Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days

 , ,
, ,  , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
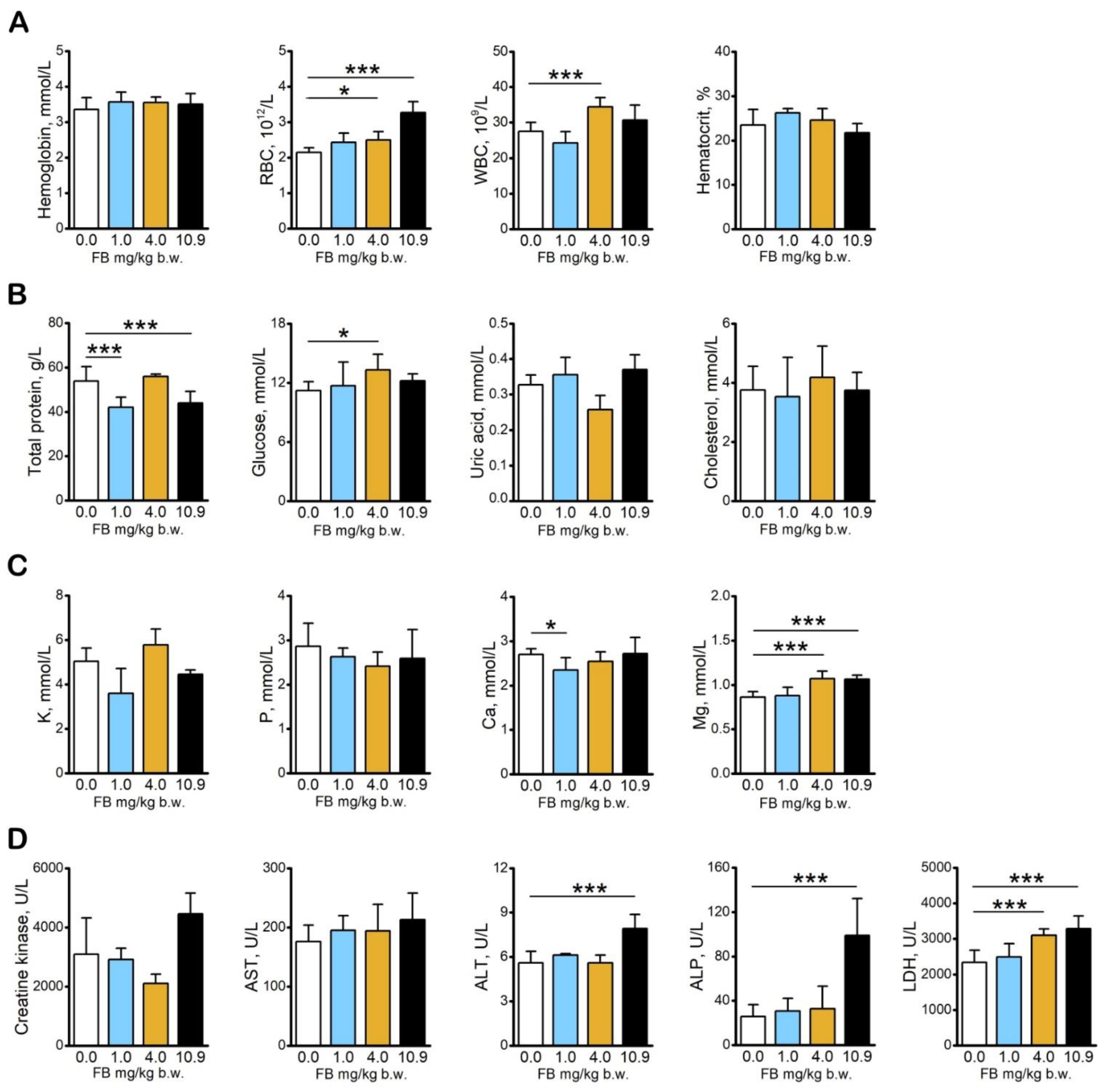
2.1. Basal Blood Morphology and Blood Serum Biochemical Analysis
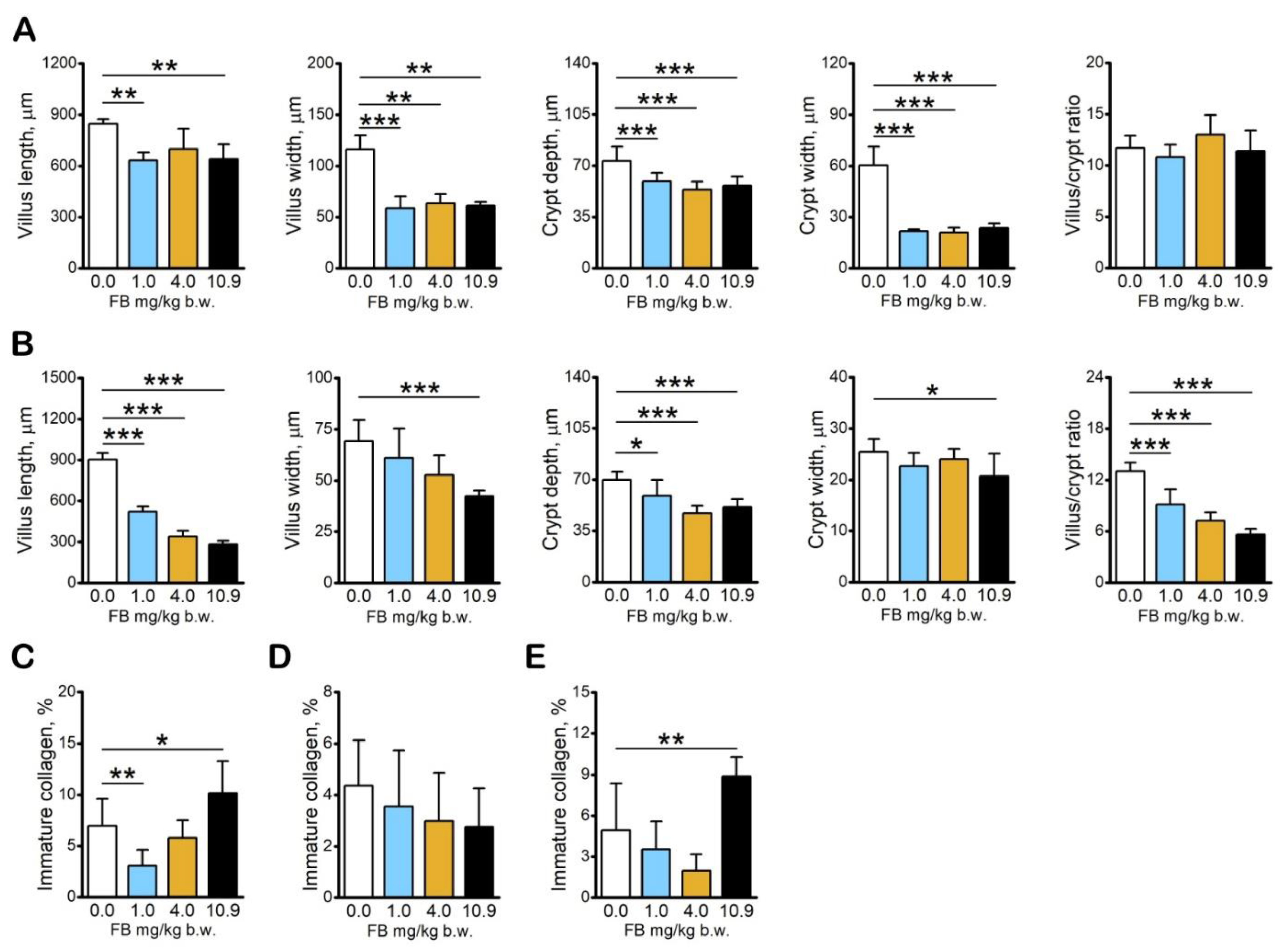
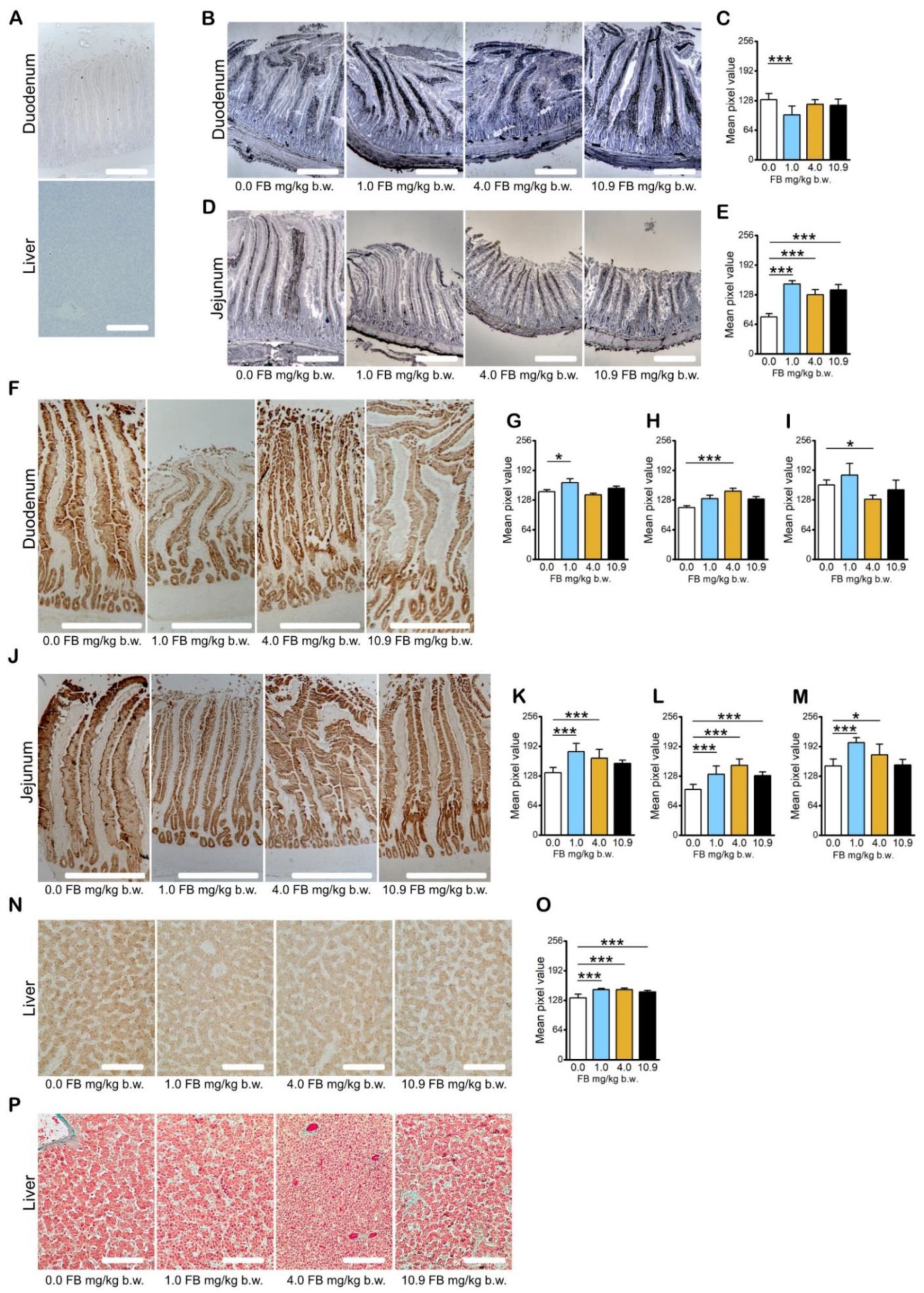
2.2. Results of Intestine and Liver Analyses
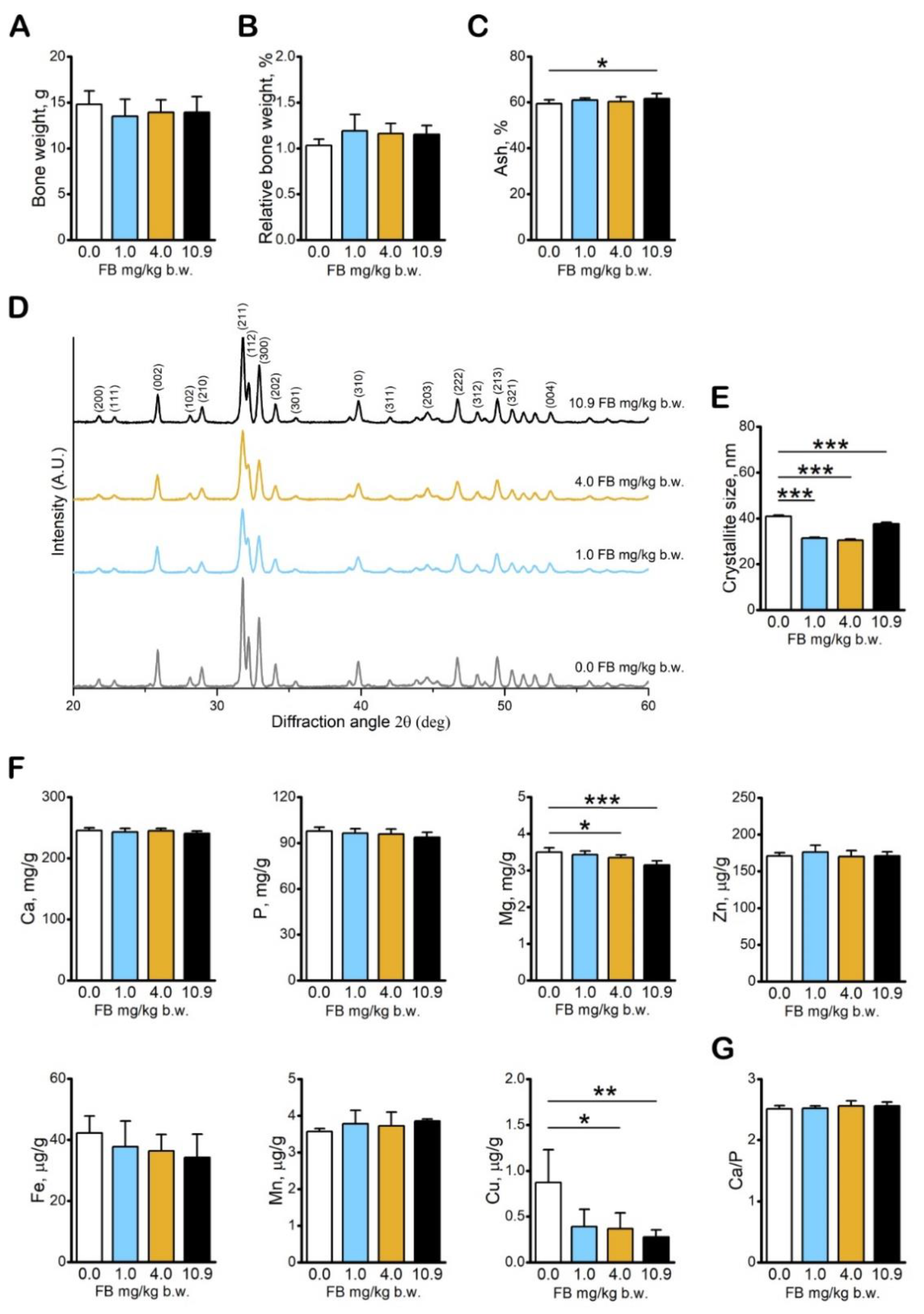
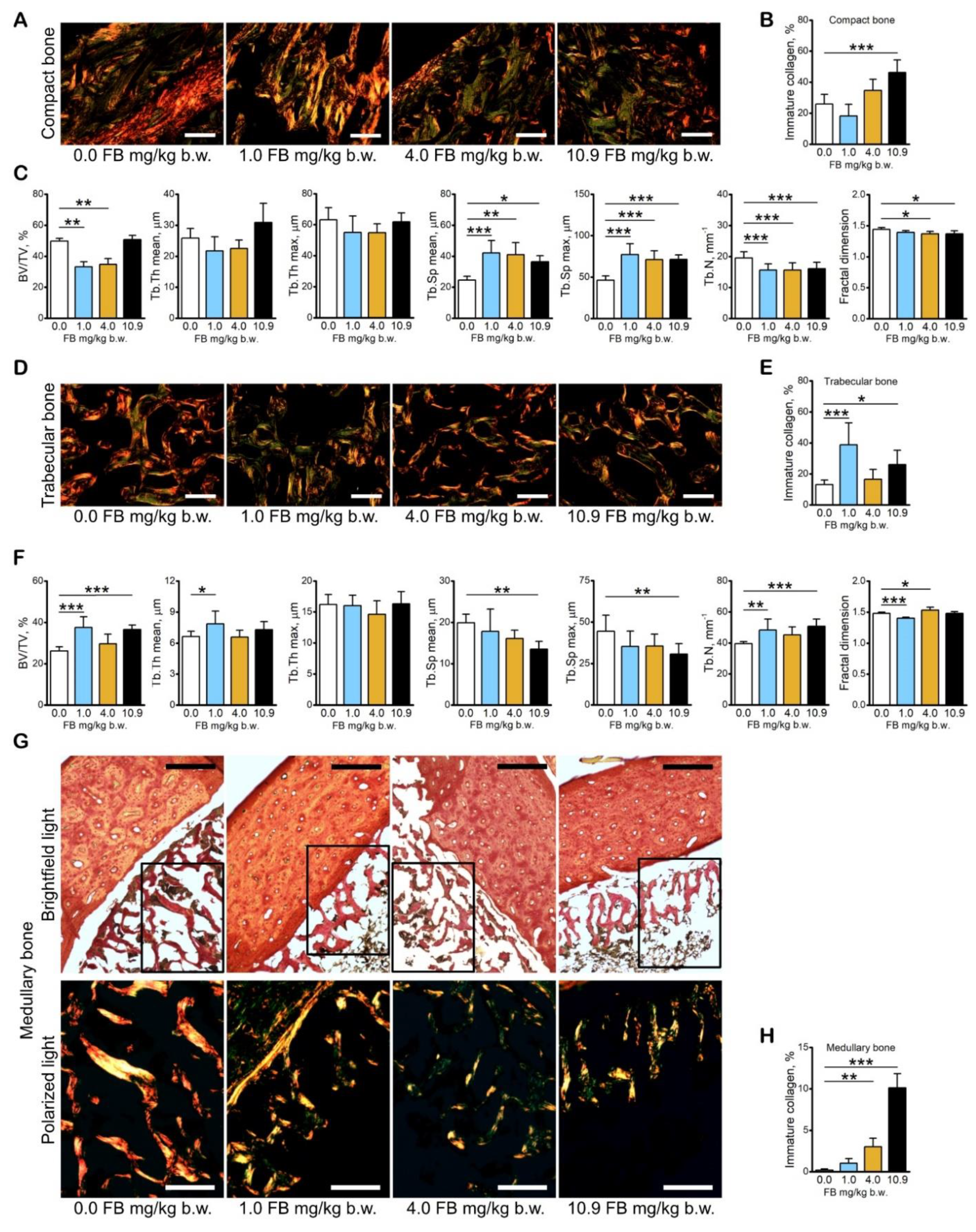
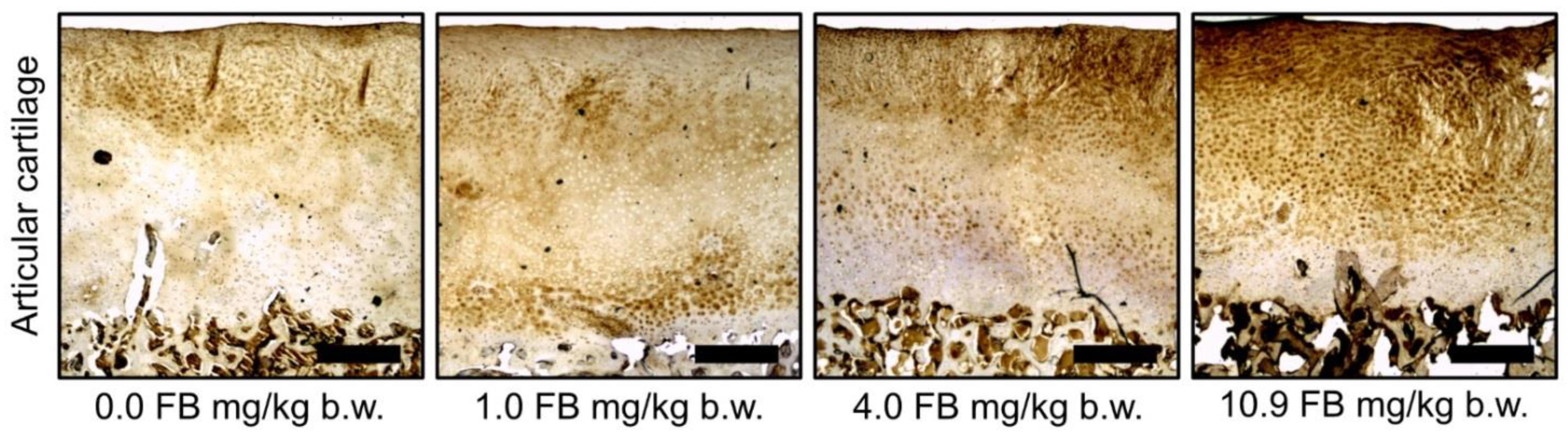
2.3. Results of Bone Analyses
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fumonisins
5.2. Animals, Experimental Procedures, and Tissue Collection
5.3. Basal Blood Morphology and Blood Serum Analyses
5.4. Analyses of Intestine and Liver Samples
5.5. Analyses of Bone Samples
5.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef]
- European Commission. Recommendation EU 2006/576 of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union L 2006, 229, 7–9. [Google Scholar]
- EFSA (European Food Safety Authority). Panel on Contaminants in the Food Chain (CONTAM). Scientific opinion on the appropriateness to set a group health-based guidance value for fumonisins and their modified forms. EFSA J. 2018, 16, 5172. [Google Scholar]
- EFSA (European Food Safety Authority). Panel on Contaminants in the Food Chain (CONTAM). Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018, 16, 5242. [Google Scholar]
- Riley, R.T.; Voss, K.A. Differential sensitivity of rat kidney and liver to fumonisin toxicity: Organ-specific differences in toxin accumulation and sphingoid base metabolism. Toxicol. Sci. 2006, 92, 335–345. [Google Scholar] [CrossRef]
- Rudyk, H.; Tomaszewska, E.; Kotsyumbas, I.; Muszyński, S.; Tomczyk-Warunek, A.; Szymańczyk, S.; Dobrowolski, P.; Wiącek, D.; Kamiński, D.; Brezvyn, O. Bone homeostasis in experimental fumonisins intoxication of rats. Ann. Anim. Sci. 2019, 18, 403–419. [Google Scholar] [CrossRef]
- Antonissen, G.; Devreese, M.; Van Immerseel, F.; Baere, S.D.; Hessenberger, S.; Martel, A.; Croubels, S. Chronic exposure to deoxynivalenol has no influence on the oral bioavailability of fumonisin B-1 in broiler chickens. Toxins 2015, 7, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; Backer, P.D.; Croubels, S. Overview of the most important mycotoxins for the pig and poultry husbandry. Vlaams Diergen. Tijds. 2013, 82, 171–180. [Google Scholar] [CrossRef]
- Jain, N.C. Essential of Veterinary Hematology; Lea & Febiger: Philadelphia, PA, USA, 1993. [Google Scholar]
- Gyenis, J.; Suto, Z.; Romvari, R.; Horn, P. Tracking the development of serum biochemical parameters in two laying hen strains—A comparative study. Arch. Tierz. 2006, 49, 593–606. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmay, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmay, G.; Sulyok, M.; Rudolf, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize—An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Muszyński, S.; Dobrowolski, P.; Kwiecień, M.; Klebaniuk, R.; Szymańczyk, S.; Tomczyk, A.; Kowalik, S.; Milczarek, A.; Świetlicka, I. The influence of dietary replacement of soybean meal with high-tannin faba beans on gut-bone axis and metabolic response in broiler chickens. Ann. Anim. Sci. 2018, 18, 801–824. [Google Scholar] [CrossRef]
- Rudyk, H.; Tomaszewska, E.; Arciszewski, M.B.; Muszyński, S.; Tomczyk-Warunek, A.; Dobrowolski, P.; Donaldson, J.; Brezvyn, O.; Kotsyumbas, I. Histomorphometrical changes in intestine structure and innervation following experimental fumonisins intoxication in male Wistar rats. Pol. J. Vet. Sci. 2020, 23, 77–88. [Google Scholar] [PubMed]
- Kubena, L.F.; Harvey, R.B.; Buckley, S.A.; Bailey, R.H.; Rottinghaus, G.E. Effects of long-term feeding of diets containing moniliformin, supplied by Fusarium fujikuroi culture material, and fumonisin, supplied by Fusarium moniliforme culture material, to laying hens. Poult. Sci. 1999, 78, 1499–1505. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Brown, T.P.; Weibking, T.S.; Rottinghaus, G.E. Fumonisin toxicity in broiler chicks. J. Vet. Diagn. Invest. 1992, 4, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Griss, L.G.; Fortuoso, B.F.; Silva, A.D.; Fracasso, M.; Lopes, T.L.; Schetinger, M.R.S.; Gundel, S.; Ourique, A.F.; Carneiro, C.; et al. Feed contaminated by fumonisin (Fusarium spp.) in chicks has a negative influence on oxidative stress and performance, and the inclusion of curcumin-loaded nanocapsules minimizes these effects. Microb. Pathog. 2020, 148, 104496. [Google Scholar] [CrossRef]
- Xu, L.; Eicher, S.D.; Applegate, T.J. Effects of increasing dietary concentrations of corn naturally contaminated with deoxynivalenol on broiler and turkey poultry performance and response to lipopolysaccharide. Poult. Sci. 2011, 90, 2766–2774. [Google Scholar] [CrossRef]
- Tessari, E.N.C.; Kobashigawa, E.; Cardoso, A.L.S.P.; Ledoux, D.R.; Rottinghaus, G.E.; Oliveira, C.A.F. Effects of aflatoxin B1 and fumonisin B1 on blood biochemical parameters in broilers. Toxins 2010, 2, 453–460. [Google Scholar] [CrossRef]
- Oliveira, E.M.; Tanure, C.B.G.S.; Castejon, F.V.; Castro, R.M.A.D.; Rocha, F.R.T.; Carvalho, F.B.; Andrade, M.A.; Stringhini, J.H. Performance and nutrient metabolizability in broilers fed diets containing corn contaminated with fumonisin B1 and esterified glucomannan. Rev. Bras. Cienc. Avic. 2015, 17, 313–318. [Google Scholar] [CrossRef]
- Dazuk, V.; Boiago, M.M.; Rolim, G.; Paravisi, A.; Copetti, P.M.; Bissacotti, B.F.; Morsch, V.M.; Vedovatto, M.; Gazoni, F.L.; Matte, F.; et al. Laying hens fed mycotoxin-contaminated feed produced by Fusarium fungi (T-2 toxin and fumonisin B1) and Saccharomyces cerevisiae lysate: Impacts on poultry health, productive efficiency, and egg quality. Microb. Pathog. 2020, 149, 104517. [Google Scholar] [CrossRef]
- Henry, M.H.; Wyatt, R.D.; Fletcher, O.J. The toxicity of purified fumonisin B1 in broiler chicks. Poult. Sci. 2000, 79, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Ledoux, D.R.; Bermudez, A.J.; Fristsche, K.L.; Rottinghaus, G.E. Effects of fumonisin B1 on selected immune responses in broiler chicks. Poult. Sci. 1999, 78, 1275–1282. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Li, Z.; Wang, G.Y.; Yang, Z.B.; Yang, W.R.; Zhang, G.G.; Wu, Y.B. Effects of Fusarium mycotoxins with yeast cell wall absorbent on hematology, serum biochemistry, and oxidative stress in broiler chickens. J. Appl. Poult. Res. 2014, 23, 165–173. [Google Scholar] [CrossRef]
- Espada, Y.; Gopegui, R.R.D.; Cuadradas, C.; Cabarnes, F.J. Fumonisin mycotoxicosis in broilers. Weights and serum chemistry modifications. Avian Dis. 1994, 38, 454–460. [Google Scholar] [CrossRef]
- Bermudez, A.J.; Ledoux, D.R.; Rottinghaus, G.E.; Bennett, G.A. The individual and combined effects of the Fusarium mycotoxins moniliformin and fumonisin B1 in turkeys. Avian Dis. 1997, 41, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Siloto, E.V.; Oliveira, E.F.A.; Sartori, J.R.; Fascina, V.B.; Martins, B.A.B.; Ledoux, D.R.; Rottinghaus, G.E.; Sartori, D.R.S. Lipid metabolism of commercial layers fed diets containing aflatoxin, fumonisin, and a binder. Poult. Sci. 2013, 92, 2077–2083. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.R.; Smith, T.K. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on performance and metabolism of laying hens. Poult. Sci. 2004, 83, 1849–1856. [Google Scholar] [CrossRef]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef]
- Lee, S.; Kim, D.-H.; Keum, M.-C.; Han, E.; An, B.-K.; Chang, H.-H.; Choi, Y.-H.; Moon, B.-H.; Lee, K.-W. Effects of fumonisin B1 and mycotoxin binders on growth performance, tibia characteristics, gut physiology, and stress indicators in broiler chickens raised in different stocking densities. Poult. Sci. 2018, 97, 845–854. [Google Scholar] [CrossRef]
- Shastak, Y.; Rodehuyscord, M. A review of the role of magnesium in poultry nutrition. Worlds Poult. Sci. J. 2015, 71, 125–138. [Google Scholar] [CrossRef]
- Torres, C.A.; Korver, D.R. Influences of trace mineral nutrition and maternal flock age on broiler embryo bone development. Poult. Sci. 2018, 97, 2996–3003. [Google Scholar] [CrossRef]
- Muszyński, S.; Tomaszewska, E.; Kwiecień, M.; Dobrowolski, P.; Tomczyk, A. Effect of dietary phytase supplementation on bone and hyaline cartilage development of broilers fed with organically complexed copper in a Cu-deficient diet. Biol. Trace Elem. Res. 2018, 182, 339–353. [Google Scholar] [CrossRef]
- Kierończyk, B.; Rawski, M.; Józefiak, D.; Świątkiewicz, S. Infectious and non-infectious factors associated with leg disorders in poultry—A review. Ann. Anim. Sci. 2017, 17, 645–666. [Google Scholar] [CrossRef]
- Rath, N.C.; Huff, G.R.; Huff, W.E.; Balog, J.M. Factors regulating bone maturity and strength in poultry. Poult. Sci. 2000, 79, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, G.; Liu, T. The effect of fumonisin B1 on isolated chondrocytes and on bone formation. Poult. Sci. 1995, 74, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Harrar, K.; Hamami, L. The fractal dimension correlated to the bone mineral density. WSEAS Trans. Signal. Process. 2008, 4, 110–126. [Google Scholar]
- Chen, Q.; Bao, N.; Yao, Q.; Li, Z.Y. Fractal dimension: A complementary diagnostic indicator of osteoporosis to bone mineral density. Med. Hypotheses 2018, 116, 136–138. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Świątkiewicz, S.; Arczewska-Włosek, A.; Wojtysiak, D.; Dobrowolski, P.; Domaradzki, P.; Świetlicka, I.; Donaldson, J.; Hułas-Stasiak, M.; Muszyński, S. Alpha-ketoglutarate: An effective feed supplement in improving bone metabolism and muscle quality of laying hens: A preliminary study. Animals 2020, 10, 2420. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, S.; Arczewska, M.; Świątkiewicz, S.; Arczewska-Włosek, A.; Dobrowolski, P.; Świetlicka, I.; Hułas-Stasiak, M.; Blicharski, T.; Donaldson, J.; Schwarz, T.; et al. The effect of dietary rye inclusion and xylanase supplementation on structural organization of bone constitutive phases in laying hens fed a wheat-corn diet. Animals 2020, 10, 2010. [Google Scholar] [CrossRef]
- Ruppel, M.E.; Miller, L.M.; Burr, D.B. The effect of the microscopic and nanoscale structure on bone fragility. Osteoporos. Int. 2008, 19, 1251–1265. [Google Scholar] [CrossRef]
- Shahnazari, M.; Lang, D.; Fosmire, G.; Sharkey, N.; Mitchell, A.; Leach, R. Strontium administration in young chickens improves bone volume and architecture but does not enhance bone structural and material strength. Calcif. Tissue Int. 2007, 80, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, P.; Rai, D.; Garg, M.L. Zinc improves the bone mechanical strength in ovariectomized rat model by restoring bone composition and hydroxyapatite crystallite dimension. Vitam. Miner. 2016, 5, 137. [Google Scholar] [CrossRef]
- Muszyński, S.; Dobrowolski, P.; Kasperek, K.; Knaga, S.; Kwiecień, M.; Donaldson, J.; Kutyła, M.; Kapica, M.; Tomaszewska, E. Effects of yeast (Saccharomyces cerevisiae) probiotics supplementation on bone quality characteristics in young Japanese quail (Coturnix japonica): The role of sex on the action of the gut-bone axis. Animals 2020, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.K.; Baere, S.D.; Mountzouris, K.C.; Su, S.; Wong, E.A.; Meulenaer, B.D.; et al. The mycotoxins deoxynivalenol and fumonisins alter the extrinsic component of intestinal barrier in broiler chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.P.; Rottinghaus, G.E.; Williams, M.E. Fumonisin mycotoxicosis in broilers: Performance and pathology. Avian Dis. 1992, 36, 450–454. [Google Scholar] [CrossRef]
- Bianchi, M.D.; Oliveira, C.A.F.; Albuquerque, R.; Guerra, J.L.; Correa, B. Effects of prolonged oral administration of aflatoxin B1 and fumonisin B1 in broiler chickens. Poult. Sci. 2005, 84, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Poersch, A.B.; Trombetta, F.; Braga, A.C.M.; Boeira, S.P.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S.; et al. Involvement of oxidative stress in subacute toxicity induced by fumonisin B1 in broiler chicks. Vet. Microbiol. 2014, 174, 180–185. [Google Scholar] [CrossRef]
- Robert, H.; Payros, D.; Pinton, P.; Théodorou, V.; Mercier-Bonin, M.; Oswald, I.P. Impact of mycotoxins on the intestine: Are mucus and microbiota new targets? J. Toxicol. Environ. Health B Crit. Rev. 2017, 20, 249–275. [Google Scholar] [CrossRef]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef]
- Hodgson, E.; Roe, R.M.; Mailman, R.B.; Chambers, J.E. (Eds.) Dictionary of Toxicology, 3rd ed.; Academic Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Smulikowska, S.; Rutkowski, A. Recommended Allowances and Nutritive Value of Feedstuffs. In Poultry Feeding Standards, 5th ed.; The Kielanowski Institute of Animal Physiology and Nutrition PAS: Jabłonna, Poland, 2018; pp. 66–74. (In Polish) [Google Scholar]
- Vudathala, D.K.; Prelusky, D.B.; Ayroud, M.; Trenholm, H.L.; Miller, J.D. Pharmacokinetic fate and pathological effects of 14C-fumonins B1 in laying hens. Nat. Toxins 1994, 2, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Hułas-Stasiak, M.; Jakubowicz-Gil, J.; Dobrowolski, P.; Grzesiak, M.; Muszyński, S.; Świątkiewicz, M.; Tomaszewska, E. Regulation of folliculogenesis by growth factors in piglet ovary exposed prenatally to β-hydroxy-β-methylbutyrate (HMB). Ann. Anim. Sci. 2020, 20, 899–917. [Google Scholar] [CrossRef]
- Puzio, I.; Muszyński, S.; Dobrowolski, P.; Kapica, M.; Pawłowska-Olszewska, M.; Donaldson, J.; Tomaszewska, E. Alterations in small intestine and liver morphology, immunolocalisation of leptin, ghrelin and nesfatin-1 as well as immunoexpression of tight junction proteins in intestinal mucosa after gastrectomy in rat model. J. Clin. Med. 2021, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Rich, L.; Whittaker, P. Collagen and picrosirius red staining: A polarized light assessment of fibrillar hue and spatial distribution. J. Morphol. Sci. 2017, 22, 97–104. [Google Scholar]
- Tomaszewska, E.; Dobrowolski, P.; Puzio, I.; Donaldson, J.; Muszyński, S. Acrylamide-induced prenatal programming of bone structure in mammal model. Ann. Anim. Sci. 2020, 20, 1257–1287. [Google Scholar] [CrossRef]
- Scherrer, P. Bestimmung der inneren Struktur und der größe von kolloidteilchen mittels röntgenstrahlen. Göttingen Nachr. 1918, 2, 98–100. [Google Scholar]
- Muszyński, S.; Tomaszewska, E.; Dobrowolski, P.; Kwiecień, M.; Wiącek, D.; Świetlicka, I.; Skibińska, M.; Szymańska-Chargot, M.; Orzeł, J.; Świetlicki, M.; et al. Analysis of bone osteometry, mineralization, mechanical and histomorphometrical properties of tibiotarsus in broiler chickens demonstrates a influence of dietary chickpea seeds (Cicer arietinum L.) inclusion as a primary protein source. PLoS ONE 2018, 13, e0208921. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, M.A. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska, E.; Rudyk, H.; Dobrowolski, P.; Donaldson, J.; Świetlicka, I.; Puzio, I.; Kamiński, D.; Wiącek, D.; Kushnir, V.; Brezvyn, O.; et al. Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days. Toxins 2021, 13, 375. https://doi.org/10.3390/toxins13060375
Tomaszewska E, Rudyk H, Dobrowolski P, Donaldson J, Świetlicka I, Puzio I, Kamiński D, Wiącek D, Kushnir V, Brezvyn O, et al. Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days. Toxins. 2021; 13(6):375. https://doi.org/10.3390/toxins13060375
Chicago/Turabian StyleTomaszewska, Ewa, Halyna Rudyk, Piotr Dobrowolski, Janine Donaldson, Izabela Świetlicka, Iwona Puzio, Daniel Kamiński, Dariusz Wiącek, Volodymyr Kushnir, Oksana Brezvyn, and et al. 2021. "Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days" Toxins 13, no. 6: 375. https://doi.org/10.3390/toxins13060375
APA StyleTomaszewska, E., Rudyk, H., Dobrowolski, P., Donaldson, J., Świetlicka, I., Puzio, I., Kamiński, D., Wiącek, D., Kushnir, V., Brezvyn, O., Muzyka, V., Doraczyńska, R., Muszyński, S., & Kotsyumbas, I. (2021). Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days. Toxins, 13(6), 375. https://doi.org/10.3390/toxins13060375