The Administration of the Synbiotic Lactobacillus bulgaricus 6c3 Strain, Inulin and Fructooligosaccharide Decreases the Concentrations of Indoxyl Sulfate and Kidney Damage in a Rat Model

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Isolation of Strains Capable to Reduce IS In Vitro
2.2. Identification of the Selected Strains
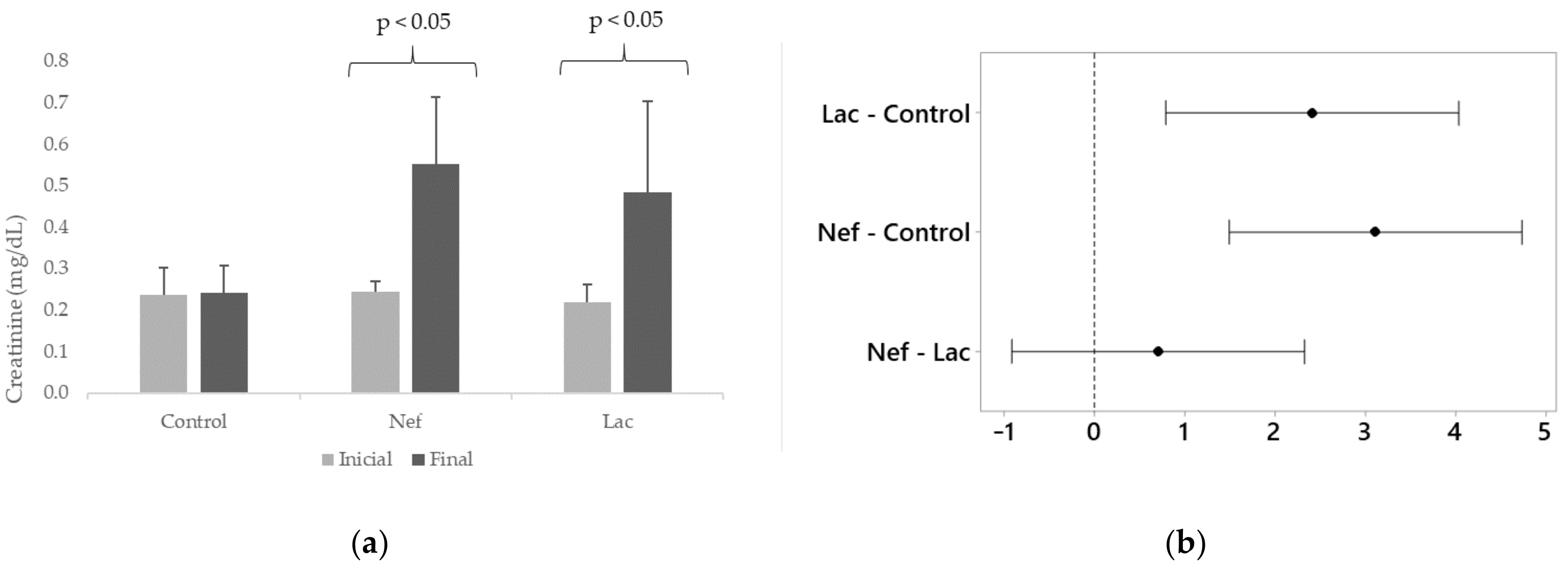
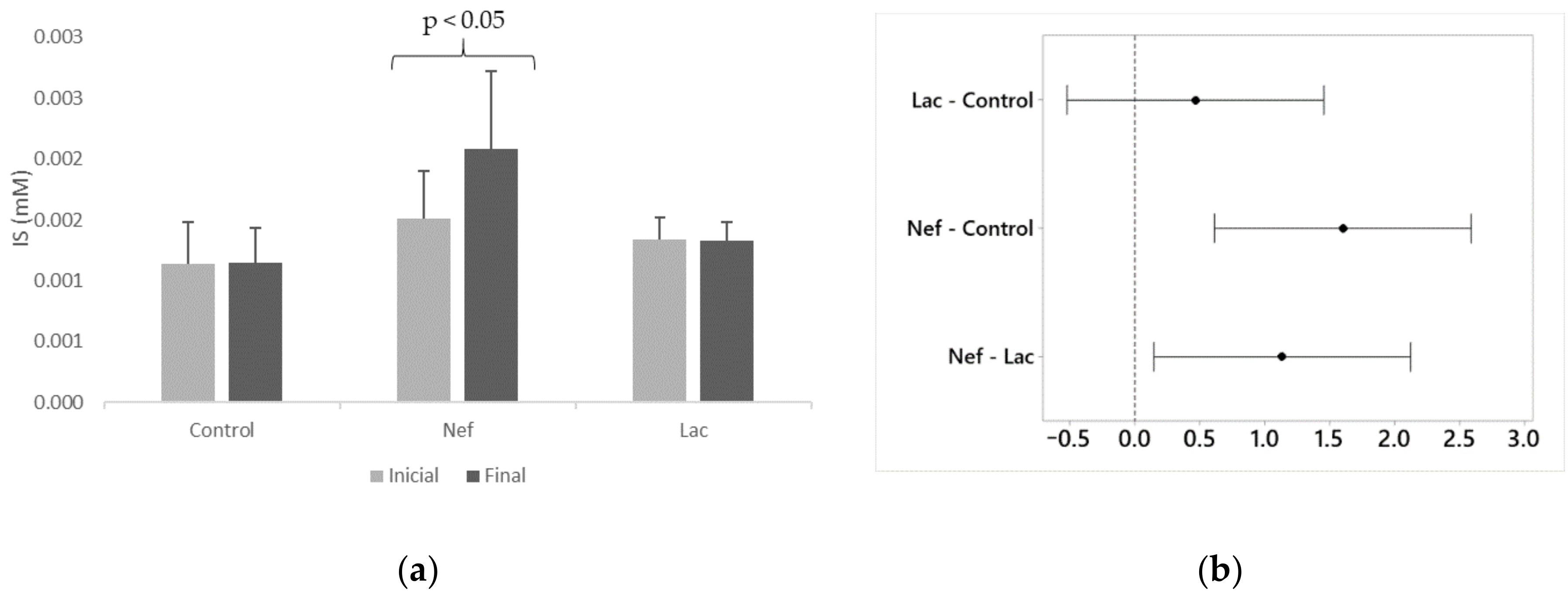
2.3. Effect of the Synbiotic in Blood Creatinine and IS Levels
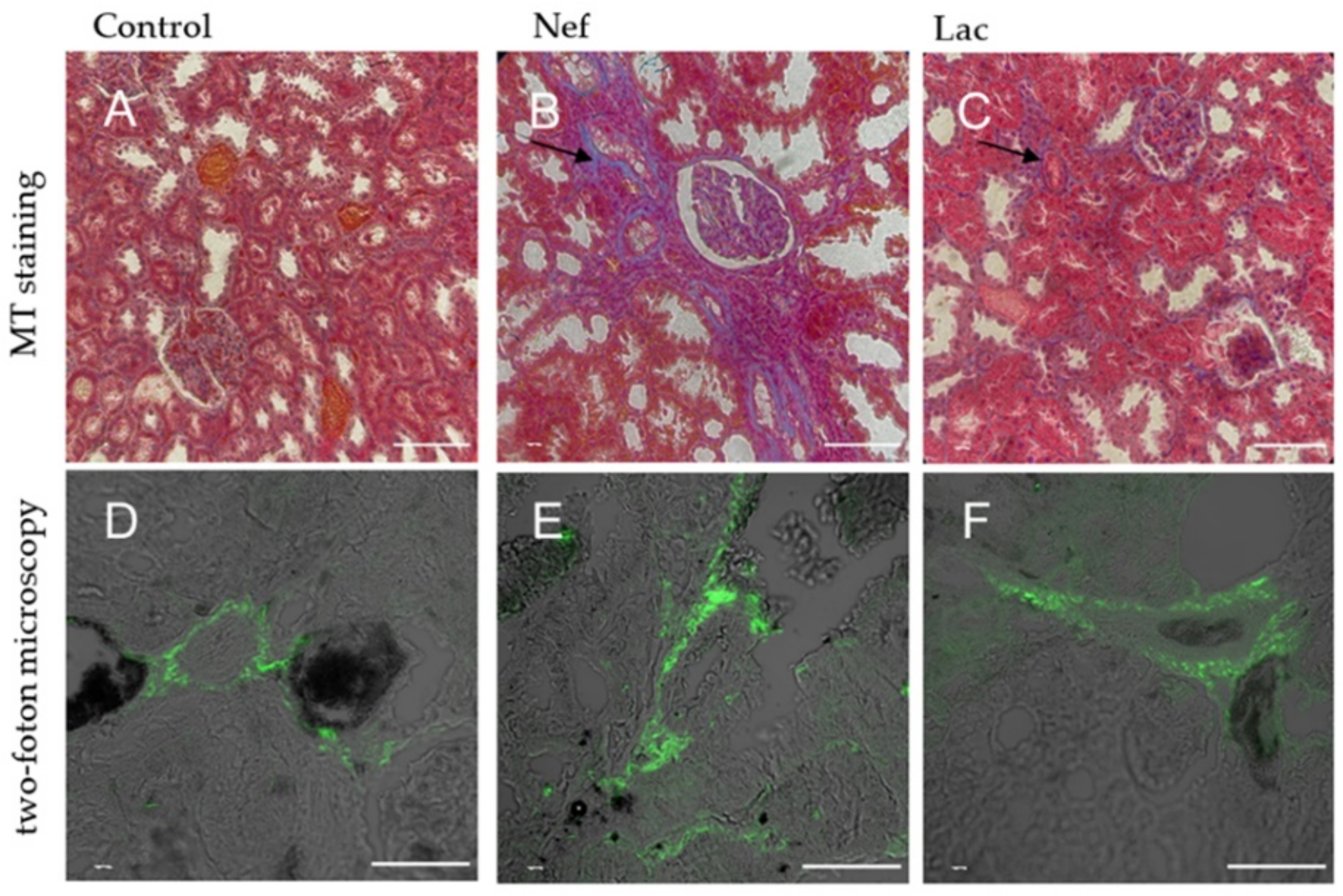
2.4. The Histological Analysis Showed that the Administration of the Sybiotic Decreased the Progression of CKD
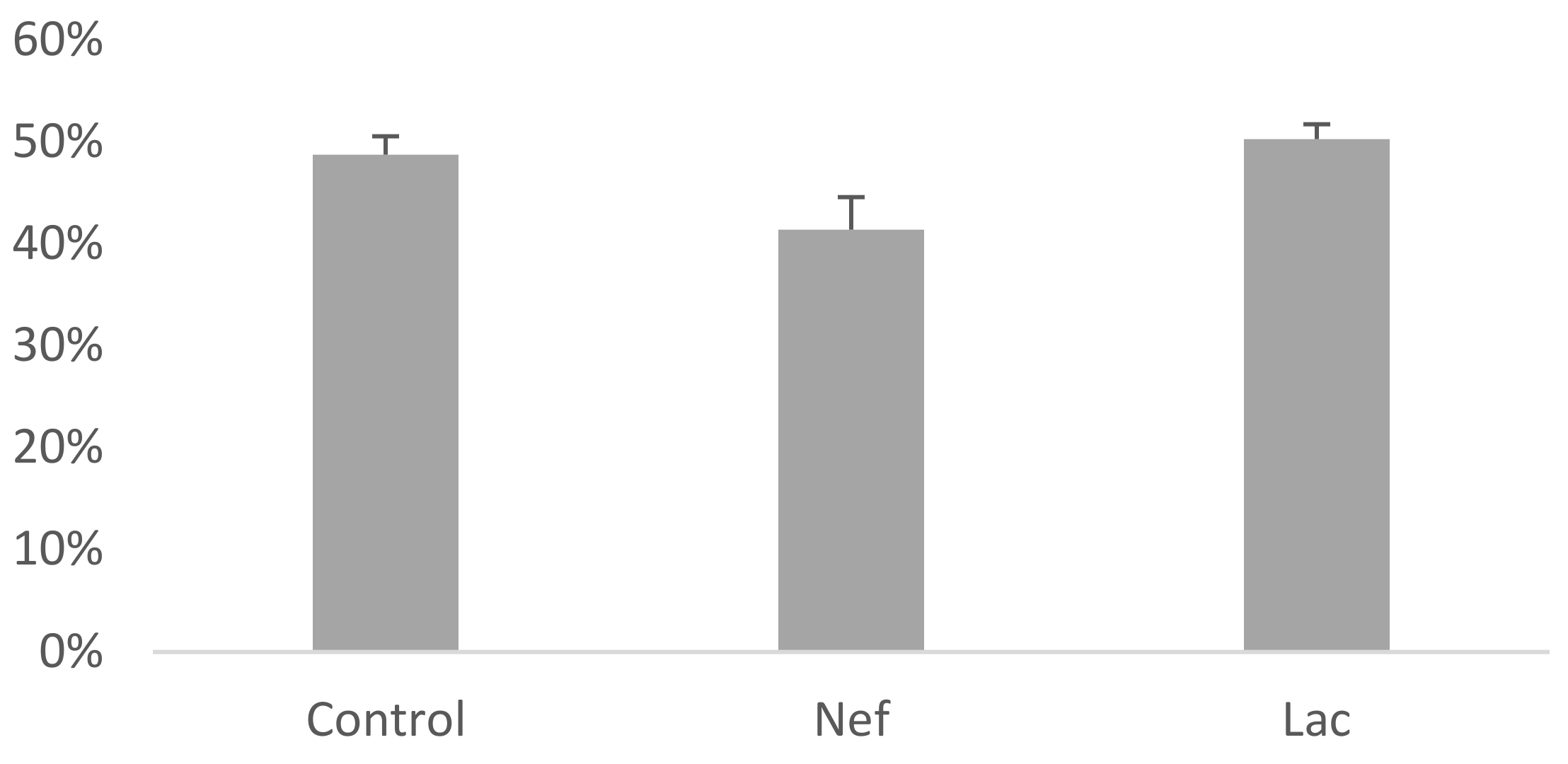
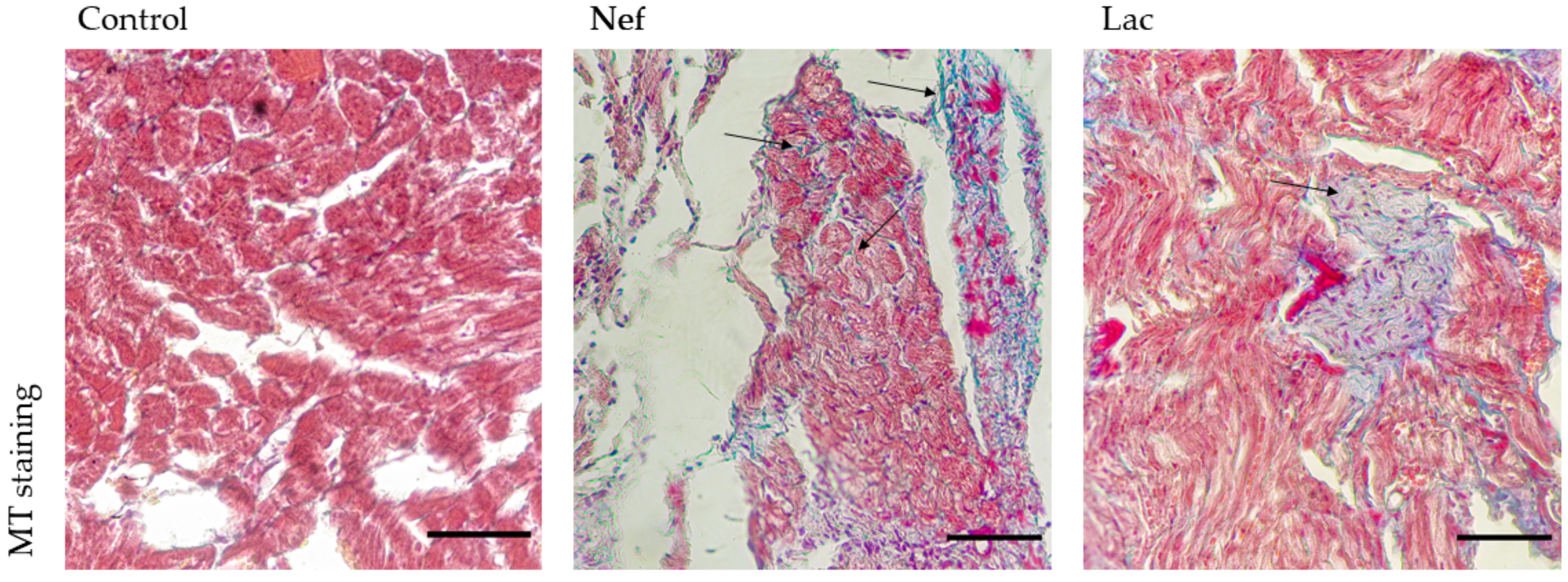
2.5. The Administration of the Synbiotic Did Not Attenuate Cardic Fibrosis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Isolation
5.2. Determintion, by HPLC-DAD, of Strains Capable to Reduce IS
5.3. DNA Amplification and Identification of the Strains
5.4. Effect of the Administration of the Synbiotic to Nephrectomized Sprague-Dawley Rats
5.5. Assesment of Creatinine and IS Concentration
5.6. Histological Analysis
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoccali, C.; Vanholder, R.; Massy, Z.A.; Ortiz, A.; Sarafidis, P.; Dekker, F.W.; Fliser, D.; Fouque, D.; Heine, G.H.; Jager, K.J.; et al. Cardiovascular Medicine Working Group of the European Renal Association—European Dialysis Transplantation, A. The systemic nature of CKD. Nat. Rev. Nephrol. 2017, 13, 344–358. [Google Scholar] [CrossRef]
- Romagnani, P.; Remuzzi, G.; Glassock, R.; Levin, A.; Jager, K.J.; Tonelli, M.; Massy, Z.; Wanner, C.; Anders, H.J. Chronic kidney disease. Nat. Rev. Dis. Primers. 2017, 3, 17088. [Google Scholar] [CrossRef]
- CDC. Chronic Kidney Disease in the United States. 2019. Available online: Availabe online: https://www.cdc.gov/kidneydisease/pdf/2019_National-Chronic-Kidney-Disease-Fact-Sheet.pdf (accessed on 8 April 2020).
- Moradi, H.; Sica, D.A.; Kalantar-Zadeh, K. Cardiovascular burden associated with uremic toxins in patients with chronic kidney disease. Am. J. Nephrol. 2013, 38, 136–148. [Google Scholar] [CrossRef]
- Meijers, B.; Evenepoel, P.; Anders, H.J. Intestinal microbiome and fitness in kidney disease. Nat. Rev. Nephrol. 2019, 15, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; DeSantis, T.Z.; Ni, Z.; Nguyen, T.H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Renal Physiol. 2016, 310, F857–F871. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Microbiota and metabolites in metabolic diseases. Nat. Rev. Endocrinol. 2019, 15, 69–70. [Google Scholar] [CrossRef]
- Yoshifuji, A.; Wakino, S.; Irie, J.; Matsui, A.; Hasegawa, K.; Tokuyama, H.; Hayashi, K.; Itoh, H. Oral adsorbent AST-120 ameliorates gut environment and protects against the progression of renal impairment in CKD rats. Clin. Exp. Nephrol. 2018, 22, 1069–1078. [Google Scholar] [CrossRef]
- Ranganathan, N.; Vyas, U.; Hanlon, K.; Ranganathan, P.; Irvin, A. Improvements in Glomerular Filtration Rate (GFR) in Chronic Kidney Disease (CKD) Patients Using a Commercial Patented and Proprietary Probiotic Prebiotic Formulation* -3rd Biennial Survey. Int. J. Nephrol. Kidney Fail 2018, 4, 1–9. [Google Scholar]
- Tao, S.; Tao, S.; Cheng, Y.; Liu, J.; Ma, L.; Fu, P. Effects of probiotic supplements on the progression of chronic kidney disease: A meta-analysis. Nephrology 2019, 24, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Sircana, A.; De Michieli, F.; Parente, R.; Framarin, L.; Leone, N.; Berrutti, M.; Paschetta, E.; Bongiovanni, D.; Musso, G. Gut microbiota, hypertension and chronic kidney disease: Recent advances. Pharmacol. Res. 2019, 144, 390–408. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Jia, Q.; Yang, J.; Jia, R.; Zhang, H. Efficacy of Probiotics Supplementation on Chronic Kidney Disease: A Systematic Review and Meta-Analysis. Kidney Blood Press. Res. 2018, 43, 1623–1635. [Google Scholar] [CrossRef] [PubMed]
- Jacouton, E.; Michel, M.-L.; Torres-Maravilla, E.; Chain, F.; Langella, P.; Bermúdez-Humarán, L.G. Elucidating the Immune-Related Mechanisms by Which Probiotic Strain Lactobacillus casei BL23 Displays Anti-tumoral Properties. Front. Microbiol. 2019, 9, 3281. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xiong, Q.; Zhao, J.; Lin, X.; He, S.; Wu, N.; Yao, Y.; Liang, W.; Zuo, X.; Ying, C. Inulin-type fructan intervention restricts the increase in gut microbiome–generated indole in patients with peritoneal dialysis: A randomized crossover study. Am. J. Clin. Nutr. 2020, 1, nqz337. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Duranti, D.; Chiara, R.; Basile, M.; Bagnati, M.J.A.R.D.M. Short-Term Effects of Pre/Probiotics on P-Cresol and Indoxyl-Sulphate Serum Concentrations During the Various Stages of Chronic Kidney Disease. Arch. Renal. Dis. Manag. 2017, 3, 001–005. [Google Scholar]
- Lee, H.M.; Lee, Y. A differential medium for lactic acid-producing bacteria in a mixed culture. Lett. Appl. Microbiol. 2008, 46, 676–681. [Google Scholar] [CrossRef]
- Koppe, L.; Mafra, D.; Fouque, D. Probiotics and chronic kidney disease. Kidney Int. 2015, 88, 958–966. [Google Scholar] [CrossRef]
- Sabatino, A.; Regolisti, G.; Brusasco, I.; Cabassi, A.; Morabito, S.; Fiaccadori, E. Alterations of intestinal barrier and microbiota in chronic kidney disease. Nephrol. Dial. Transplant. 2015, 30, 924–933. [Google Scholar] [CrossRef]
- Poesen, R.; Evenepoel, P.; de Loor, H.; Kuypers, D.; Augustijns, P.; Meijers, B. Metabolism, Protein Binding, and Renal Clearance of Microbiota–Derived p-Cresol in Patients with CKD. Clin. J. Am. Soc. Nephrol. 2016, 11, 1136–1144. [Google Scholar] [CrossRef]
- Anders, H.J.; Andersen, K.; Stecher, B. The intestinal microbiota, a leaky gut, and abnormal immunity in kidney disease. Kidney Int. 2013, 83, 1010–1016. [Google Scholar] [CrossRef]
- Fang, C.Y.; Lu, J.R.; Chen, B.J.; Wu, C.; Chen, Y.P.; Chen, M.J. Selection of uremic toxin-reducing probiotics in vitro and in vivo. J. Funct. Foods 2014, 7, 407–415. [Google Scholar] [CrossRef]
- Nowak, A.; Libudzisz, Z. Ability of intestinal lactic bacteria to bind or/and metabolise phenol and p-cresol. Ann. Microbiol. 2007, 57, 329–335. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, Y.S.; Liu, F. The influence of mutant lactobacilli on serum creatinine and urea nitrogen concentrations and renal pathology in 5/6 nephrectomized rats. Ren. Fail. 2016, 38, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Li, K.Y.; Wang, P.J.; Huang, H.W.; Chen, M.J. Alleviating chronic kidney disease progression through modulating the critical genus of gut microbiota in a cisplatin-induced Lanyu pig model. J. Food Drug Anal. 2020, 28, 103–114. [Google Scholar] [CrossRef]
- Lai, S.; Molfino, A.; Testorio, M.; Perrotta, A.M.; Currado, A.; Pintus, G.; Pietrucci, D.; Unida, V.; La Rocca, D.; Biocca, S.; et al. Effect of Low-Protein Diet and Inulin on Microbiota and Clinical Parameters in Patients with Chronic Kidney Disease. Nutrients 2019, 11, 3306. [Google Scholar] [CrossRef]
- Alla, F.; Sadeek, E.A. Effect of Arabic Gum as prebiotics and Lactobacillus casei Shirota (LcS) as probiotic on oxidative stress and renal function in adenine–induced chronic renal failure in rats. Eur. J. Nutr. Food Saf. 2018, 8, 29–46. [Google Scholar]
- Rocchetti, M.T.; Cosola, C.; di Bari, I.; Magnani, S.; Galleggiante, V.; Scandiffio, L.; Dalfino, G.; Netti, G.S.; Atti, M.; Corciulo, R.; et al. Efficacy of Divinylbenzenic Resin in Removing Indoxyl Sulfate and P-Cresol Sulfate in Hemodialysis Patients: Results from an In Vitro Study and an In Vivo Pilot Trial (xuanro4-Nature 3.2). Toxins 2020, 12, 170. [Google Scholar] [CrossRef]
- Wu, B.; Jiang, H.; He, Q.; Wang, M.; Xue, J.; Liu, H.; Shi, K.; Wei, M.; Liang, S.; Zhang, L. Liquid Chromatography/Mass Spectrometry Reveals the Effect of Lactobacillus Treatment on the Faecal Metabolite Profile of Rats with Chronic Renal Failure. Nephron 2017, 135, 156–166. [Google Scholar] [CrossRef]
- Wanchai, K.; Yasom, S.; Tunapong, W.; Chunchai, T.; Eaimworawuthikul, S.; Thiennimitr, P.; Chaiyasut, C.; Pongchaidecha, A.; Chatsudthipong, V.; Chattipakorn, S.; et al. Probiotic Lactobacillus paracasei HII01 protects rats against obese-insulin resistance-induced kidney injury and impaired renal organic anion transporter 3 function. Clin. Sci. 2018, 132, 1545–1563. [Google Scholar] [CrossRef]
- Vanholder, R.; Schepers, E.; Pletinck, A.; Nagler, E.V.; Glorieux, G. The uremic toxicity of indoxyl sulfate and p-cresyl sulfate: A systematic review. J. Am. Soc. Nephrol. 2014, 25, 1897–1907. [Google Scholar] [CrossRef]
- Feng, Y.L.; Cao, G.; Chen, D.Q.; Vaziri, N.D.; Chen, L.; Zhang, J.; Wang, M.; Guo, Y.; Zhao, Y.Y. Microbiome-metabolomics reveals gut microbiota associated with glycine-conjugated metabolites and polyamine metabolism in chronic kidney disease. Cell. Mol. Life Sci. 2019, 76, 4961–4978. [Google Scholar] [CrossRef]
- Yisireyili, M.; Shimizu, H.; Saito, S.; Enomoto, A.; Nishijima, F.; Niwa, T. Indoxyl sulfate promotes cardiac fibrosis with enhanced oxidative stress in hypertensive rats. Life Sci. 2013, 92, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Kamiński, T.W.; Pawlak, K.; Karbowska, M.; Myśliwiec, M.; Pawlak, D. Indoxyl sulfate—The uremic toxin linking hemostatic system disturbances with the prevalence of cardiovascular disease in patients with chronic kidney disease. BMC Nephrol. 2017, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-M.; Guo, J.; Zeng, X.-A.; Sun, D.-W.; Brennan, C.S.; Zhou, Q.-X.; Zhou, J.-S. The probiotic role of Lactobacillus plantarum in reducing risks associated with cardiovascular disease. Int. J. Food Sci. Technol. 2017, 52, 127–136. [Google Scholar] [CrossRef]
- Malik, M.; Suboc, T.M.; Tyagi, S.; Salzman, N.; Wang, J.; Ying, R.; Tanner, M.J.; Kakarla, M.; Baker, J.E.; Widlansky, M.E. Lactobacillus plantarum 299v Supplementation Improves Vascular Endothelial Function and Reduces Inflammatory Biomarkers in Men with Stable Coronary Artery Disease. Circ. Res. 2018, 123, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; Wu, V.; Wu, P.C.; Wu, C.J. Meta-Analysis of the Associations of p-Cresyl Sulfate (PCS) and Indoxyl Sulfate (IS) with Cardiovascular Events and All-Cause Mortality in Patients with Chronic Renal Failure. PLoS ONE 2015, 10, e0132589. [Google Scholar] [CrossRef]
- De Faria Barros, A.; Borges, N.A.; Nakao, L.S.; Dolenga, C.J.; do Carmo, F.L.; de Carvalho Ferreira, D.; Stenvinkel, P.; Bergman, P.; Lindholm, B.; Mafra, D. Effects of probiotic supplementation on inflammatory biomarkers and uremic toxins in non-dialysis chronic kidney patients: A double-blind, randomized, placebo-controlled trial. J. Funct. Foods 2018, 46, 378–383. [Google Scholar] [CrossRef]
- Karbowska, M.; Kaminski, T.W.; Marcinczyk, N.; Misztal, T.; Rusak, T.; Smyk, L.; Pawlak, D. The Uremic Toxin Indoxyl Sulfate Accelerates Thrombotic Response after Vascular Injury in Animal Models. Toxins 2017, 9, 229. [Google Scholar] [CrossRef]
- Wang, X.; Chaudhry, M.A.; Nie, Y.; Xie, Z.; Shapiro, J.I.; Liu, J. A Mouse 5/6th Nephrectomy Model That Induces Experimental Uremic Cardiomyopathy. JoVE 2017, 129, e55825. [Google Scholar]
- Leary, S.L.; Underwood, W.; Anthony, R.; Cartner, S.; Corey, D.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.A.; Meyer, R. AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- Muñoz, D.; Castillo, H.; Henriquez, J.P.; Marcellini, S. Bone regeneration after traumatic skull injury in Xenopus tropicalis. Mech. Dev. 2018, 154, 153–161. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Time 0 h (mM) | Time 48 h (mM) | Decrease (%) |
|---|---|---|---|
| 1c2 | 2.77 ± 0.13 | 2.48 ± 0.05 | 11% |
| 6c3 | 2.93 ± 0.23 | 2.47 ± 0.07 | 16% |
| VIIIc2 | 3.49 ± 0.10 | 3.04 ± 0.02 | 12% |
| 1c2 + 6c3 + VIIIc2 | 2.71 ± 0.01 | 2.53 ± 0.09 | 7% |
| Group | Fibrotic Area (%) |
|---|---|
| Control | 0 ± 0 |
| Nef | 25 ± 1.9 |
| Lac | 12 ± 1.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerez-Morales, A.; Merino, J.S.; Díaz-Castillo, S.T.; Smith, C.T.; Fuentealba, J.; Bernasconi, H.; Echeverría, G.; García-Cancino, A. The Administration of the Synbiotic Lactobacillus bulgaricus 6c3 Strain, Inulin and Fructooligosaccharide Decreases the Concentrations of Indoxyl Sulfate and Kidney Damage in a Rat Model. Toxins 2021, 13, 192. https://doi.org/10.3390/toxins13030192
Jerez-Morales A, Merino JS, Díaz-Castillo ST, Smith CT, Fuentealba J, Bernasconi H, Echeverría G, García-Cancino A. The Administration of the Synbiotic Lactobacillus bulgaricus 6c3 Strain, Inulin and Fructooligosaccharide Decreases the Concentrations of Indoxyl Sulfate and Kidney Damage in a Rat Model. Toxins. 2021; 13(3):192. https://doi.org/10.3390/toxins13030192
Chicago/Turabian StyleJerez-Morales, Alonso, José S. Merino, Sindy T. Díaz-Castillo, Carlos T. Smith, Jorge Fuentealba, Humberto Bernasconi, Gerson Echeverría, and Apolinaria García-Cancino. 2021. "The Administration of the Synbiotic Lactobacillus bulgaricus 6c3 Strain, Inulin and Fructooligosaccharide Decreases the Concentrations of Indoxyl Sulfate and Kidney Damage in a Rat Model" Toxins 13, no. 3: 192. https://doi.org/10.3390/toxins13030192
APA StyleJerez-Morales, A., Merino, J. S., Díaz-Castillo, S. T., Smith, C. T., Fuentealba, J., Bernasconi, H., Echeverría, G., & García-Cancino, A. (2021). The Administration of the Synbiotic Lactobacillus bulgaricus 6c3 Strain, Inulin and Fructooligosaccharide Decreases the Concentrations of Indoxyl Sulfate and Kidney Damage in a Rat Model. Toxins, 13(3), 192. https://doi.org/10.3390/toxins13030192