Two Different Inoculation Methods Unveiled the Relative Independence of DON Accumulation in Wheat Kernels from Disease Severity on Spike after Infection by Fusarium Head Blight


Abstract
1. Introduction
2. Results
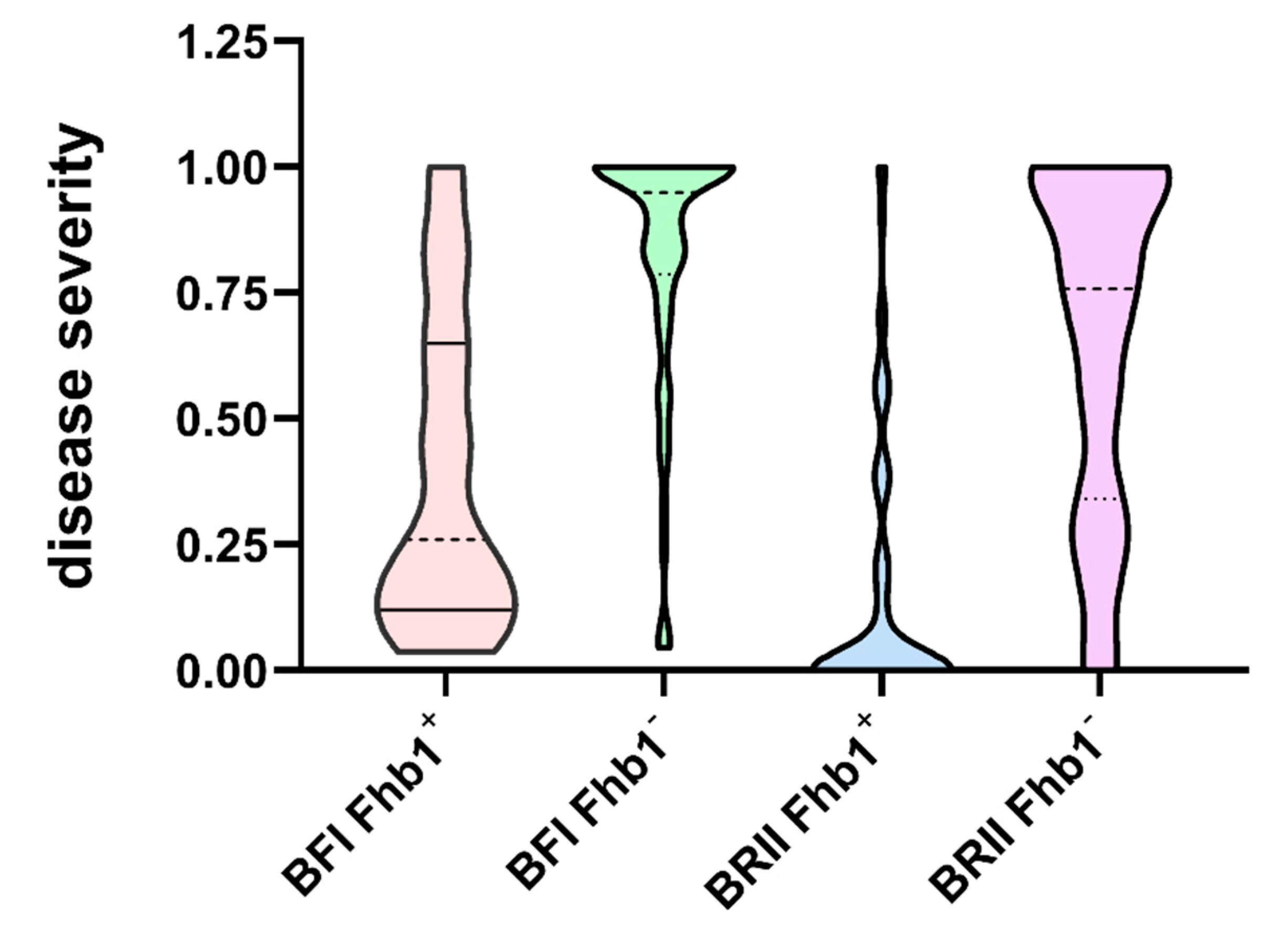
2.1. The BFI and BRII Methods Produced Differential Patterns of Disease Severity
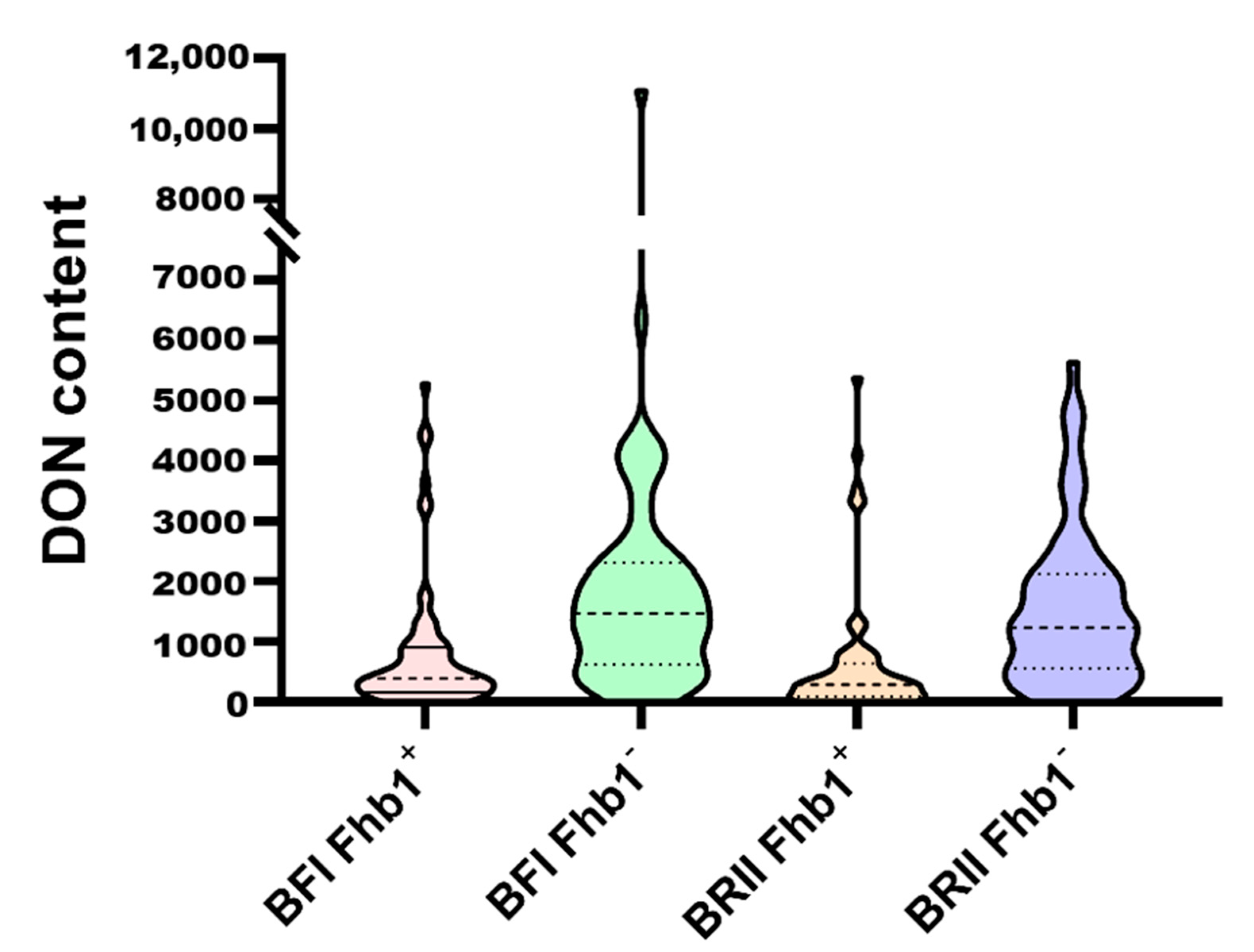
2.2. The BFI and BRII Methods Generated Similar Patterns of DON Content in Kernels
2.3. Low Disease Severity Does Not Mean Low DON Content
2.4. Evaluation of Overall Resistance and Selection of LSLD Lines
3. Discussion
3.1. Complex Relationship between Disease Severity and DON Concentration in Kernels
3.2. Different Inoculation Methods Did Not Alter the Effects of Fhb1 on PSS and DON
3.3. No Correlations among 15A-DON, D3G and DON
3.4. Overall Resistance to FHB Should Be More Useful and Reasonable in Wheat Breeding
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Fusarium graminearum Strain, Inoculum Preparation and FHB Inoculation
5.3. Phenotypic Evaluation
5.4. Determination of DON Contents and Its Derivative Forms
5.5. Classification of Disease Severity and DON
5.6. Evaluation of Overall Resistance to FHB
5.7. DNA Extraction and Fhb1 Genotyping
5.8. Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bai, G.; Shaner, G. management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Champeil, A.; Doré, T.; Fourbet, J. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant. Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Mesterhazy, A. Types and components of resistance to Fusarium head blight of wheat. Plant. Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health Part B 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Kouadio, J.H.; Mobio, T.A.; Baudrimont, I.; Moukha, S.; Dano, S.D.; Creppy, E.E. Comparative study of cytotoxicity and oxidative stress induced by deoxynivalenol, zearalenone or fumonisin B1 in human intestinal cell line Caco-2. Toxicology 2005, 213, 56–65. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Shi, J.; Liu, X.; Qiu, J.; Ji, F.; Xu, J.; Dong, F.; Yin, X.; Ran, J. Deoxynivalenol contamination in wheat and its management. Sci. Agric. Sin. 2014, 47, 3641–3654. [Google Scholar]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A Unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant. Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.; Tan, L.; Chin, L.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Cui, L.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Zhou, L.; Liu, Y. A minor survey of deoxynivalenol in Fusarium infected wheat from Yangtze–Huaihe river basin region in China. Food Control 2013, 30, 469–473. [Google Scholar] [CrossRef]
- Boutigny, A.-L.; Richard-Forget, F.; Barreau, C. Natural mechanisms for cereal resistance to the accumulation of Fusarium trichothecenes. Eur. J. Plant. Pathol. 2008, 121, 411–423. [Google Scholar] [CrossRef]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked Mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Schwartz, H.; Krska, R.; Moll, W.-D.; Knasmüller, S.; Ritzmann, M.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in rats. Toxicol. Lett. 2012, 213, 367–373. [Google Scholar] [CrossRef]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Hennig-Pauka, I.; Moll, W.-D.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef]
- Tan, Y.; Zhu, R.; Wu, A.; Liu, N. Major types of masked mycotoxins and state-of-the-art methodological advance for their detection. Sci. Sin. Chim. 2016, 46, 251–256. [Google Scholar] [CrossRef]
- Bai, G.; Kolb, F.L.; Shaner, G.; Domier, L.L. Amplified fragment length polymorphism markers linked to a major quantitative trait locus controlling scab resistance in wheat. Phytopathology 1999, 89, 343–348. [Google Scholar] [CrossRef]
- Waldron, B.L.; Moreno-Sevilla, B.; Anderson, J.A.; Stack, R.W.; Frohberg, R.C. RFLP Mapping of QTL for Fusarium head blight resistance in wheat. Crop. Sci. 1999, 39, 805–811. [Google Scholar] [CrossRef]
- Bai, G.; Plattner, R.; Desjardins, A.; Kolb, F.; McIntosh, R.A. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant. Breed. 2001, 120, 1–6. [Google Scholar] [CrossRef]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Ákos, M.; Krska, R.; et al. The ability to detoxify the Mycotoxin Deoxynivalenol colocalizes with a major quantitative trait locus for fusarium head blight resistance in wheat. Mol. Plant. Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Geddes, J.; Eudes, F.; Tucker, J.R.; Legge, W.G.; Selinger, L. Evaluation of inoculation methods on infection and deoxynivalenol production by Fusarium graminearumon barley. Can. J. Plant. Pathol. 2008, 30, 66–73. [Google Scholar] [CrossRef]
- Mesterházy, A.; Lehoczki-Krsjak, S.; Varga, M.; Szabó-Hevér, Á.; Tóth, B.; Lemmens, M. Breeding for FHB resistance via fusarium damaged kernels and deoxynivalenol accumulation as well as inoculation methods in winter wheat. Agric. Sci. 2015, 6, 970–1002. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Li, M.A.; Zhu, M.S.; Li, L.; He, X.; Sun, Z.; Li, T. Basal Rachis Internode Injection (BRII): A novel inoculation method to evaluate wheat resistance to Fusarium head blight. Phytopathology 2021. [Google Scholar] [CrossRef]
- Jayatilake, D.V.; Bai, G.H.; Dong, Y.H. A novel quantitative trait locus for Fusarium head blight resistance in chromosome 7A of wheat. Theor. Appl. Genet. 2011, 122, 1189–1198. [Google Scholar] [CrossRef]
- Yu, J.-B.; Bai, G.-H.; Cai, S.-B.; Dong, Y.-H.; Ban, T. New Fusarium head blight-resistant sources from asian wheat germplasm. Crop. Sci. 2008, 48, 1090–1097. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Bartók, T.; Mirocha, C.G.; Komoróczy, R. Nature of wheat resistance to Fusarium head blight and the role of deoxynivalenol for breeding. Plant. Breed. 1999, 118, 97–110. [Google Scholar] [CrossRef]
- Ma, H.X.; Zhang, K.M.; Gao, L.; Bai, G.H.; Chen, H.G.; Cai, Z.X.; Lu, W.Z. Quantitative trait loci for resistance to Fusarium head blight and deoxynivalenol accumulation in Wangshuibai wheat under field conditions. Plant. Pathol. 2006, 55, 739–745. [Google Scholar] [CrossRef]
- Kang, Z.; Buchenauer, H. Immunocytochemical localization of fusarium toxins in infected wheat spikes by Fusarium culmorum. Physiol. Mol. Plant. Pathol. 1999, 55, 275–288. [Google Scholar] [CrossRef]
- Kang, Z.; Buchenauer, H. Ultrastructural and immunocytochemical investigation of pathogen development and host responses in resistant and susceptible wheat spikes infected by Fusarium culmorum. Physiol. Mol. Plant. Pathol. 2000, 57, 255–268. [Google Scholar] [CrossRef]
- Liu, S.; Hall, M.D.; Griffey, C.A.; McKendry, A.L. Meta-Analysis of QTL associated with Fusarium head blight resistance in wheat. Crop. Sci. 2009, 49, 1955–1968. [Google Scholar] [CrossRef]
- Zheng, T.; Hua, C.; Li, L.; Sun, Z.; Yuan, M.; Bai, G.; Humphreys, G.; Li, T. Integration of meta-QTL discovery with omics: Towards a molecular breeding platform for improving wheat resistance to Fusarium head blight. Crop. J. 2020. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Steiner, B.; Hartl, L.; Griesser, M.; Angerer, N.; Lengauer, D.; Miedaner, T.; Schneider, B.; Lemmens, M. Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. II. Resistance to fungal penetration and spread. Theor. Appl. Genet. 2003, 107, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Steiner, B.; Vautrin, S.; Nussbaumer, T.; Siegwart, G.; Zamini, M.; Jungreithmeier, F.; Gratl, V.; Lemmens, M.; Mayer, K.F.X.; et al. Suppressed recombination and unique candidate genes in the divergent haplotype encoding Fhb1, a major Fusarium head blight resistance locus in wheat. Theor. Appl. Genet. 2016, 129, 1607–1623. [Google Scholar] [CrossRef]
- Li, G.; Zhou, J.; Jia, H.; Gao, Z.; Fan, M.; Luo, Y.; Zhao, P.; Xue, S.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]
- Rawat, N.; Pumphrey, M.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.; et al. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef]
- De Loubresse, N.G.; Prokhorova, I.V.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nat. Cell Biol. 2014, 513, 517–522. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Dall’Erta, A.; Mantovani, P.; Massi, A.; Galaverna, G. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat. World Mycotoxin J. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Audenaert, K.; De Boevre, M.; Vanheule, A.; Callewaert, J.; Bekaert, B.; Höfte, M.; De Saeger, S.; Haesaert, G. Mycotoxin glucosylation in commercial wheat varieties: Impact on resistance to Fusarium graminearum under laboratory and field conditions. Food Control 2013, 34, 756–762. [Google Scholar] [CrossRef]
- Yang, X.; Pan, Y.; Singh, P.K.; He, X.; Ren, Y.; Zhao, L.; Zhang, N.; Cheng, S.; Chen, F. Investigation and genome-wide association study for Fusarium crown rot resistance in Chinese common wheat. BMC Plant. Biol. 2019, 19, 153. [Google Scholar] [CrossRef] [PubMed]
- Farber, O.; Kadmon, R. Assessment of alternative approaches for bioclimatic modeling with special emphasis on the Mahalanobis distance. Ecol. Model. 2003, 160, 115–130. [Google Scholar] [CrossRef]
- Yu, J.-B.; Bai, G.-H.; Cai, S.-B.; Ban, T. Marker-assisted characterization of Asian wheat lines for resistance to Fusarium head blight. Theor. Appl. Genet. 2006, 113, 308–320. [Google Scholar] [CrossRef]
- Li, T.; Luo, M.; Zhang, D.; Wu, D.; Li, L.; Bai, G. Effective marker alleles associated with type 2 resistance to Fusarium head blight infection in fields. Breed. Sci. 2016, 66, 350–357. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mao, X.; Hua, C.; Yang, L.; Zhang, Y.; Sun, Z.; Li, L.; Li, T. The effects of selenium on wheat Fusarium head blight and DON accumulation were selenium compound-dependent. Toxins 2020, 12, 573. [Google Scholar] [CrossRef]
- McLachlan, G.J. Mahalanobis distance. Resonance 1999, 4, 20–26. [Google Scholar] [CrossRef]
- Bernardo, A.N.; Ma, H.; Zhang, D.; Bai, G. Single nucleotide polymorphism in wheat chromosome region harboring Fhb1 for Fusarium head blight resistance. Mol. Breed. 2012, 29, 477–488. [Google Scholar] [CrossRef]
- Su, Z.; Jin, S.; Zhang, D.; Bai, G. Development and validation of diagnostic markers for Fhb1 region, a major QTL for Fusarium head blight resistance in wheat. Theor. Appl. Genet. 2018, 131, 2371–2380. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Methods | Fhb1 | Min | Max | Mean Value | Allelic Difference | DF | SD | T Value | Sig. |
|---|---|---|---|---|---|---|---|---|---|---|
| PSS | BFI | Yes | 0.04 | 1.00 | 0.38 | 0.43 | 70 | 0.40 | −8.98 | p < 0.01 |
| No | 0 | 1.00 | 0.81 | |||||||
| BRII | Yes | 0 | 1.00 | 0.16 | 0.50 | 70 | 0.44 | −9.47 | p < 0.01 | |
| No | 0 | 1.00 | 0.66 | |||||||
| DON (μg·kg−1) | BFI | Yes | 0 | 5281.60 | 787.60 | 987.67 | 70 | 1979.22 | −4.21 | p < 0.01 |
| No | 0 | 11081.80 | 1775.26 | |||||||
| BRII | Yes | 0 | 5373.17 | 554.03 | 916.59 | 70 | 1465.02 | −5.20 | p < 0.01 | |
| No | 0 | 5630.47 | 1470.61 |
| BRII-PSS | BFI-DON | BRII-DON | |
|---|---|---|---|
| BFI-PSS | 0.633 ** | 0.427 ** | 0.386 ** |
| BRII-PSS | 0.331 ** | 0.336 ** | |
| BFI-DON | 0.455 ** |
| PSS | DON | |||
|---|---|---|---|---|
| BFI | BRII | BFI | BRII | |
| Min | 0.04 | 0.00 | 0.00 | 0.00 |
| 25% | 0.21 | 0.00 | 326.96 | 212.84 |
| 50% | 0.67 | 0.33 | 883.31 | 620.44 |
| 75% | 1.00 | 0.84 | 1722.30 | 1378.63 |
| Max | 1.00 | 1.00 | 11081.75 | 5630.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Hua, C.; Hu, Y.; Li, L.; Sun, Z.; Li, T. Two Different Inoculation Methods Unveiled the Relative Independence of DON Accumulation in Wheat Kernels from Disease Severity on Spike after Infection by Fusarium Head Blight. Toxins 2021, 13, 353. https://doi.org/10.3390/toxins13050353
Wang R, Hua C, Hu Y, Li L, Sun Z, Li T. Two Different Inoculation Methods Unveiled the Relative Independence of DON Accumulation in Wheat Kernels from Disease Severity on Spike after Infection by Fusarium Head Blight. Toxins. 2021; 13(5):353. https://doi.org/10.3390/toxins13050353
Chicago/Turabian StyleWang, Rong, Chen Hua, Yi Hu, Lei Li, Zhengxi Sun, and Tao Li. 2021. "Two Different Inoculation Methods Unveiled the Relative Independence of DON Accumulation in Wheat Kernels from Disease Severity on Spike after Infection by Fusarium Head Blight" Toxins 13, no. 5: 353. https://doi.org/10.3390/toxins13050353
APA StyleWang, R., Hua, C., Hu, Y., Li, L., Sun, Z., & Li, T. (2021). Two Different Inoculation Methods Unveiled the Relative Independence of DON Accumulation in Wheat Kernels from Disease Severity on Spike after Infection by Fusarium Head Blight. Toxins, 13(5), 353. https://doi.org/10.3390/toxins13050353