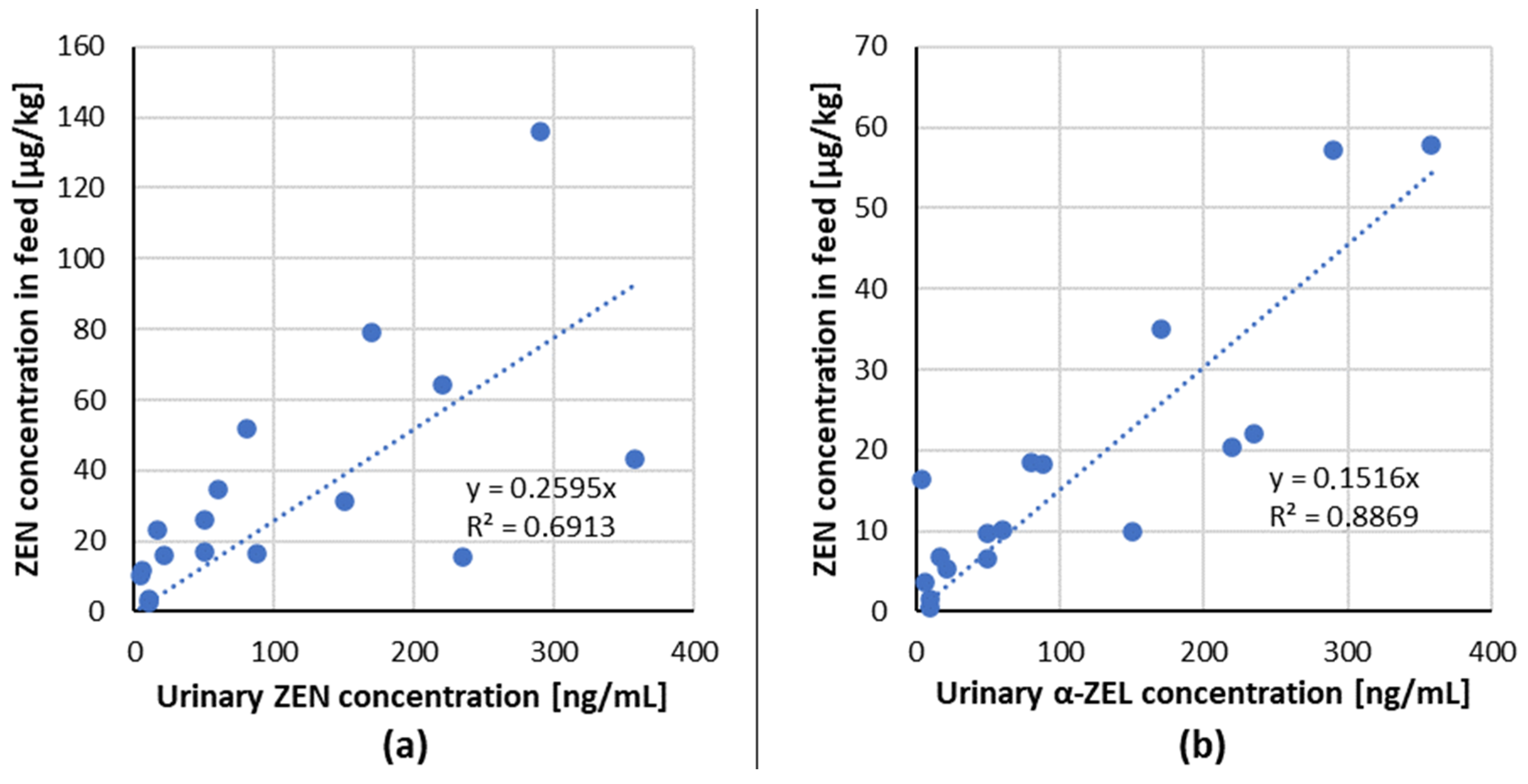
3.1.1. ZEN Urinary Biomarkers
3.1.2. Faecal Biomarkers
3.1.3. Serum Biomarkers
3.1.4. Administration of ZEN and Its Modified Forms
3.2. Deoxynivalenol Administration
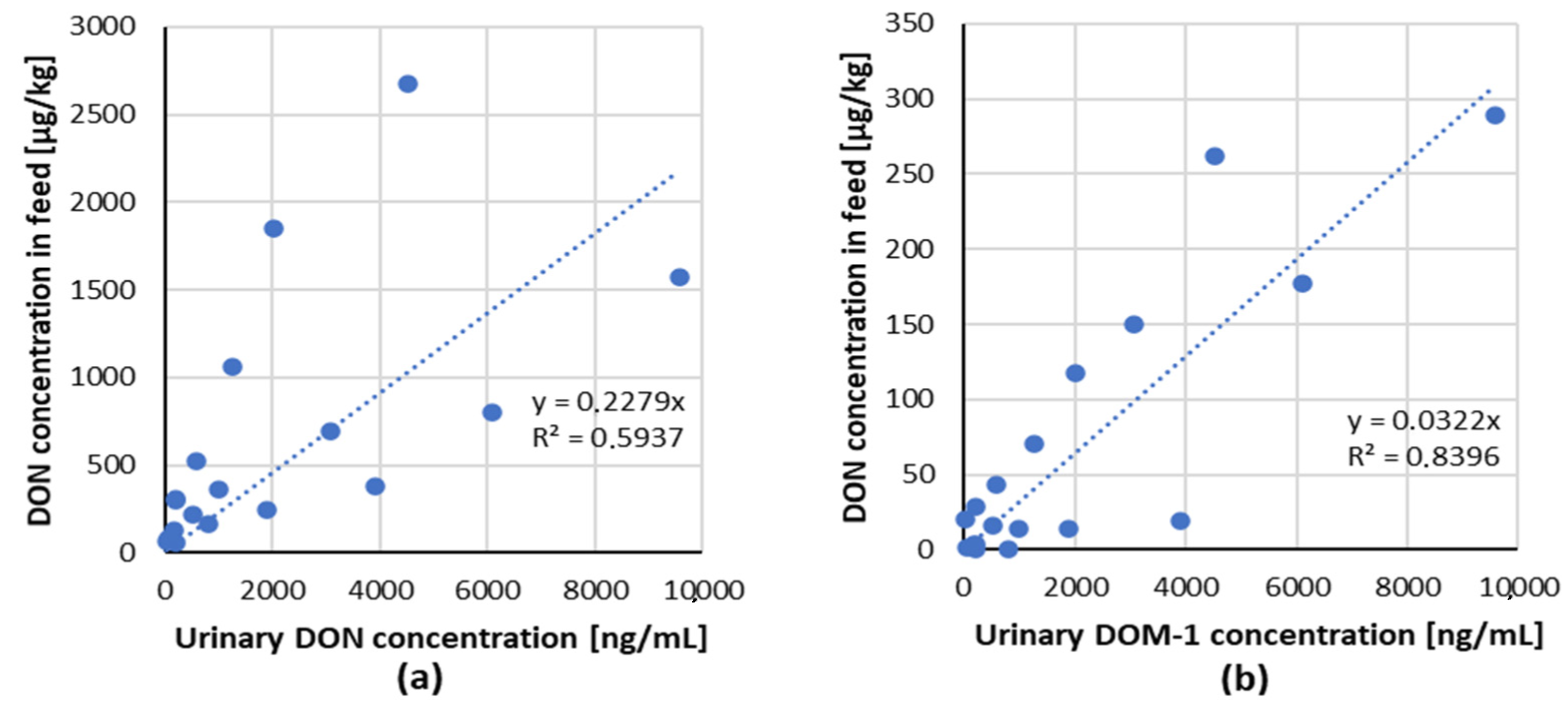
3.2.1. DON Urinary Biomarkers
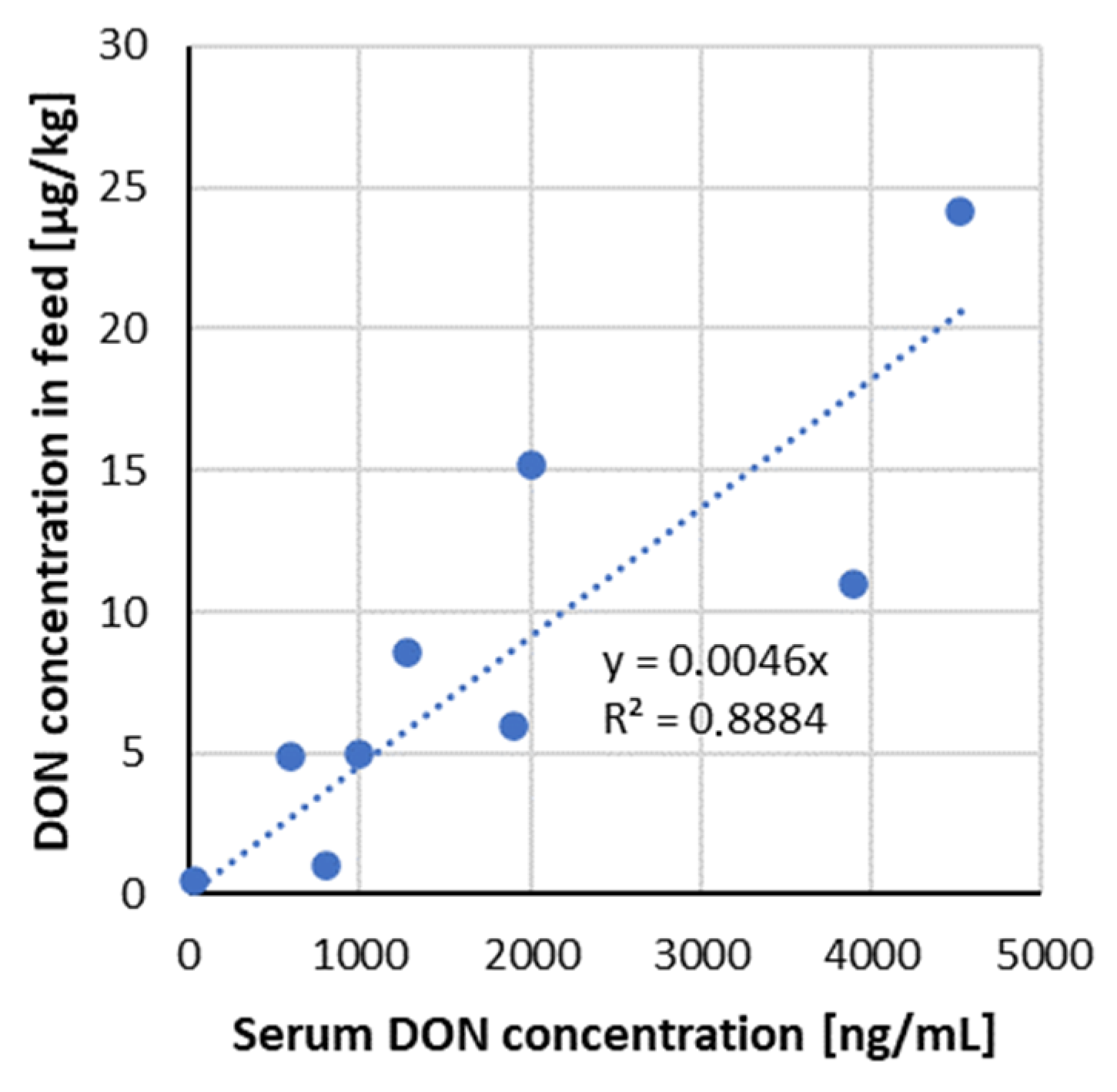
3.2.2. DON Serum Biomarkers
3.2.3. DON Faecal Biomarkers
3.3. Administration of DON and its Modified Forms
3.4. Other Mycotoxins
3.4.1. AFB1 Administration
3.4.2. OTA Administration
3.4.3. FB1 Administration
3.4.4. T-2 Administration
3.5. The Most Suitable Biomarkers Found in Different Biological Matrices
4. Methods for Mycotoxin Biomarkers Analysis
4.1. Urine Sample Preparation
4.2. Faeces
4.3. Serum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neme, K.; Mohammed, A. Mycotoxin occurrence in grains and the role of postharvest management as a mitigation strategies. A review. Food Control 2017, 78, 412–425. [Google Scholar] [CrossRef]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin contamination in the EU feed supply chain: A focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives. Report on Meeting of Experts Committeed and Study Groups, Evalualtion of Certain Mycotoxins; Excecutive Board, EB148/47; 23 December 2020. [Google Scholar]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Guerre, P. Worldwide Mycotoxins Exposure in Pig and Poultry Feed Formulations. Toxins 2016, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Heyndrickx, E.; Sioen, I.; Huybrechts, B.; Callebaut, A.; De Henauw, S.; De Saeger, S. Human biomonitoring of multiple mycotoxins in the Belgian population: Results of the BIOMYCO study. Environ. Int. 2015, 84, 82–89. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific Opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916. [Google Scholar] [CrossRef] [Green Version]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, A.R. Masked mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize–An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boevre, M.; Jacxsens, L.; Lachat, C.; Eeckhout, M.; Diana, J.; Mavungu, D.; Audenaert, K.; Maene, P.; Haesaert, G.; Kolsteren, P.; et al. Human exposure to mycotoxins and their masked forms through cereal-based foods in Belgium. Toxicol. Lett. 2013, 218, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekaert, N.; Devreese, M.; De Mil, T.; Fraeyman, S.; Antonissen, G.; De Baere, S.; De Backer, P.; Vermeulen, A.; Croubels, S. Oral Bioavailability, Hydrolysis, and Comparative Toxicokinetics of 3-Acetyldeoxynivalenol and 15-Acetyldeoxynivalenol in Broiler Chickens and Pigs. J. Agric. Food Chem. 2015, 63, 8734–8742. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachová, A.; et al. In Vivo contribution of deoxynivalenol-3-β-d-glucoside to deoxynivalenol exposure in broiler chickens and pigs: Oral bioavailability, hydrolysis and toxicokinetics. Arch. Toxicol. 2017, 91, 699–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iheshiulor, O.O.M.; Esonu, B.O.; Chuwuka, O.K.; Omede, A.A.; Okoli, I.C.; Ogbuewu, I.P. Effects of Mycotoxins in Animal Nutrition: A Review. Asian J. Anim. Sci. 2011, 5, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [Green Version]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Contaminants in the Food Chain. Opinion of the Scientific Panel on Contaminants in the Food Chain on a Request from the Commission Related To Ochratoxin A in Food. EFSA J. 2006, 4, 365. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Opinion of the Scientific Panel on Contaminants in Food Chain on a Request from the Commission related to Fumonisins as Undesirable Substances in Animal Feed. EFSA J. 2005, 235, 1–32. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2011, 9, 2481. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—The IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Bunaciu, R.P.; Pascale, F.; Tudor, D.S.; Avram, N.; Sarca, M.; Cureu, I.; Criste, R.D.; Suta, V.; et al. Changes in performance, blood parameters, humoral and cellular immune responses in weanling piglets exposed to low doses of aflatoxin. J. Anim. Sci. 2002, 80, 1250–1257. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Peles, F.; Sipos, P.; Győri, Z.; Pfliegler, W.P.; Giacometti, F.; Serraino, A.; Pagliuca, G.; Gazzotti, T.; Pócsi, I. Adverse Effects, Transformation and Channeling of Aflatoxins Into Food Raw Materials in Livestock. Front. Microbiol. 2019, 10, 2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiemann, U.; Dänicke, S. In Vivo and In Vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs: A review. Food Addit. Contam. 2007, 24, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.W.; Constable, P.D.; Tumbleson, M.E.; Rottinghaus, G.E.; Haschek, W.M. Sequence of cardiovascular changes leading to pulmonary edema in swine fed culture material containing fumonisin. Am. J. Vet. Res. 1999, 60, 1292–1300. [Google Scholar]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect. 2001, 109 (Suppl. 2), 251–257. [Google Scholar]
- Theumer, M.G.; López, A.G.; Masih, D.T.; Chulze, S.N.; Rubinstein, H.R. Immunobiological effects of fumonisin B1 in experimental subchronic mycotoxicoses in rats. Clin. Diagn. Lab. Immunol. 2002, 9, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Zomborszky-Kovács, M.; Kovács, F.; Horn, P.; Vetési, F.; Repa, I.; Tornyos, G.; Tóth, Á. Investigations into the time- and dose-dependent effect of fumonisin B1 in order to determine tolerable limit values in pigs. Livest. Prod. Sci. 2002, 76, 251–256. [Google Scholar] [CrossRef]
- Zomborszky-Kovács, M.; Vetési, F.; Horn, P.; Repa, I.; Kovács, F. Effects of prolonged exposure to low-dose fumonisin B1 in pigs. J. Vet. Med. Ser. B 2002, 49, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at doses below EU regulatory limits induce histological alterations in piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef] [Green Version]
- Abrunhosa, L.; Paterson, R.R.M.; Venâncio, A. Biodegradation of ochratoxin a for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [Green Version]
- Stoev, S.D.; Gundasheva, D.; Zarkov, I.; Mircheva, T.; Zapryanova, D.; Denev, S.; Mitev, Y.; Daskalov, H.; Dutton, M.; Mwanza, M.; et al. Experimental mycotoxic nephropathy in pigs provoked by a mouldy diet containing ochratoxin A and fumonisin B1. Exp. Toxicol. Pathol. 2012, 64, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P.; Gyrd-Hansen, N.; Hatd, B.; Larsen, S.; Nielsen, J.P.; Smith, M.; Ivanoff, C.; Meisner, H. Renal enzyme activities in experimental ochratoxin a-induced porcine nephropathy: Diagnostic potential of phosphoenolpyruvate carboxyk1nase and gamma-glutamyl transpeptidase activity. J. Toxicol. Environ. Health 1988, 23, 1–14. [Google Scholar] [CrossRef]
- Pietruszka, K.; Piątkowska, M.; Jedziniak, P. Occurrence of ochratoxin a in animal tissues and feeds in Poland in 2014–2016. J. Vet. Res. 2017, 61, 483–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, R.; Wolffram, S. Alkalinization of Urinary pH Accelerates Renal Excretion of Ochratoxin A in Pigs. J. Nutr. 2004, 134, 2355–2358. [Google Scholar] [CrossRef] [Green Version]
- Boudergur, C.; Burel, C.; Dragacci, S.; Favrot, M.; Fremy, J.; Massimi, C.; Prigent, P.; Debongnie, P.; Pussemier, L.; Boudra, H.; et al. Review of mycotoxin-detoxifying agents used as feed additives: Mode of action, efficacy and feed/food safety. EFSA Supporting Publ. 2017, 6, 22E. [Google Scholar] [CrossRef]
- Meissonnier, G.M.; Laffitte, J.; Raymond, I.; Benoit, E.; Cossalter, A.-M.; Pinton, P.; Bertin, G.; Oswald, I.P.; Galtier, P. Subclinical doses of T-2 toxin impair acquired immune response and liver cytochrome P450 in pigs. Toxicology 2008, 247, 46–54. [Google Scholar] [CrossRef]
- Takemura, H.; Shim, J.Y.; Sayama, K.; Tsubura, A.; Zhu, B.T.; Shimoi, K. Characterization of the estrogenic activities of zearalenone and zeranol In Vivo and In Vitro. J. Steroid Biochem. Mol. Biol. 2007, 103, 170–177. [Google Scholar] [CrossRef]
- Minervini, F.; Aquila, M.E.D. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistol, G.C.; Gras, M.A.; Marin, D.E.; Israel-Roming, F.; Stancu, M.; Taranu, I. Natural feed contaminant zearalenone decreases the expressions of important pro-and anti-inflammatory mediators and mitogen-activated protein kinase/NF-κB signalling molecules in pigs. Br. J. Nutr. 2014, 111, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.F.; Zhang, N.Y.; Peng, Y.Z.; Desheng, Q. Interaction of zearalenone and soybean isoflavone in diets on the growth performance, organ development and serum parameters in prepubertal gilts. J. Anim. Physiol. Anim. Nutr. 2012, 96, 939–946. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Chi, F.; Kim, I.H. Effects of montmorillonite clay on growth performance, nutrient digestibility, vulva size, faecal microflora, and oxidative stress in weaning gilts challenged with zearalenone. Anim. Feed Sci. Technol. 2012, 178, 158–166. [Google Scholar] [CrossRef]
- Brezina, U.; Rempe, I.; Kersten, S.; Valenta, H.; Humpf, H.U.; Dänicke, S. Diagnosis of intoxications of piglets fed with Fusarium toxin-contaminated maize by the analysis of mycotoxin residues in serum, liquor and urine with LC-MS/MS. Arch. Anim. Nutr. 2014, 68, 425–447. [Google Scholar] [CrossRef]
- Döll, S.; Dänicke, S.; Ueberschär, K.H.; Valenta, H.; Schnurrbusch, U.; Ganter, M.; Klobasa, F.; Flachowsky, G. Effects of graded levels of Fusarium toxin contaminated maize in diets for female weaned piglets. Arch. Anim. Nutr. 2003, 57, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, S.; Solfrizzo, H.; Visconti, A.; Powers, S.; Cossalter, A.M.; Pinton, P.; Oswald, I.P. Validation study on urinary biomarkers of exposure for aflatoxin B1, ochratoxin A, fumonisin B1, deoxynivalenol and zearalenone in piglets. World Mycotoxin J. 2013, 6, 299–308. [Google Scholar] [CrossRef]
- Thieu, N.Q.; Pettersson, H. Zearalenone, deoxynivalenol and aflatoxin B1 and their metabolites in pig urine as biomarkers for mycotoxin exposure. Mycotoxin Res. 2009, 25, 59–66. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, G.; Zhao, H.; Zheng, J.; Hu, F.; Fang, B. Liquid chromatography-tandem mass spectrometry method for toxicokinetics, tissue distribution, and excretion studies of T-2 toxin and its major metabolites in pigs. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 958, 75–82. [Google Scholar] [CrossRef]
- Fodor, J.; Meyer, K.; Riedlberger, M.; Bauer, J.; Horn, P.; Kovacs, F.; Kovacs, M. Distribution and elimination of fumonisin analogues in weaned piglets after oral administration of Fusarium verticillioides fungal culture. Food Addit. Contam. 2006, 23, 492–501. [Google Scholar] [CrossRef]
- Catteuw, A.; Broekaert, N.; De Baere, S.; Lauwers, M.; Gasthuys, E.; Huybrechts, B.; Callebaut, A.; Ivanova, L.; Uhlig, S.; De Boevre, M.; et al. Insights into In Vivo Absolute Oral Bioavailability, Biotransformation, and Toxicokinetics of Zearalenone, α-Zearalenol, β-Zearalenol, Zearalenone-14-glucoside, and Zearalenone-14-sulfate in Pigs. J. Agric. Food Chem. 2019, 67, 3448–3458. [Google Scholar] [CrossRef]
- Dänicke, S.; Brüssow, K.-P.; Valenta, H.; Ueberschär, K.-H.; Tiemann, U.; Schollenberger, M. On the effects of graded levels of Fusarium toxin contaminated wheat in diets for gilts on feed intake, growth performance and metabolism of deoxynivalenol and zearalenone. Mol. Nutr. Food Res. 2005, 49, 932–943. [Google Scholar] [CrossRef]
- Thanner, S.; Czeglédi, L.; Schwartz-Zimmermann, H.E.; Berthiller, F.; Gutzwiller, A. Urinary deoxynivalenol (DON) and zearalenone (ZEA) as biomarkers of DON and ZEA exposure of pigs. Mycotoxin Res. 2016, 32, 69–75. [Google Scholar] [CrossRef]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of zearalenone and its major modified forms in pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Hennig-Pauka, I.; Moll, W.D.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, G.S.; Pettersson, H.; Lindberg, J.E. Absorption, metabolism and excretion of 3-acetyl don in pigs. Arch. Anim. Nutr. 2003, 57, 335–345. [Google Scholar] [CrossRef]
- Jodlbauer, J.; Zöllner, P.; Lindner, W. Determination of zeranol, taleranol, zearalenone, α- and β-zearalenol in urine and tissue by high-performance liquid chromatography-tandem mass spectrometry. Chromatographia 2000, 51, 681–687. [Google Scholar] [CrossRef]
- Lauwers, M.; Croubels, S.; Letor, B.; Gougoulias, C.; Devreese, M. Biomarkers for Exposure as A Tool for Efficacy Testing of A Mycotoxin Detoxifier in Broiler Chickens and Pigs. Toxins 2019, 11, 187. [Google Scholar] [CrossRef] [Green Version]
- Brezina, U.; Valenta, H.; Rempe, I.; Kersten, S.; Humpf, H.U.; Dänicke, S. Development of a liquid chromatography tandem mass spectrometry method for the simultaneous determination of zearalenone, deoxynivalenol and their metabolites in pig serum. Mycotoxin Res. 2014, 30, 171–186. [Google Scholar] [CrossRef]
- Fleck, S.C.; Churchwell, M.I.; Doerge, D.R. Metabolism and pharmacokinetics of zearalenone following oral and intravenous administration in juvenile female pigs. Food Chem. Toxicol. 2017, 106, 193–201. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and Potential Effects on Humans. J. Toxicol. Environ. Health Part B 2005, 8, 39–69. [Google Scholar] [CrossRef]
- Lauwers, M.; De Baere, S.; Letor, B.; Rychlik, M.; Croubels, S.; Devreese, M. Multi LC-MS/MS and LC-HRMS methods for determination of 24 mycotoxins including major phase I and II biomarker metabolites in biological matrices from pigs and broiler chickens. Toxins 2019, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Altafini, A.; Armorini, S.; Zaghini, A.; Sardi, L.; Roncada, P. Tissue distribution of ochratoxin A in pigs after administration of two-levels contaminated diets. World Mycotoxin J. 2017, 10, 263–272. [Google Scholar] [CrossRef]
- Malagutti, L.; Zannotti, M.; Scampini, A.; Sciaraffia, F. Effects of Ochratoxin A on heavy pig production. Anim. Res. 2005, 54, 179–184. [Google Scholar] [CrossRef]
- Stoev, S.D.; Paskalev, M.; MacDonald, S.; Mantle, P.G. Experimental one year ochratoxin Atoxicosis in pigs. Exp. Toxicol. Pathol. 2002, 53, 481–487. [Google Scholar] [CrossRef]
- Aoudia, N.; Callu, P.; Grosjean, F.; Larondelle, Y. Effectiveness of mycotoxin sequestration activity of micronized wheat fibres on distribution of ochratoxin A in plasma, liver and kidney of piglets fed a naturally contaminated diet. Food Chem. Toxicol. 2009, 47, 1485–1489. [Google Scholar] [CrossRef]
- Pleadin, J.; Kudumija, N.; Kovačević, D.; Scortichini, G.; Milone, S.; Kmetič, I. Comparison of ochratoxin A levels in edible pig tissues and in biological fluids after exposure to a contaminated diet. Mycotoxin Res. 2016, 32, 145–151. [Google Scholar] [CrossRef]
- Perši, N.; Pleadin, J.; Kovačević, D.; Scortichini, G.; Milone, S. Ochratoxin A in raw materials and cooked meat products made from OTA-treated pigs. Meat Sci. 2014, 96, 203–210. [Google Scholar] [CrossRef]
- Tkaczyk, A.; Jedziniak, P.; Zielonka, Ł.; Dąbrowski, M.; Ochodzki, P.; Rudawska, A. Biomarkers of Deoxynivalenol, Citrinin, Ochratoxin A and Zearalenone in Pigs after Exposure of Naturally Contaminated Feed Close to Guidance Values; National Veterinary Research Institute: Puławy, Poland, 2021; manuscript in preparation.
- Dilkin, P.; Direito, G.; Simas, M.M.S.; Mallmann, C.A.; Correa, B. Toxicokinetics and toxicological effects of single oral dose of fumonisin B1 containing Fusarium verticillioides culture material in weaned piglets. Chem. Biol. Interact. 2010, 185, 157–162. [Google Scholar] [CrossRef]
- Souto, P.C.M.C.; Jager, A.V.; Tonin, F.G.; Petta, T.; Di Gregório, M.C.; Cossalter, A.M.; Pinton, P.; Oswald, I.P.; Rottinghaus, G.E.; Oliveira, C.A.F. Determination of fumonisin B1 levels in body fluids and hair from piglets fed fumonisin B1-contaminated diets. Food Chem. Toxicol. 2017, 108, 1–9. [Google Scholar] [CrossRef]
- Dietrich, D.R.; Heussner, A.H.; O’Brien, E. Ochratoxin A: Comparative pharmacokinetics and toxicological implications (experimental and domestic animals and humans). Food Addit. Contam. 2005, 22, 45–52. [Google Scholar] [CrossRef]
- Warth, B.; Sulyok, M.; Krska, R. LC-MS/MS-based multibiomarker approaches for the assessment of human exposure to mycotoxins. Anal. Bioanal. Chem. 2013, 405, 5687–5695. [Google Scholar] [CrossRef] [Green Version]
- Berthiller, F.; Crews, C.; Dall’asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Song, S.; Ediage, E.N.; Wu, A.; De Saeger, S. Development and application of salting-out assisted liquid/liquid extraction for multi-mycotoxin biomarkers analysis in pig urine with high performance liquid chromatography/tandem mass spectrometry. J. Chromatogr. A 2013, 1292, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Rui Guo, W.; Xiao Ou, S. Simultaneous Detection Method for Mycotoxins and their Metabolites in Animal Urine by Using Impurity Adsorption Purification followed by Liquid Chromatography-Tandem Mass Detection. J. Chromatogr. Sep. Tech. 2015, 6, 308. [Google Scholar] [CrossRef]
- Arndt, T. Urine-creatinine concentration as a marker of urine dilution: Reflections using a cohort of 45,000 samples. Forensic Sci. Int. 2009, 186, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Tkaczyk, A.; Jedziniak, P. Dilute-and-Shoot HPLC-UV Method for Determination of Urinary Creatinine as a Normalization Tool in Mycotoxin Biomonitoring in Pigs. Molecules 2020, 25, 2445. [Google Scholar] [CrossRef]
- Gambacorta, L.; Olsen, M.; Solfrizzo, M. Pig urinary concentration of mycotoxins and metabolites reflects regional differences, mycotoxin intake and feed contaminations. Toxins 2019, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Zöllner, P.; Jodlbauer, J.; Kleinova, M.; Kahlbacher, H.; Kuhn, T.; Hochsteiner, W.; Lindner, W. Concentration levels of zearalenone and its metabolites in urine, muscle tissue, and liver samples of pigs fed with mycotoxin-contaminated oats. J. Agric. Food Chem. 2002, 50, 2494–2501. [Google Scholar] [CrossRef]
- Shephard, G.S.; Burger, H.M.; Gambacorta, L.; Gong, Y.Y.; Krska, R.; Rheeder, J.P.; Solfrizzo, M.; Srey, C.; Sulyok, M.; Visconti, A.; et al. Multiple mycotoxin exposure determined by urinary biomarkers in rural subsistence farmers in the former Transkei, South Africa. Food Chem. Toxicol. 2013, 62, 217–225. [Google Scholar] [CrossRef]
- Ali, N.; Degen, G.H. Urinary biomarkers of exposure to the mycoestrogen zearalenone and its modified forms in German adults. Arch. Toxicol. 2018, 92, 2691–2700. [Google Scholar] [CrossRef] [PubMed]
- Huybrechts, B.; Martins, J.C.; Debongnie, P.; Uhlig, S.; Callebaut, A. Fast and sensitive LC-MS/MS method measuring human mycotoxin exposure using biomarkers in urine. Arch. Toxicol. 2015, 89, 1993–2005. [Google Scholar] [CrossRef]
- Gerding, J.; Ali, N.; Schwartzbord, J.; Cramer, B.; Brown, D.L.; Degen, G.H.; Humpf, H.U. A comparative study of the human urinary mycotoxin excretion patterns in Bangladesh, Germany, and Haiti using a rapid and sensitive LC-MS/MS approach. Mycotoxin Res. 2015, 31, 127–136. [Google Scholar] [CrossRef]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Fruhmann, P.; Hametner, C.; Adam, G.; Fröhlich, J.; Krska, R. Direct quantification of deoxynivalenol glucuronide in human urine as biomarker of exposure to the Fusarium mycotoxin deoxynivalenol. Anal. Bioanal. Chem. 2011, 401, 195–200. [Google Scholar] [CrossRef]
- Tkaczyk, A.; Jedziniak, P. Development of a multi-mycotoxin LC-MS/MS method for the determination of biomarkers in pig urine. Mycotoxin Res. 2021. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; De Mil, T.; Fraeyman, S.; De Baere, S.; De Saeger, S.; De Backer, P.; Croubels, S. Development and validation of an LC–MS/MS method for the toxicokinetic study of deoxynivalenol and its acetylated derivatives in chicken and pig plasma. J. Chromatogr. B 2014, 971, 43–51. [Google Scholar] [CrossRef]
- De Baere, S.; Osselaere, A.; Devreese, M.; Vanhaecke, L.; De Backer, P.; Croubels, S. Development of a liquid-chromatography tandem mass spectrometry and ultra-high-performance liquid chromatography high-resolution mass spectrometry method for the quantitative determination of zearalenone and its major metabolites in chicken and pig plasma. Anal. Chim. Acta 2012, 756, 37–48. [Google Scholar] [CrossRef]
- Devreese, M.; De Baere, S.; De Backer, P.; Croubels, S. Quantitative determination of the Fusarium mycotoxins beauvericin, enniatin A, A1, B and B1 in pig plasma using high performance liquid chromatography–tandem mass spectrometry. Talanta 2013, 106, 212–219. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Pig Type | Adverse Effect | LOAEL | NOAEL | Ref. |
|---|---|---|---|---|---|
| DON | - | vomiting | 2.8 mg/kg feed | 0.7–12 mg/kg feed | [17] |
| reduced feed intake and weight gain | 0.35 mg/kg feed | 0.7 mg/kg feed | |||
| FB1 | - | lung lesions | 0.4 mg/kg b.w./day | - | [20] |
| increased serum Sa:So (sphinganine:sphingosine) ratios | 5 mg /kg feed (0.2 mg/kg b.w./day | - | |||
| OTA | - | progressive nephropathy | 1 mg/kg feed (40 µg/kg b.w.) | - | [19] |
| effects on renal (diagnostic) enzyme levels and kidney function | 0.2 mg/kg feed (8 µg/kg b.w.) | - | |||
| T-2 | - | immunological or haematological effects | 0.2 mg/kg feed (8 µg/kg b.w.) | - | [21] |
| ZEN | female prepubertal piglets | oestrogenic effects such as increased uterus weight and reddened and swollen vulva | 17.6 µg/kg b.w./day | 10.4 µg/kg b.w./day | [18] |
| sexually mature female pigs | prolonged cycling | 200 µg/kg b.w./day | 40 µg/kg b.w./day |
| ZEN Feeding Group | Dose [µg/kg Feed] | Feed Type | Matrix 1 | Biomarker | Biomarker Concentration (ng/mL) | Matrix 2 | Biomarker | Biomarker Concentration (ng/mL) | Experiment Time | Number and Type of Pig | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4 | naturally contaminated wheat | urine | ZEN α-ZEL | 10.4 16.4 | serum | - | - | 35 days | 12–180 days old gilts | [54] |
| 2 | 88 | 16.7 18.3 | |||||||||
| 3 | 235 | 15.4 22.1 | |||||||||
| 4 | 358 | 43.1 57.9 | |||||||||
| 1 | 10 | naturally contaminated maize | urine | ZEN α-ZEL | 2.7 ** 1.5 | - | - | - | 37 days | 100 female piglets | [48] |
| 2 | 60 | 34.8 10.2 | |||||||||
| 3 | 150 | 31.5 9.9 | |||||||||
| 4 | 220 | 64.1 20.3 | |||||||||
| 1 | 350 | naturally contaminated maize | urine | ZEN ZEN-GlcAc α-ZEL α-ZEL-GlcAc | - | - | - | - | 28 days | 24 castrated male pigs | [55] |
| 2 | 720 | ||||||||||
| 3 | 1680 | ||||||||||
| 4 | 2360 | ||||||||||
| 1 | 10 | contaminated maise | urine | ZEN α-ZEL β-ZEL ZAN α-ZAL | 3.4 * (0.5–11.4) 0.6 (<LOQ–35.2) <LOQ <LOQ <LOQ | serum | ZEN α-ZEL | 0 0 | 29 days | 125 female weaned piglets | [47] |
| 2 | 50 | 25.9 (1.0–82.0) 9.7 (0–22.7) <LOQ <LOQ <LOQ | 0–0.1 0 | ||||||||
| 3 | 80 | 51.7 (1.1–122) 18.6 (0–63.2) 1.4 (<LOQ–18.2) 0.4 (<LOQ–1.4) <LOQ | 0–0.2 0 | ||||||||
| 4 | 170 | 79 (9.5–237) 35.1 (2.3–89.2) 10.2 (<LOQ–42.1) 0.8 (<LOQ–2.3) <LOQ | 0–0.3 0–0.8 | ||||||||
| 5 | 290 | 136 (7.7–327) 57.2 (2.4–122) 15.1 (0–42.9) 1.7 (0–3.4) 0.5 (0–0.7) | 0–0.5 0–1.6 | ||||||||
| 1 | 6.04 | feed boluses fortified with pure cultures | urine | ZEN α-ZEL β-ZEL | 11.63 ± 7.52 3.60 ± 3.70 nd | - | - | - | 3 days | 16 weaned piglets | [49] |
| 2 | 17.23 | 23.06 ± 11.42 6.76 ± 4.43 nd | |||||||||
| 3 | 21.13 | 15.76 ± 9.62 5.24 ± 3.29 nd | |||||||||
| 4 | 50.26 | 17.08 ± 4.15 6.58 ± 2.36 0.39 ± 0.36 |
| Feeding Group | Dose (µg/kg Feed) | Feed Type | Matrix 1 | Biomarker | Biomarker Level (ng/mL) | Matrix 2 | Biomarker | Biomarker Level (ng/mL) | Experiment Time | Number and Type of Pig | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 210 | contaminated wheat | urine | DON DOM-1 | 299 29 | serum | DONDOM-1 | 1 0.1 | 35 days | 12–180 days old gilts | [54] |
| 2 | 3070 | 690 150 | 4.1 1.3 | ||||||||
| 3 | 6100 | 804 177 | 14.3 2.8 | ||||||||
| 4 | 9570 | 1572 289 | 21.6 4.1 | ||||||||
| 1 | 200 | contaminated maize | urine | DON DOM-1 | 56 <LOD | serum | DONDOM-1 | <LOQ | 37 days | 100 female piglets | [48] |
| 2 | 800 | 162 <LOD | 1 (<LOQ-4) | ||||||||
| 3 | 1000 | 360 14 | 5 (<LOQ-12) | ||||||||
| 4 | 1900 | 246 14 | 6 (<LOQ-13) | ||||||||
| 5 | 3900 | 380 19 | 11 (6–19) | ||||||||
| 1 | 1110 | contaminated maize | urine | - | - | - | - | 28 days | 24 castrated male pigs | [55] | |
| 2 | 2320 | ||||||||||
| 3 | 3700 | ||||||||||
| 4 | 5000 | ||||||||||
| 1 | 30 | contaminated maize | urine | DONDOM-1 | 68.3 (18.6–231) 20.4 (5.3–171) | serum | DONDOM-1 | 0.5 (0–1.4) 0 | 29 days | 125 female weaned piglets | [47] |
| 2 | 590 | 524 (50.8–1070) 43.4 (1.8–140) | 4.9 (2.7–7.9) 0 | ||||||||
| 3 | 1270 | 1065 (96.2–2120) 70.3 (1.6–336) | 8.6 (4.6–15.9) 0 | ||||||||
| 4 | 2010 | 1850 (288–4050) 118 (1.2–513) | 15.2 (8.1–24.9) 2.9 (0–4.0) | ||||||||
| 5 | 4520 | 2680 (244–4990) 262 (0.7–979) | 24.4 (9.9–42.8) 4.5 (0–7.4) | ||||||||
| 1 | 63.58 | fortified feed boluses | urine | DONDOM-1 | 80.55 ± 28.96 1.19 ± 1.38 | - | - | - | 3 days | 16 weaned piglets | [49] |
| 2 | 181.51 | 125.06 ± 41.64 3.36 ±1.35 | |||||||||
| 3 | 214.36 | 305.94 ± 143.17 3.44 ± 4.14 | |||||||||
| 4 | 509.53 | 218.18 ± 33.35 16.33 ± 5.95 |
| Mycotoxin | Feeding Group | Dose | Feed Type | Matrix 1 | Biomarker | Biomarker Level (ng/mL) | Matrix 2 | Biomarker | Biomarker Level (ng/mL) | Experiment Time | Number and Type of Pig | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AFB1 | 1 | 0.16 µg/kg b.w. | feed boluses fortified with pure cultures | urine | AFM1 | 0.14 | - | 3 days | 16 weaned piglets | [49] | ||
| 2 | 0.45 µg/kg b.w. | 0.36 | ||||||||||
| 3 | 0.54 µg/kg b.w. | 0.50 | ||||||||||
| 4 | 1.28 µg/kg b.w. | 0.88 | ||||||||||
| 1 | 127 µg/kg feed | pure crystal AFB1 dissolved in methanol (20 mL) and sprayed on 2 kg of feed | urine | AFM1 AFB1 AFB2 | 12.6 9.9 0.64 | - | 18 days | 4 castrated male pigs | [50] | |||
| 2 | 227 µg/kg feed | 17.2 1.9 0.76 | ||||||||||
| 3 | 327 µg/kg feed | 22.6 3.7 0.51 | ||||||||||
| OTA | 1 | 0.16 µg/kg b.w. | feed boluses fortified with mycotoxins | urine | OTA | 0.12 | - | 3 days | 16 weaned piglets | [49] | ||
| 2 | 0.46 µg/kg b.w. | 0.65 | ||||||||||
| 3 | 0.56 µg/kg b.w. | 0.52 | ||||||||||
| 4 | 1.32 µg/kg b.w. | 0.36 | ||||||||||
| OTA | 1 | 50 µg/kg feed | basal diet mixed with pure OTA standard solutions | plasma | OTA | 22.2 ± 2.6 | - | 15 days | 12 pigs | [65] | ||
| 2 | 500 µg/kg feed | 217.4 ± 25.1 | ||||||||||
| 1 | 25 µg/kg feed | crystalline OTA | urine | OTA | 3.1–4.35 | - | 119 days | 24 pigs | [66] | |||
| 1 | 800 µg/kg feed | naturally contaminated feed | serum | OTA | 852–1582 | - | 180 days | 6 pigs | [67] | |||
| 1 | 120 µg/kg feed | naturally contaminated feed | plasma | OTA | 82.8 | 28 days | 48 piglets | [68] | ||||
| 1 | 250 µg/kg feed | OTA-fortified feed | urine | OTA | 16.1 | serum | OTA | 4.8 | 28 days | 10 pigs | [69] | |
| 1 | 300 µg/kg feed | pure OTA standard mixed with lactose and formulated as gelatine capsules | plasma | OTA | 6.4 | 30 days | 10 pigs | [70] | ||||
| 1 | 114 µg/kg feed | naturally contaminated feed | urine | OTA OTα | 18.8 ± 6.4 9.5 ± 2.6 | serum | OTA OTα | 141 ± 47.9 < LLOQ | 28 days | 24 pigs | [71] | |
| 2 | 226 µg/kg feed | 36.5 ± 11.6 16.2 ± 5.5 | 278 ± 106 0.69 ± 0.10 | |||||||||
| FB1 | 1 | 3.71 µg/kg b.w. | feed boluses fortified with pure cultures | urine | FB1 | 1.55 | - | 3 days | 16 weaned piglets | [49] | ||
| 2 | 10.6 µg/kg b.w. | 3.36 | ||||||||||
| 3 | 64.2 µg/kg b.w. | 77.37 | ||||||||||
| 4 | 150 µg/kg b.w. | 117.78 | ||||||||||
| 1 | 5 mg/kg b.w. | aqueous stock solutions | urine | FB1 | - | faeces | FB1 | - | 96 h | male weaned piglets | [72] | |
| 1 | 3.1 µg/g | culture material added to feed | urine | FB1 | 16.09 ± 21.94 | plasma | FB1 | 0.16 ± 0.04 | 28 days | 24 piglets | [73] | |
| 2 | 6.1 µg/g | 24.08 ± 25.96 | 0.26 ± 0.06 | |||||||||
| 3 | 9.0 µg/g | 18.88 ± 4.41 | 0.42 ± 0.10 | |||||||||
| FB1 FB2 FB3 | 1 | 50 mg/animal | F.verticillioides fungal culture mixed into feed | urine (13–17 day) | FB1 FB2 | 4.5 ± 3.9 mg 0.5 ± 0.5 mg | faeces | FB1FB2 | 28.2 ± 27.3 mg 2 ± 1.1 mg | 22 days | 6 cross-bred pigs (plasma) /15 weaned barrows (urine) | [52] |
| 2 | 20 mg/animal | |||||||||||
| 3 | 5 mg/animal | |||||||||||
| T-2 | 1 | 500 | aqueous stock solutions | urine | T-2 HT-2 T-2 triol | 30.9 ± 2.1 614.4 ± 177 306 ± 70 | faeces | HT-2 | 104.6 ± 14.2 (36 h) | 1 day | 7 cross-bred pigs | [51] |
| Mycotoxin/Matrix/Time | Urine | Faeces | Plasma |
|---|---|---|---|
| DON | DON (4–8 h) DOM-1 DON-GlcAc | DOM-1 (8–24 h) DON | DON-3-GlcAc (3–4 h) DON * (3–4 h) DOM-1 |
| ZEN | ZEN-GlcAc (12–24 h) ZEN * (12–24 h) α-ZEL β-ZEL ZAN α-ZAL β-ZAL | ZEN (12–24 h) α-ZEL (12–24 h) ZAN | ZEN-GlcAc (0.25–3 h) ZEN * (0.25–3 h) α-ZEL (0.25–4 h) |
| OTA | OTA OTα | n.d. | OTA (72–120 h) OTα |
| FB1/FB2 | FB1 (8–24 h) FB2 | FB1 (8–24 h) FB2 | FB1 (2 h) |
| AFB1 | AFM1 AFB1 AFB2 | n.d. | n.d. |
| T-2 | HT-2 (12–24 h) T-2 triol (12–24 h) T-2 | HT-2 | T-2 (3 h) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tkaczyk, A.; Jedziniak, P. Mycotoxin Biomarkers in Pigs—Current State of Knowledge and Analytics. Toxins 2021, 13, 586. https://doi.org/10.3390/toxins13080586
Tkaczyk A, Jedziniak P. Mycotoxin Biomarkers in Pigs—Current State of Knowledge and Analytics. Toxins. 2021; 13(8):586. https://doi.org/10.3390/toxins13080586
Chicago/Turabian StyleTkaczyk, Agnieszka, and Piotr Jedziniak. 2021. "Mycotoxin Biomarkers in Pigs—Current State of Knowledge and Analytics" Toxins 13, no. 8: 586. https://doi.org/10.3390/toxins13080586
APA StyleTkaczyk, A., & Jedziniak, P. (2021). Mycotoxin Biomarkers in Pigs—Current State of Knowledge and Analytics. Toxins, 13(8), 586. https://doi.org/10.3390/toxins13080586