Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants?


Abstract
:1. Introduction
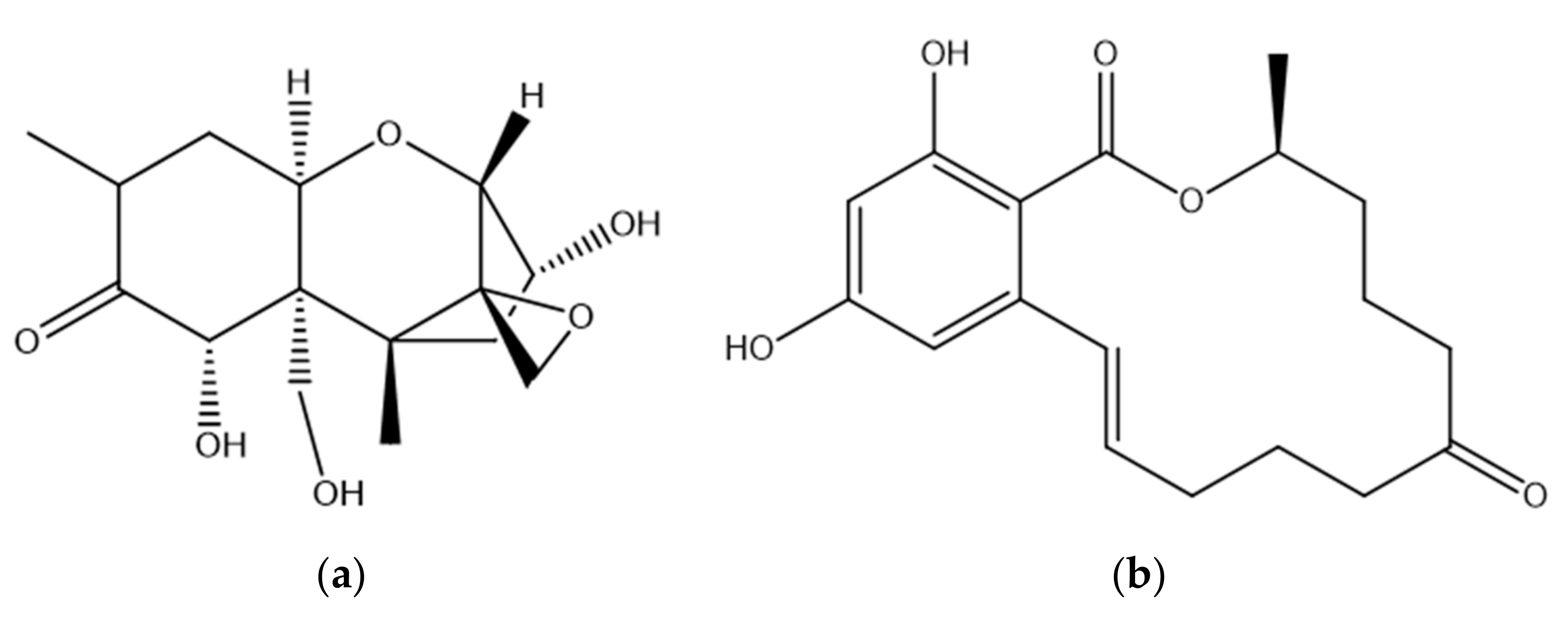
2. Deoxynivalenol
2.1. DON Occurrence
2.2. DON Toxicity
3. Zearalenone
3.1. ZEN Occurrence
3.2. ZEN Toxicity
4. Co-Occurrence of DON and ZEN
5. In Vitro Studies on Co-Exposure to DON and ZEN

| Cell Line Used | Toxin Levels | Exposure Time and Route | Conclusions | Observations | Comment | References |
| HepaRG | DON 0.2–10 µM ZEN 1.5–75 µM | 48 h | Synergistic at 48 h cell viability | Cell viability | Doses correspond to IC50 values after 48 h | [105] |
| DON 7.35 µM ZEN 55.1 µM | 18 h | Additive at 18 h cell mortality | Cell mortality | |||
| HepaRG | DON 2.5 µM ZEN 0.24 µM | 14 days 28 days 42 days | Additive | Cell viability | Doses correspond to maximum levels permitted in cereals for humans | [106] |
| HepaRG | DON 0.2 µM ZEN 20 µM | 1 and 24 h | Synergistic at 1 h Antagonistic at 24 h | Cell proteome | Doses correspond to IC10 values after 48 h | [107] |
| BEL-7402 | DON 0.37–16.9 µM ZEN 0.31–31.4 µM | 24 h | Additive | Cell viability | Mixtures used DON + ZEN 0.37 + 0.31 µM 0.68 + 0.63 µM 1.69 + 1.57 µM 3.7 + 3.1 µM 6.8 + 15.7 µM 16.9 + 31.4 µM | [108] |
| HepG2 | DON 0.34–67.5 µM ZEN 15.7–157 µM | 24 h | Synergistic | Cell viability | Mixtures used DON + ZEN 0.034 + 15.7 µM 1.69 + 47.1 µM 3.7 + 62.8 µM 37 + 126 µM 67.5 + 157 µM | [109] |
| HepG2 | DON 0.02–2 µM ZEN 0.28–34.5 µM | 48 h | Synergistic | Cell viability | DON + ZEN 0.02 + 0.28 µM 0.03 + 0.53 µM 0.07 + 1.1 µM 0.14 + 3 µM 0.27 + 4.4 µM 0.51 + 8.6 µM 1 + 17.3 µM 2 + 34.5 µM | [110] |
| RAW 246.7 | DON 0.0027–0.34 µM ZEN 0.28–37.69 µM | 48 h | Synergistic | Cell viability | DON + ZEN 0.0027 + 0.28 µM 0.0054 + 0.6 µM 0.01 + 1.19 µM 0.02 + 2.36 µM 0.04 + 4.71 µM 0.08 + 9.42 µM 0.17 + 18.8 µM 0.34 + 37.69 µM | [110] |
| Caco-2 | DON 3.3–16.7 µM ZEN 10–50 µM | 24 h | Antagonistic | Cell viability | DON and ZEN combination in 1:3 ratio | [112] |
| HCT116 | DON 100 µM ZEN 40 µM | 24 h | Antagonistic | Cytotoxicity, mitochondrial apoptosis | Doses correspond to IC30 values after 24 h | [113] |
| 48 h | Cell cycle analysis | |||||
| IPEC-J2 | Cytotoxic concentration DON 2 µM DON ZEN 40 µM | 48 h | Cytotoxic concentration Reported as non-additive | Cell viability | Dose correspond to cytotoxic and non-cytotoxic concentrations | [90] |
| Non-cytotoxic concentration DON 0.5 µM ZEN 10 µM | Non-cytotoxic concentration Synergistic | |||||
| PK15 | DON 0.25 µM ZEN 20 µM | 24 h | Synergistic | ROS levels Apoptosis | Doses used are concentrations close to IC10 concentration which were 0.157 and 27.583 µM for DON and ZEN, respectively | [114] |
| Porcine splenic lymphocytes | DON + ZEN 0.2 + 0.25 µM 1 + 1.26 µM 5.1 + 6.28 µM | 48 h | Synergistic | Apoptosis Oxidative injury | [115] | |
| Porcine lymphocytes | DON + ZEN 0.24 + 15.7 μM 0.71 + 31.4 μM | 24/48/72 h | Antagonistic | Cell viability | Doses used were below IC50 concentration after 24, 48, 72 h exposure | [116] |
| Antagonistic at lower concentration Synergistic at 72 h at higher concentration | Genotoxicity | |||||
| THP-1 | DON 0.1–10 μM ZEN 2–100 μM | 48 h | Antagonistic | Cell viability | DON + ZEN + 2 μM 0.8 + 16 μM 2 + 40 μM 4 + 80 μM 10 + 100 μM | [117] |
| ANA-1 | DON 0–33.7 µM ZEN 0–37.7 µM | 24 h | Synergistic | Cell viability and apoptosis | DON + ZEN concentration used for apoptosis and metabolism study 0.34 + 25.1 µM | [118] |
| Antagonistic | Cell metabolism | |||||
| BF-2 | DON 0–16.2 µM ZEN 0–120.3 µM | 48 h | Antagonism | Cell viability fish Oxidative stress fish | DON + ZEN 0.13 + 1.33 0.25 + 2.66 0.51 + 5.32 1.01 + 10.64 2.02 + 21.29 4.05 + 42.58 8.1 + 85.15 16.2 + 170.3 | [119] |
| Caco-2, HepaRG and THP-1 Co-culture | DON + ZEN Concentration used when Caco-2 cells were in luminal compartment: 1.6 + 24 µM 3 + 31 µM Concentration used when HepaRG cells were in luminal compartment: +20 µM 2.3 + 33 µM | 48 h | No cytotoxicity with low concentration and in tri-culture Synergistic effect with higher concentration in bi-culture system | Cell viability | [120] | |
| Animal used | Toxin levels | Exposure time and route | Conclusions | Observations | Comment | References |
| Zebrafish larvae | DON 67.5 µM ZEN 6.28 µM | 72 h | Antagonistic | Cell mortality | [119] | |
| Mice | DON 1500, 2500 µg/kg bodyweight ZEN 20,000, 30,000 µg/kg bodyweight | 12 days Intraperitoneal injection | Antagonistic | Oxidative stress Renal apoptosis | DON + ZEN 1500 + 20,000 1500 + 30,000 2500 + 20,000 25 + 30,000 µg/kg bodyweight | [121] |
| Mice | DON 5000 µg/kg bodyweight ZEN 5000 µg/kg bodyweight | 2 weeks Oral administration | Antagonistic | Oxidative stress | No change observed on liver weight | [111] |
| Synergistic | Apoptosis | |||||
| Rats | DON 30 µg/animal/day ZEN 15 µg/animal/day | 14 days Administered as a gavage dose | Antagonistic | Liver weight Glutathione level in liver Malondialdehyde level in kidney | Doses are according to EU limits in finished feed for young pigs | [122] |
| Mice | DON 0.5–2 μM ZEN 10–40 μM | 24 h | Synergistic | Cell viability Immune function | DON and ZEN combined 1:20 | [123] |
| Rats | DON 16.5 µg/animal/day ZEN 12.75 µg/animal/day | 5 days Intraperitoneal administration | Synergistic | Glutathione and glutathione peroxidase activity in the liver | Doses correspond to 1 mg/kg diet for DON and 1.5 mg/kg diet for ZEN which are close to EU limits in finished feed for young pigs | [124] |
| Mice | DON 2000 mg/kg ZEN 20,000 mg/kg | 21 days Intragastric administration | Antagonistic | Metabolic profiling of liver and serum | [125] | |
| Mice | DON 2000 mg/kg ZEN 20,000 mg/kg | 3 weeks Intragastric administration | Antagonistic | Metabolic pathway | [126] | |
| Mice | DON 1500, 2500 µg/kg body weight ZEN 20,000, 30,000 µg/kg body weight | 4 days Intraperitoneal injection | Synergistic | Apoptosis Antioxidant levels | DON + ZEN 1500 + 20,000 1500 + 30,000 2500 + 20,000 2500 + 30,000 | [127] |
| Female piglets | DON 1000.6 µg/kg ZEN 269.1 µg/kg DON + ZEN 1007.5 + 265.4 µg/kg | 3 weeks Ad libitum feeding | Synergistic | Body weight gain Average daily feed intake Intestinal functions | Barley naturally contaminated with DON and corn naturally contaminated with ZEN was used to manufacture the feed | [128] |
6. In Vivo Studies on Co-Exposure to DON and ZEN
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.-S.; Groopman, J.D. DNA Damage by Mycotoxins. Mutat. Res. 1999, 424, 167–181. [Google Scholar] [CrossRef]
- Peraica, M.; Radić, B.; Lucić, A.; Pavlović, M. Toxic Effects of Mycotoxins in Humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar]
- Hussein, H. Toxicity, Metabolism, and Impact of Mycotoxins on Humans and Animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin Occurrence in Feed and Feed Raw Materials Worldwide: Long-Term Analysis with Special Focus on Europe and Asia: Mycotoxin Occurrence in Feed and Feed Raw Materials Worldwide. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, Toxicology, and Exposure Assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.G.; Ross, P.F. Methods for Detection and Quantitation of Fumonisins in Corn, Cereal Products and Animal Excreta. J. Food Prot. 1994, 57, 536–540. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin Contamination of the Feed Supply Chain: Implications for Animal Productivity and Feed Security. Anim. Feed Sci. Tech. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Pestka, J.; Casale, W. Naturally Occurring Fungal Toxins. Environ. Sci. Technol. 1990, 23, 613–638. [Google Scholar]
- Jackson, L.S.; Bullerman, L.B. Effect of Processing on Fusarium Mycotoxins. In Impact of Processing on Food Safety; Jackson, L.S., Knize, M.G., Morgan, J.N., Eds.; Springer: Boston, MA, USA, 1999; Volume 459, pp. 243–261. ISBN 978-1-4613-7201-1. [Google Scholar]
- Sarma, U.P.; Bhetaria, P.J.; Devi, P.; Varma, A. Aflatoxins: Implications on Health. Ind. J. Clin. Biochem. 2017, 32, 124–133. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of Trichothecene Mycotoxins on Eukaryotic Cells: A Review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef]
- Bui-Klimke, T.R.; Wu, F. Ochratoxin A and Human Health Risk: A Review of the Evidence. Crit. Rev. Food Sci. Nutr. 2015, 55, 1860–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and Its Metabolites: Effect on Human Health, Metabolism and Neutralisation Methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an Estrogenic Mycotoxin, Is an Immunotoxic Compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J. Deoxynivalenol: Toxicity, Mechanisms and Animal Health Risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Nagy, C.M.; Fejer, S.N.; Berek, L.; Molnar, J.; Viskolcz, B. Hydrogen Bondings in Deoxynivalenol (DON) Conformations—A Density Functional Study. J. Mol. Struct. 2005, 726, 55–59. [Google Scholar] [CrossRef]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; De Saeger, S.; Yang, H.; Zhang, Q.; Wu, A. Detoxification of Deoxynivalenol via Glycosylation Represents Novel Insights on Antagonistic Activities of Trichoderma When Confronted with Fusarium Graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef] [Green Version]
- Ueno, Y.; Nakajima, M.; Sakai, K.; Ishii, K.; Sato, N.; Shimada, N. Comparative Toxicology of Trichothec Mycotoxins: Inhibition of Protein Synthesis in Animal Cells. J. Biochem. 1973, 74, 285–296. [Google Scholar]
- Karlovsky, P. Biological Detoxification of the Mycotoxin Deoxynivalenol and Its Use in Genetically Engineered Crops and Feed Additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Dohnal, V.; Kuca, K.; Yuan, Z. Trichothecenes: Structure-Toxic Activity Relationships. Curr. Drug Metab. 2013, 14, 641–660. [Google Scholar] [CrossRef]
- Sundstøl Eriksen, G.; Pettersson, H.; Lundh, T. Comparative Cytotoxicity of Deoxynivalenol, Nivalenol, Their Acetylated Derivatives and de-Epoxy Metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Mayer, E.; Novak, B.; Springler, A.; Schwartz-Zimmermann, H.E.; Nagl, V.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G. Effects of Deoxynivalenol (DON) and Its Microbial Biotransformation Product Deepoxy-Deoxynivalenol (DOM-1) on a Trout, Pig, Mouse, and Human Cell Line. Mycotoxin Res. 2017, 33, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and Its Metabolite Deepoxy-Deoxynivalenol: Multi-Parameter Analysis for the Evaluation of Cytotoxicity and Cellular Effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the Toxicity, Occurrence, Metabolism, Detoxification, Regulations and Intake of Zearalenone: An Oestrogenic Mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Daigle, K.W. Protein Synthesis Inhibition by 8-Oxo-12,13-Epoxytrichothecenes. Biochim. Biophys. Acta 1987, 923, 206–213. [Google Scholar] [CrossRef]
- EFSA Opinion of the Scientific Panel on Contaminants in the Food Chain [CONTAM] Related to Deoxynivalenol (DON) as Undesirable Substance in Animal Feed. EFSA J. 2004, 2, 73. [CrossRef]
- De Walle, J.V.; Sergent, T.; Piront, N.; Toussaint, O.; Schneider, Y.-J.; Larondelle, Y. Deoxynivalenol Affects in Vitro Intestinal Epithelial Cell Barrier Integrity through Inhibition of Protein Synthesis. Toxicol. Appl. Pharmacol. 2010, 245, 291–298. [Google Scholar] [CrossRef]
- Dänicke, S.; Goyarts, T.; Döll, S.; Grove, N.; Spolders, M.; Flachowsky, G. Effects of the Fusarium Toxin Deoxynivalenol on Tissue Protein Synthesis in Pigs. Toxicol. Lett. 2006, 165, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, K.; Awad, W.A.; Soodoi, C.; Sasgary, S.; Strasser, A.; Böhm, J. Effects of Feed Contaminant Deoxynivalenol on Plasma Cytokines and MRNA Expression of Immune Genes in the Intestine of Broiler Chickens. PLoS ONE 2013, 8, e71492. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Srivastava, S.; Dewangan, J.; Divakar, A.; Kumar Rath, S. Global Occurrence of Deoxynivalenol in Food Commodities and Exposure Risk Assessment in Humans in the Last Decade: A Survey. Crit. Rev. Food Sci. Nutr. 2019, 60, 1346–1374. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Naehrer, K. A Three-Year Survey on the Worldwide Occurrence of Mycotoxins in Feedstuffs and Feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Chang, H.; Kim, D.; Chung, S.; Lee, C. Long-Term Occurrence of Deoxynivalenol in Feed and Feed Raw Materials with a Special Focus on South Korea. Toxins 2018, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of Deoxynivalenol (Vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Anukul, N.; Vangnai, K.; Mahakarnchanakul, W. Significance of Regulation Limits in Mycotoxin Contamination in Asia and Risk Management Programs at the National Level. J. Food Drug Anal. 2013, 21, 227–241. [Google Scholar] [CrossRef] [Green Version]
- U.S. Food and Drug Administration Guidance for Industry and FDA: Advisory Levels for Deoxynivalenol (DON) in Finished Wheat Products for Human Consumption and Grains and Grain By-Products Used for Animal Feed. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-fda-advisory-levels-deoxynivalenol-don-finished-wheat-products-human (accessed on 30 July 2021).
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs; European Commission: Brussels, Belgium; Luxembourg, 2006. [Google Scholar]
- European Commission. Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding; European Commission: Brussels, Belgium; Luxembourg, 2006; Volume 229. [Google Scholar]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A Major Player in the Multifaceted Response of Fusarium to Its Environment. Toxins 2013, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and Potential Effects on Humans. J. Toxicol. Environ. Health B Crit. Rev. 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Hoogenboom, L.A.P.; Bokhorst, J.G.; Northolt, M.D.; van de Vijver, L.P.L.; Broex, N.J.G.; Mevius, D.J.; Meijs, J.A.C.; Van der Roest, J. Contaminants and Microorganisms in Dutch Organic Food Products: A Comparison with Conventional Products. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2008, 25, 1195–1207. [Google Scholar] [CrossRef]
- Giraud, F.; Pasquali, M.; Jarroudi, M.E.; Vrancken, C.; Brochot, C.; Cocco, E.; Hoffmann, L.; Delfosse, P.; Bohn, T. Fusarium Head Blight and Associated Mycotoxin Occurrence on Winter Wheat in Luxembourg in 2007/2008. Food Addit. Contam. Part A 2010, 27, 825–835. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; de Rijk, T.C.; Booij, C.J.H.; Goedhart, P.W.; Boers, E.a.M.; Zhao, C.; Waalwijk, C.; Mol, H.G.J.; van der Lee, T.a.J. Occurrence of Fusarium Head Blight Species and Fusarium Mycotoxins in Winter Wheat in the Netherlands in 2009. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2012, 29, 1716–1726. [Google Scholar] [CrossRef]
- González, H.H.L.; Moltó, G.A.; Pacin, A.; Resnik, S.L.; Zelaya, M.J.; Masana, M.; Martínez, E.J. Trichothecenes and Mycoflora in Wheat Harvested in Nine Locations in Buenos Aires Province, Argentina. Mycopathologia 2008, 165, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Xu, J.; Liu, X.; Yin, X.; Shi, J. Natural Occurrence of Deoxynivalenol and Zearalenone in Wheat from Jiangsu Province, China. Food Chem. 2014, 157, 393–397. [Google Scholar] [CrossRef]
- Shephard, G.S.; van der Westhuizen, L.; Katerere, D.R.; Herbst, M.; Pineiro, M. Preliminary Exposure Assessment of Deoxynivalenol and Patulin in South Africa. Mycotoxin Res. 2010, 26, 181–185. [Google Scholar] [CrossRef]
- Waché, Y.J.; Valat, C.; Postollec, G.; Bougeard, S.; Burel, C.; Oswald, I.P.; Fravalo, P. Impact of Deoxynivalenol on the Intestinal Microflora of Pigs. Int. J. Mol. Sci. 2008, 10, 1–17. [Google Scholar] [CrossRef]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a Pig Jejunal Explant Culture for Studying the Gastrointestinal Toxicity of the Mycotoxin Deoxynivalenol: Histopathological Analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef]
- Awad, W.A.; Aschenbach, J.R.; Zentek, J. Cytotoxicity and Metabolic Stress Induced by Deoxynivalenol in the Porcine Intestinal IPEC-J2 Cell Line: Deoxynivalenol Effects on Intestinal Epithelial Cells. J. Anim. Physiol. Anim. Nutr. 2012, 96, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, L.; Geng, C.; Cao, J.; Zhong, L. The Role of Oxidative Stress in Deoxynivalenol-Induced DNA Damage in HepG2 Cells. Toxicon 2009, 54, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Bony, S.; Carcelen, M.; Olivier, L.; Devaux, A. Genotoxicity Assessment of Deoxynivalenol in the Caco-2 Cell Line Model Using the Comet Assay. Toxicol. Lett. 2006, 166, 67–76. [Google Scholar] [CrossRef]
- Le Hégarat, L.; Takakura, N.; Simar, S.; Nesslany, F.; Fessard, V. The In Vivo Genotoxicity Studies on Nivalenol and Deoxynivalenol. EFSA Support. Publ. 2014, 11, 697E. [Google Scholar] [CrossRef] [Green Version]
- Groschwitz, K.R.; Hogan, S.P. Intestinal Barrier Function: Molecular Regulation and Disease Pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of Deoxynivalenol and Its Acetylated and Modified Forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for in Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Pinton, P.; Nougayrède, J.-P.; Del Rio, J.-C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.-P.; Kolf-Clauw, M.; Oswald, I.P. The Food Contaminant Deoxynivalenol, Decreases Intestinal Barrier Permeability and Reduces Claudin Expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-Q.; Wang, S.-B.; Wang, R.-G.; Zhang, W.; Wang, P.-L.; Su, X.-O. Phosphoproteome Analysis Reveals the Molecular Mechanisms Underlying Deoxynivalenol-Induced Intestinal Toxicity in IPEC-J2 Cells. Toxins 2016, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Beev, G.; Denev, S.; Bakalova, D. Zearalenone—Producing Activity of Fusarium Graminearum and Fusarium Oxysporum Isolated from Bulgarian Wheat. Bulg. J. Agric. Sci. 2013, 19, 255–259. [Google Scholar]
- Kumar, V.; Basu, M.S.; Rajendran, T.P. Mycotoxin Research and Mycoflora in Some Commercially Important Agricultural Commodities. Crop. Prot. 2008, 27, 891–905. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Tudor, D.S. Effects of Zearalenone and Its Derivatives on the Innate Immune Response of Swine. Toxicon 2010, 56, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Council for Agricultural Science and Technology. Mycotoxins: Risks in Plant, Animal, and Human Systems. Task Force Rep. 2003, 139, 1–199. [Google Scholar]
- Yazar, S.; Omurtag, G.Z. Fumonisins, Trichothecenes and Zearalenone in Cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Espín, S.; García-Fernández, A.-J.; Ruiz, M.-J. Estrogenic Activity of Zearalenone, α-Zearalenol and β-Zearalenol Assessed Using the E-Screen Assay in MCF-7 Cells. Toxicol. Mech. Methods 2018, 28, 239–242. [Google Scholar] [CrossRef]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the Immune Response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and Reproductive Function in Farm Animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Wu, W.; Pan, J.; Long, M. Detoxification Strategies for Zearalenone Using Microorganisms: A Review. Microorganisms 2019, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- IARC (International Agency For Research on Cancer). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins: Views and expert opinions of an IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, which met in Lyon, 9–16 June 1992. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1993; Volume 56, ISBN 978-92-832-1256-0. [Google Scholar]
- Olsen, M.; Pettersson, H.; Kiessling, K.-H. Reduction of Zearalenone to Zearalenol in Female Rat Liver by 3α-Hydroxysteroid Dehydrogenase. Acta Pharmacol. Toxicol. 1981, 48, 157–161. [Google Scholar] [CrossRef]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of Aflatoxin, Ochratoxin, Zearalenone, and Three Trichothecenes by Intact Rumen Fluid, Rumen Protozoa, and Rumen Bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debevere, S.; Cools, A.; Baere, S.D.; Haesaert, G.; Rychlik, M.; Croubels, S.; Fievez, V. In Vitro Rumen Simulations Show a Reduced Disappearance of Deoxynivalenol, Nivalenol and Enniatin B at Conditions of Rumen Acidosis and Lower Microbial Activity. Toxins 2020, 12, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeling, K.; Boguhn, J.; Strobel, E.; Dänicke, S.; Valenta, H.; Ueberschär, K.H.; Rodehutscord, M. On the Effects of Fusarium Toxin Contaminated Wheat and Wheat Chaff on Nutrient Utilisation and Turnover of Deoxynivalenol and Zearalenone in Vitro (Rusitec). Toxicol. In Vitro 2006, 20, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Dänicke, S.; Lebzien, P.; Valenta, H.; Ueberschär, K.H.; Flachowsky, G. On the Effects OfFusarium-Contaminated Wheat and the Feed Intake Level on Ruminal Fermentation and Toxin-Turnover of Cows. Mycotoxin Res. 2005, 21, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Dänicke, S.; Ueberschär, K.H.; Lebzien, P.; Flachowsky, G. On the Effects of Fusarium Toxin-Contaminated Wheat and the Feed Intake Level on the Metabolism and Carry over of Zearalenone in Dairy Cows. Food Addit. Contam 2005, 22, 847–855. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Feng, Y.-L.; Song, J.-L.; Zhou, X.-S. Zearalenone: A Mycotoxin With Different Toxic Effect in Domestic and Laboratory Animals’ Granulosa Cells. Front. Genet. 2018, 9, 667. [Google Scholar] [CrossRef] [Green Version]
- Shier, W.T.; Shier, A.C.; Xie, W.; Mirocha, C.J. Structure-Activity Relationships for Human Estrogenic Activity in Zearalenone Mycotoxins. Toxicon 2001, 39, 1435–1438. [Google Scholar] [CrossRef]
- Yang, C.; Song, G.; Lim, W. Effects of Mycotoxin-Contaminated Feed on Farm Animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef] [PubMed]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species Differences in the Hepatic Biotransformation of Zearalenone. Vet. J. 2006, 172, 96–102. [Google Scholar] [CrossRef]
- Kos, J.; Janić Hajnal, E.; Malachová, A.; Steiner, D.; Stranska, M.; Krska, R.; Poschmaier, B.; Sulyok, M. Mycotoxins in Maize Harvested in Republic of Serbia in the Period 2012–2015. Part 1: Regulated Mycotoxins and Its Derivatives. Food Chem. 2020, 312, 126034. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Berrada, H.; Miere, D.; Loghin, F.; Mañes, J. Study on Trichothecene and Zearalenone Presence in Romanian Wheat Relative to Weather Conditions. Toxins 2019, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golge, O.; Kabak, B. Occurrence of Deoxynivalenol and Zearalenone in Cereals and Cereal Products from Turkey. Food Control. 2020, 110, 106982. [Google Scholar] [CrossRef]
- Pante, G.C.; Silva, M.V.; Romoli, J.C.Z.; Rocha, G.H.O.; Bando, E.; Nerilo, S.B.; Mossini, S.A.G.; Machinski, M. Occurrence of Zearalenone in Corn Meal Commercialized in South Region of Brazil and Daily Intake Estimates in the Brazilian Population. J. Food Saf. 2019, 39, e12672. [Google Scholar] [CrossRef]
- Rai, A.; Dixit, S.; Singh, S.P.; Gautam, N.K.; Das, M.; Tripathi, A. Presence of Zearalenone in Cereal Grains and Its Exposure Risk Assessment in Indian Population. J. Food Sci. 2018, 83, 3126–3133. [Google Scholar] [CrossRef]
- Czembor, E.; Stępień, Ł.; Waśkiewicz, A. Effect of Environmental Factors on Fusarium Species and Associated Mycotoxins in Maize Grain Grown in Poland. PLoS ONE 2015, 10, e0133644. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Fang, H.; Zhao, Y.; Jin, Y.; Shen, J.; Zhou, C.; Zhou, Y.; Fu, Y.; Wang, J.; et al. Transcriptional Profiling of Zearalenone-Induced Inhibition of IPEC-J2 Cell Proliferation. Toxicon 2019, 172, 8–14. [Google Scholar] [CrossRef]
- Kouadio, J.H.; Mobio, T.A.; Baudrimont, I.; Moukha, S.; Dano, S.D.; Creppy, E.E. Comparative Study of Cytotoxicity and Oxidative Stress Induced by Deoxynivalenol, Zearalenone or Fumonisin B1 in Human Intestinal Cell Line Caco-2. Toxicology 2005, 213, 56–65. [Google Scholar] [CrossRef]
- Hassen, W.; Golli, E.E.; Baudrimont, I.; Mobio, A.T.; Ladjimi, M.M.; Creppy, E.E.; Bacha, H. Cytotoxicity and Hsp 70 Induction in Hep G2 Cells in Response to Zearalenone and Cytoprotection by Sub-Lethal Heat Shock. Toxicology 2005, 207, 293–301. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and Inflammatory Effects of Individual and Combined Exposure of HepG2 Cells to Zearalenone and Its Metabolites. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Abassi, H.; Ayed-Boussema, I.; Shirley, S.; Abid, S.; Bacha, H.; Micheau, O. The Mycotoxin Zearalenone Enhances Cell Proliferation, Colony Formation and Promotes Cell Migration in the Human Colon Carcinoma Cell Line HCT116. Toxicol. Lett. 2016, 254, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and Combined Cytotoxic Effects of Fusarium Toxins (Deoxynivalenol, Nivalenol, Zearalenone and Fumonisins B1) on Swine Jejunal Epithelial Cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef]
- Gao, F.; Jiang, L.; Chen, M.; Geng, C.; Yang, G.; Ji, F.; Zhong, L.; Liu, X. Genotoxic Effects Induced by Zearalenone in a Human Embryonic Kidney Cell Line. Mutat. Res. 2013, 755, 6–10. [Google Scholar] [CrossRef]
- Venkataramana, M.; Chandra Nayaka, S.; Anand, T.; Rajesh, R.; Aiyaz, M.; Divakara, S.T.; Murali, H.S.; Prakash, H.S.; Lakshmana Rao, P.V. Zearalenone Induced Toxicity in SHSY-5Y Cells: The Role of Oxidative Stress Evidenced by N-Acetyl Cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef]
- Hassen, W.; Ayed-Boussema, I.; Oscoz, A.A.; De Cerain Lopez, A.; Bacha, H. The Role of Oxidative Stress in Zearalenone-Mediated Toxicity in Hep G2 Cells: Oxidative DNA Damage, Gluthatione Depletion and Stress Proteins Induction. Toxicology 2007, 232, 294–302. [Google Scholar] [CrossRef]
- Fu, Y.; Jin, Y.; Zhao, Y.; Shan, A.; Fang, H.; Shen, J.; Zhou, C.; Yu, H.; Zhou, Y.F.; Wang, X.; et al. Zearalenone Induces Apoptosis in Bovine Mammary Epithelial Cells by Activating Endoplasmic Reticulum Stress. J. Dairy Sci. 2019, 102, 10543–10553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.-L.; Li, N.; Wang, H.; Ma, J.-M.; Shen, W.; Li, L. Zearalenone Exposure Induces the Apoptosis of Porcine Granulosa Cells and Changes Long Noncoding RNA Expression To Promote Antiapoptosis by Activating the JAK2–STAT3 Pathway. J. Agric. Food Chem. 2019, 67, 12117–12128. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Si, M.; Li, X.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. Zearalenone Induces Apoptosis of Rat Sertoli Cells through Fas-Fas Ligand and Mitochondrial Pathway. Enrivon. Toxicol. 2019, 34, 424–433. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Zhou, X.-Q.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Wang, S.-W.; Kuang, S.-Y.; Tang, L.; Feng, L. Effects of Dietary Zearalenone on Oxidative Stress, Cell Apoptosis, and Tight Junction in the Intestine of Juvenile Grass Carp (Ctenopharyngodon Idella). Toxins 2019, 11, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current Situation of Mycotoxin Contamination and Co-Occurrence in Animal Feed-Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.F.; Martins, H.M.; Costa, J.M.; Bernardo, F. Co-Occurrence of Deoxynivalenol and Zearalenone in Crops Marketed in Portugal. Food Addit. Contam. Part. B Surveill. 2008, 1, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, K.C.; Rocha, L.O.; Savi, G.D.; Carnielli-Queiroz, L.; Almeida, F.G.; Minella, E.; Corrêa, B. Occurrence of Deoxynivalenol and Zearalenone in Brewing Barley Grains from Brazil. Mycotoxin Res. 2018, 34, 173–178. [Google Scholar] [CrossRef]
- Calori-Domingues, M.A.; Bernardi, C.M.G.; Nardin, M.S.; de Souza, G.V.; dos Santos, F.G.R.; de Abreu Stein, M.; Da Gloria, E.M.; Dos Santos Dias, C.T.; De Camargo, A.C. Co-Occurrence and Distribution of Deoxynivalenol, Nivalenol and Zearalenone in Wheat from Brazil. Food Addit. Contam Part. B Surveill. 2016, 9, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Mwihia, E.W.; Lyche, J.L.; Mbuthia, P.G.; Ivanova, L.; Uhlig, S.; Gathumbi, J.K.; Maina, J.G.; Eshitera, E.E.; Eriksen, G.S. Co-Occurrence and Levels of Mycotoxins in Fish Feeds in Kenya. Toxins 2020, 12, 627. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B1, Deoxynivalenol and Zearalenone in Feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Chen, L.; Nüssler, A.K.; Liu, L.; Yang, W. Current Sights for Mechanisms of Deoxynivalenol-Induced Hepatotoxicity and Prospective Views for Future Scientific Research: A Mini Review: DON-Induced Hepatotoxicity and Prospective Views. J. Appl. Toxicol. 2017, 37, 518–529. [Google Scholar] [CrossRef]
- Smith, M.-C.; Hymery, N.; Troadec, S.; Pawtowski, A.; Coton, E.; Madec, S. Hepatotoxicity of Fusariotoxins, Alone and in Combination, towards the HepaRG Human Hepatocyte Cell Line. Food Chem. Toxicol. 2017, 109, 439–451. [Google Scholar] [CrossRef]
- Smith, M.-C.; Madec, S.; Pawtowski, A.; Coton, E.; Hymery, N. Individual and Combined Toxicological Effects of Deoxynivalenol and Zearalenone on Human Hepatocytes in in Vitro Chronic Exposure Conditions. Toxicol. Lett. 2017, 280, 238–246. [Google Scholar] [CrossRef]
- Smith, M.-C.; Timmins-Schiffman, E.; Coton, M.; Coton, E.; Hymery, N.; Nunn, B.L.; Madec, S. Differential Impacts of Individual and Combined Exposures of Deoxynivalenol and Zearalenone on the HepaRG Human Hepatic Cell Proteome. J. Proteom. 2018, 173, 89–98. [Google Scholar] [CrossRef]
- Gu, W.; Zhu, P.; Jiang, D.; He, X.; Li, Y.; Ji, J.; Zhang, L.; Sun, Y.; Sun, X. A Novel and Simple Cell-Based Electrochemical Impedance Biosensor for Evaluating the Combined Toxicity of DON and ZEN. Biosens. Bioelectron. 2015, 70, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhu, P.; Pi, F.; Zhang, Y.; Li, Y.; Wang, J.; Sun, X. Development of a Simple and Convenient Cell-Based Electrochemical Biosensor for Evaluating the Individual and Combined Toxicity of DON, ZEN, and AFB1. Biosens. Bioelectron. 2017, 97, 345–351. [Google Scholar] [CrossRef]
- Zhou, H.; George, S.; Hay, C.; Lee, J.; Qian, H.; Sun, X. Individual and Combined Effects of Aflatoxin B1, Deoxynivalenol and Zearalenone on HepG2 and RAW 264.7 Cell Lines. Food Chem. Toxicol. 2017, 103, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-H.; Lei, M.; Zhang, N.-Y.; Zhao, L.; Krumm, C.S.; Qi, D.-S. Hepatotoxic Effects of Mycotoxin Combinations in Mice. Food Chem. Toxicol. 2014, 74, 289–293. [Google Scholar] [CrossRef]
- Ji, J.; Wang, Q.; Wu, H.; Xia, S.; Guo, H.; Blaženović, I.; Zhang, Y.; Sun, X. Insights into Cellular Metabolic Pathways of the Combined Toxicity Responses of Caco-2 Cells Exposed to Deoxynivalenol, Zearalenone and Aflatoxin B1. Food Chem. Toxicol. 2019, 126, 106–112. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Sharaf el dein, O.; Hajlaoui, M.R.; Lemaire, C.; Bacha, H. In Vitro Investigation of Toxicological Interactions between the Fusariotoxins Deoxynivalenol and Zearalenone. Toxicon 2014, 84, 1–6. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, S.; Zhang, M.; Yang, L.; Cheng, B.; Li, J.; Shan, A. Individual and Combined Effects of Fusarium Toxins on Apoptosis in PK15 Cells and the Protective Role of N-Acetylcysteine. Food Chem. Toxicol. 2018, 111, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Deng, H.; Deng, Y.; Liang, Z.; Deng, J.; Zuo, Z.; Hu, Y.; Shen, L.; Yu, S.; Cao, S. Combined Effects of Deoxynivalenol and Zearalenone on Oxidative Injury and Apoptosis in Porcine Splenic Lymphocytes in Vitro. Exp. Toxicol. Pathol. 2017, 69, 612–617. [Google Scholar] [CrossRef]
- Kachlek, M.; Szabó-Fodor, J.; Blochné Bodnár, Z.; Horvatovich, K.; Kovács, M. Preliminary Results on the Interactive Effects of Deoxynivalenol, Zearalenone and Fumonisin B1 on Porcine Lymphocytes. Acta Vet. Hung. 2017, 65, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.-C.; Madec, S.; Troadec, S.; Coton, E.; Hymery, N. Effects of Fusariotoxin Co-Exposure on THP-1 Human Immune Cells. Cell Biol. Toxicol. 2018, 34, 191–205. [Google Scholar] [CrossRef]
- Ji, J.; Zhu, P.; Pi, F.; Sun, C.; Jiang, H.; Sun, J.; Wang, X.; Zhang, Y.; Sun, X. GC-TOF/MS-Based Metabolomic Strategy for Combined Toxicity Effects of Deoxynivalenol and Zearalenone on Murine Macrophage ANA-1 Cells. Toxicon 2016, 120, 175–184. [Google Scholar] [CrossRef]
- Zhou, H.; George, S.; Li, C.; Gurusamy, S.; Sun, X.; Gong, Z.; Qian, H. Combined Toxicity of Prevalent Mycotoxins Studied in Fish Cell Line and Zebrafish Larvae Revealed That Type of Interactions Is Dose-Dependent. Aquat. Toxicol. 2017, 193, 60–71. [Google Scholar] [CrossRef]
- Smith, M.-C.; Gheux, A.; Coton, M.; Madec, S.; Hymery, N.; Coton, E. In Vitro Co-Culture Models to Evaluate Acute Cytotoxicity of Individual and Combined Mycotoxin Exposures on Caco-2, THP-1 and HepaRG Human Cell Lines. Chem. Biol. Interact. 2018, 281, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Ren, Z.; Gao, S.; Chen, Y.; Yang, Y.; Yang, D.; Deng, J.; Zuo, Z.; Wang, Y.; Shen, L. Individual and Combined Effects of Deoxynivalenol and Zearalenone on Mouse Kidney. Environ. Toxicol. Pharmacol. 2015, 40, 686–691. [Google Scholar] [CrossRef]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Bázár, G.; Kocsó, D.; Ali, O.; Kovács, M. Individual and Combined Effects of Fumonisin B1, Deoxynivalenol and Zearalenone on the Hepatic and Renal Membrane Lipid Integrity of Rats. Toxins 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, G.; Sun, K.; Xia, S.; Feng, Z.; Zou, H.; Gu, J.; Yuan, Y.; Zhu, J.; Liu, Z.; Bian, J. Decrease in Immune Function and the Role of Mitogen-Activated Protein Kinase (MAPK) Overactivation in Apoptosis during T Lymphocytes Activation Induced by Zearalenone, Deoxynivalenol, and Their Combinations. Chemosphere 2020, 255, 126999. [Google Scholar] [CrossRef]
- Szabó-Fodor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Bóta, B.; Kachlek, M.; Mézes, M.; Balogh, K.; Kövér, G.; Nagy, I.; et al. Interaction between the Three Frequently Co-Occurring Fusarium Mycotoxins in Rats. J. Anim. Physiol. Anim. Nutr. 2019, 103, 370–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Zhu, P.; Cui, F.; Pi, F.; Zhang, Y.; Li, Y.; Wang, J.; Sun, X. The Antagonistic Effect of Mycotoxins Deoxynivalenol and Zearalenone on Metabolic Profiling in Serum and Liver of Mice. Toxins 2017, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Zhu, P.; Blaženović, I.; Cui, F.; Gholami, M.; Sun, J.; Habimana, J.; Zhang, Y.; Sun, X. Explaining Combinatorial Effects of Mycotoxins Deoxynivalenol and Zearalenone in Mice with Urinary Metabolomic Profiling. Sci. Rep. 2018, 8, 3762. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.H.; Deng, H.D.; Deng, Y.T.; Deng, J.L.; Zuo, Z.C.; Yu, S.M.; Shen, L.H.; Cui, H.M.; Xu, Z.W.; Hu, Y.C. Effect of the Fusarium Toxins, Zearalenone and Deoxynivalenol, on the Mouse Brain. Environ. Toxicol. Pharmacol. 2016, 46, 62–70. [Google Scholar] [CrossRef]
- Jia, R.; Liu, W.; Zhao, L.; Cao, L.; Shen, Z. Low Doses of Individual and Combined Deoxynivalenol and Zearalenone in Naturally Moldy Diets Impair Intestinal Functions via Inducing Inflammation and Disrupting Epithelial Barrier in the Intestine of Piglets. Toxicol. Lett. 2020, 333, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Motiu, M.; Taranu, I. Food Contaminant Zearalenone and Its Metabolites Affect Cytokine Synthesis and Intestinal Epithelial Integrity of Porcine Cells. Toxins 2015, 7, 1979–1988. [Google Scholar] [CrossRef]
- Pinton, P.; Oswald, I.P. Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Oswald, I.P. The Effects of Mycotoxins, Fungal Food Contaminants, on the Intestinal Epithelial Cell-Derived Innate Immune Response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, T.Q.; Wu, J.; Zhang, X.L.; Xu, Y.Y.; Qian, Y.Z. Multi-Parameter Analysis of Combined Hepatotoxicity Induced by Mycotoxin Mixtures in HepG2 Cells. World Mycotoxin J. 2018, 11, 225–235. [Google Scholar] [CrossRef]

{kind=link}
| Mycotoxin | Fungal Species | Human Health Effects |
|---|---|---|
| Aflatoxins | Aspergillus | Haemorrhage, liver damage. carcinogenesis, gastrointestinal dysfunction, anaemia, jaundice, reduced reproductivity |
| Trichothecenes | Fusarium | Growth stunt, reproductive disorder, vomiting, feed refusal, reduced ovarian function |
| Ochratoxins | Aspergillus, Penicillium | Carcinogenesis, nephrotoxicity |
| Zearalenone | Fusarium | Hormonal imbalance, oestrogenic effects |
| Fumonisins | Fusarium | Nephrotoxicity, esophageal cancer |
| Foodstuff | Maximum Level (µg/kg) |
| Unprocessed cereals except durum wheat, oats and maize | 1250 |
| Unprocessed durum wheat and oats | 1750 |
| Unprocessed maize except for unprocessed maize intended to be processed by wet milling | 1750 |
| Cereals intended for direct human consumption, cereal flour, bran and germ as end product marketed for direct human consumption | 750 |
| Dry pasta | 750 |
| Bread, pastries, biscuits, cereal snacks and breakfast cereal | 500 |
| Processed cereal based foods and baby foods for infants and young children | 200 |
| Products Intended for Animal Feed | Guidance Value Relative to a Feedingstuff with a Moisture Content of 12% (µg/kg) |
| Cereals and cereal products with the exception of maize by-products | 8000 |
| Maize by-products | 12,000 |
| Complementary and complete feedingstuff (with the exception of those listed below) | 5000 |
| Complementary and complete feedingstuff for pigs | 900 |
| Complementary and complete feedingstuff for calves (<4 months), lambs, and kids | 2000 |
| Foodstuff | Maximum Level (µg/kg) |
| Unprocessed cereals other than maize | 100 |
| Unprocessed maize except for unprocessed maize intended to be processed by wet milling | 350 |
| Cereals intended for direct human consumption, cereal flour, bran, and germ as the end product marketed for direct human consumption | 75 |
| Bread (including small bakery wares), pastries, biscuits, cereal snacks, and breakfast cereals, excluding maize-based snacks and maize-based breakfast cereals | 50 |
| Maize intended for direct human consumption, maize-based snacks and maize-based breakfast cereals | 100 |
| Processed cereal based foods (excluding processed maize-based foods) and baby foods for infants and young children | 20 |
| Processed maize-based foods for infants and young children | 20 |
| Products Intended for Animal Feed | Guidance Value Relative to a Feedingstuff with a Moisture Content of 12% (µg/kg) |
| Cereals and cereal products with the exception of maize by-products | 2000 |
| Maize by-products | 3000 |
| Complementary and complete feedingstuff for piglets and gilts (young sows) | 100 |
| Complementary and complete feedingstuff for sows and fattening pigs | 250 |
| Complementary and complete feedingstuff for calves, dairy cattle, sheep (including lamb) and goats (including kids) | 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thapa, A.; Horgan, K.A.; White, B.; Walls, D. Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants? Toxins 2021, 13, 561. https://doi.org/10.3390/toxins13080561
Thapa A, Horgan KA, White B, Walls D. Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants? Toxins. 2021; 13(8):561. https://doi.org/10.3390/toxins13080561
Chicago/Turabian StyleThapa, Asmita, Karina A. Horgan, Blánaid White, and Dermot Walls. 2021. "Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants?" Toxins 13, no. 8: 561. https://doi.org/10.3390/toxins13080561
APA StyleThapa, A., Horgan, K. A., White, B., & Walls, D. (2021). Deoxynivalenol and Zearalenone—Synergistic or Antagonistic Agri-Food Chain Co-Contaminants? Toxins, 13(8), 561. https://doi.org/10.3390/toxins13080561