The Two Sweet Sides of Janus Lectin Drive Crosslinking of Liposomes to Cancer Cells and Material Uptake

 , , and
, , and 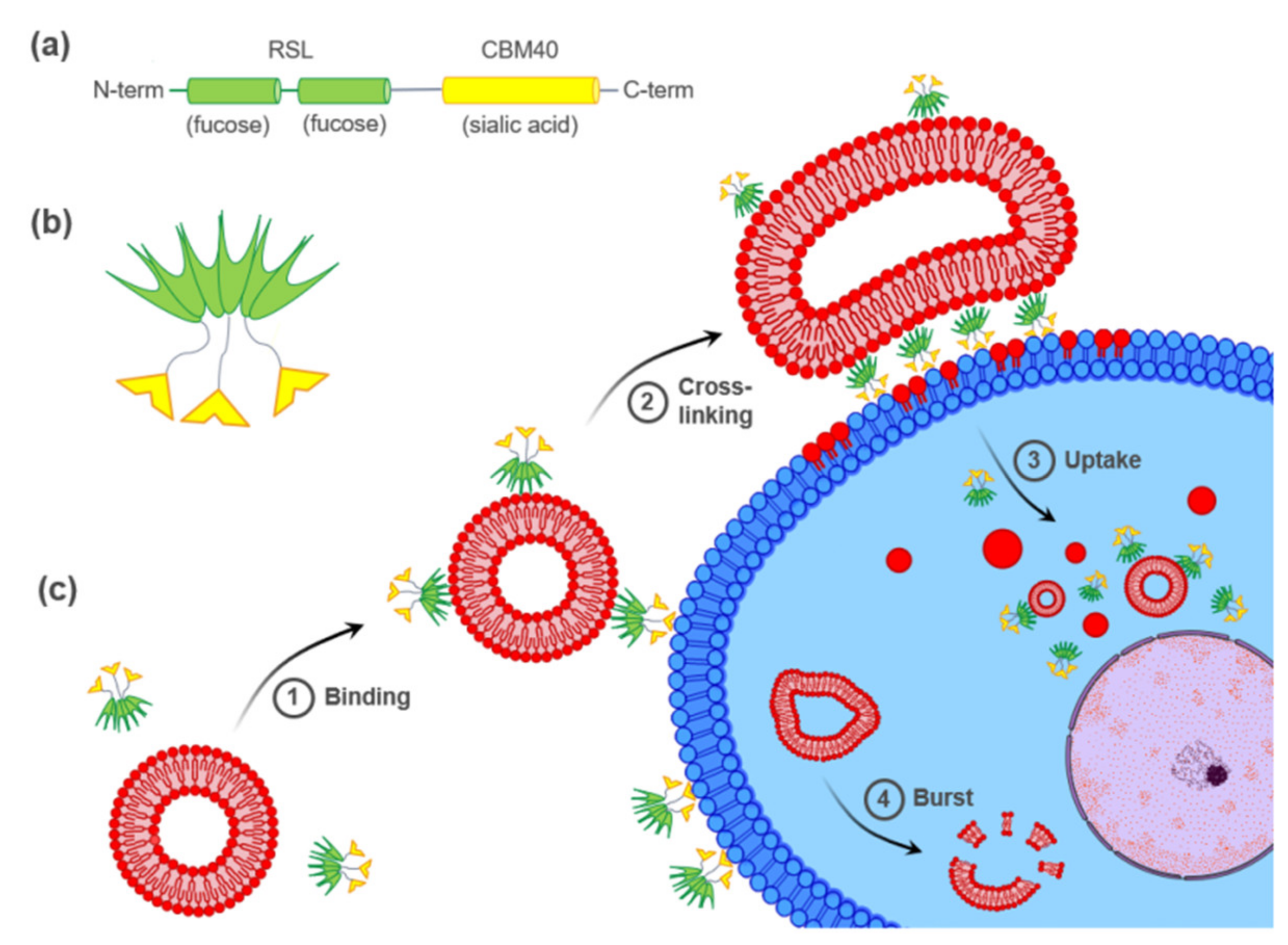{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
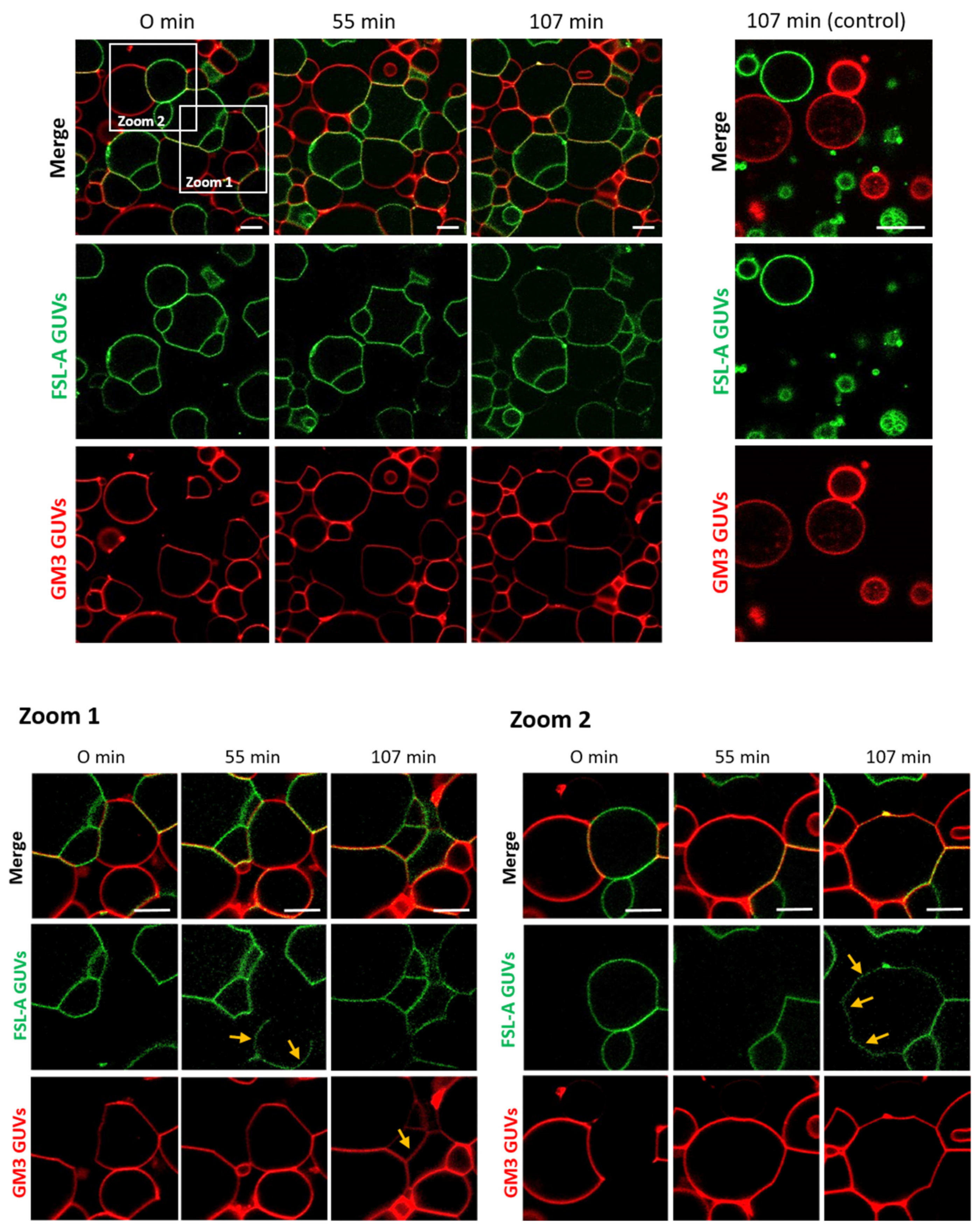
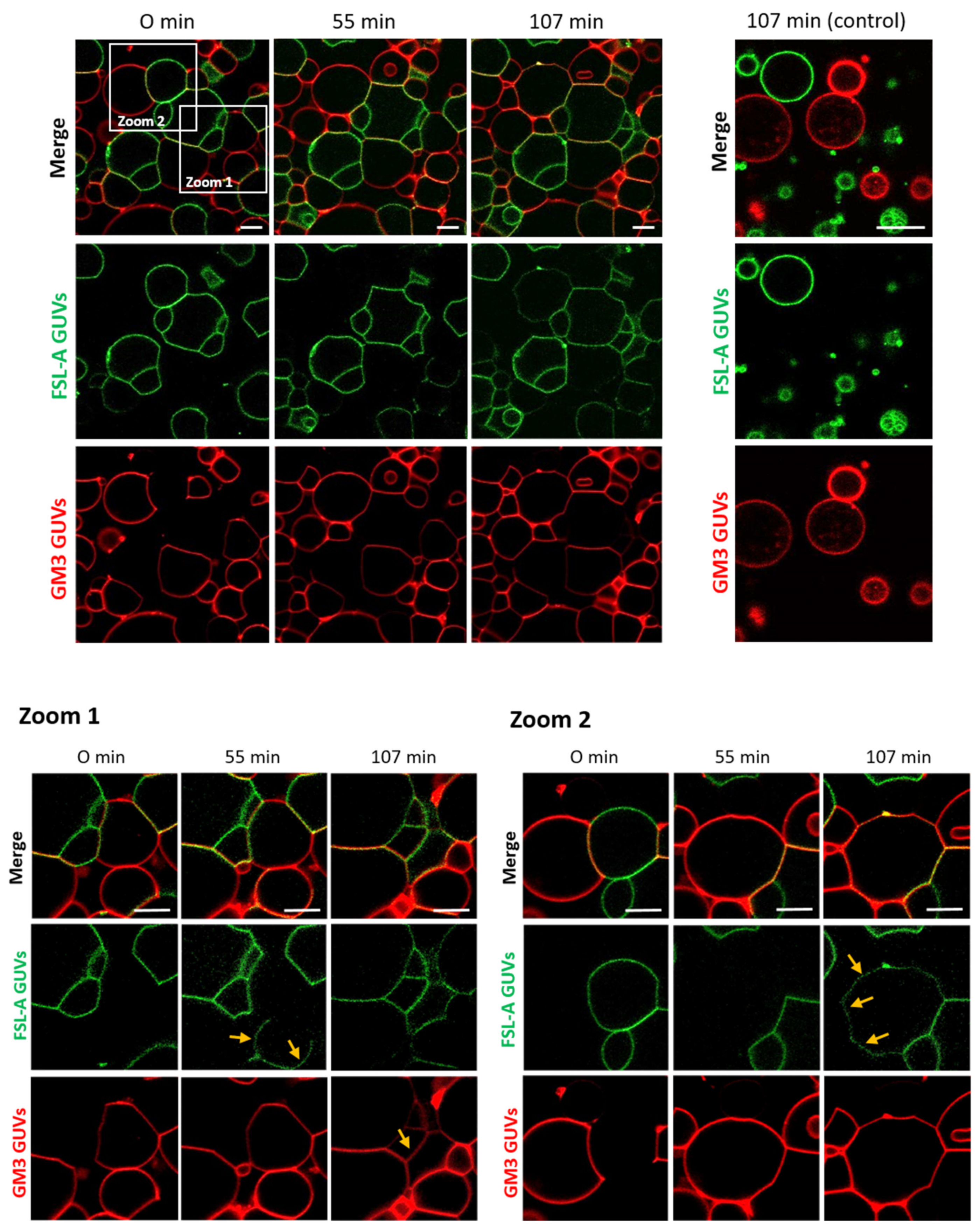
2.1. FS-Janus Lectin Induces Lipid Exchange between GM3- and Blood Group A Trisaccharide-Decorated GUVs
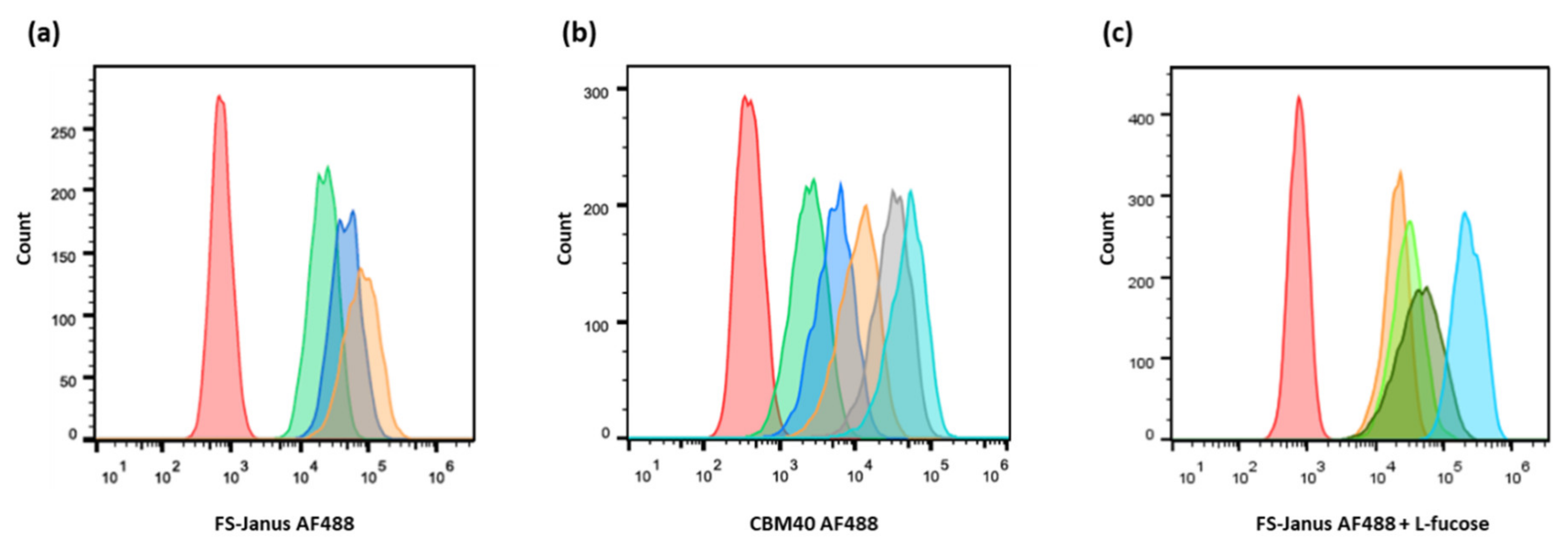
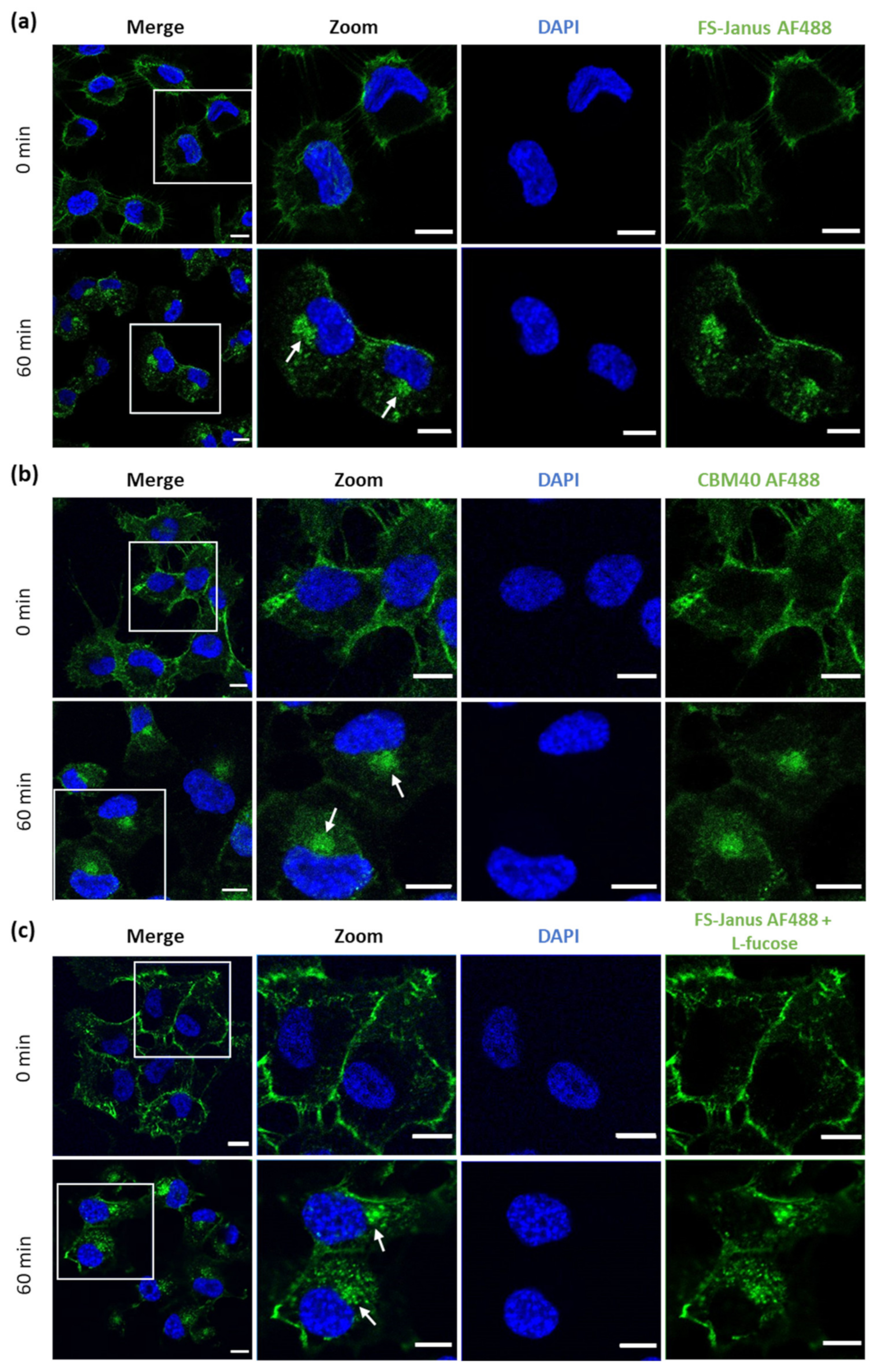
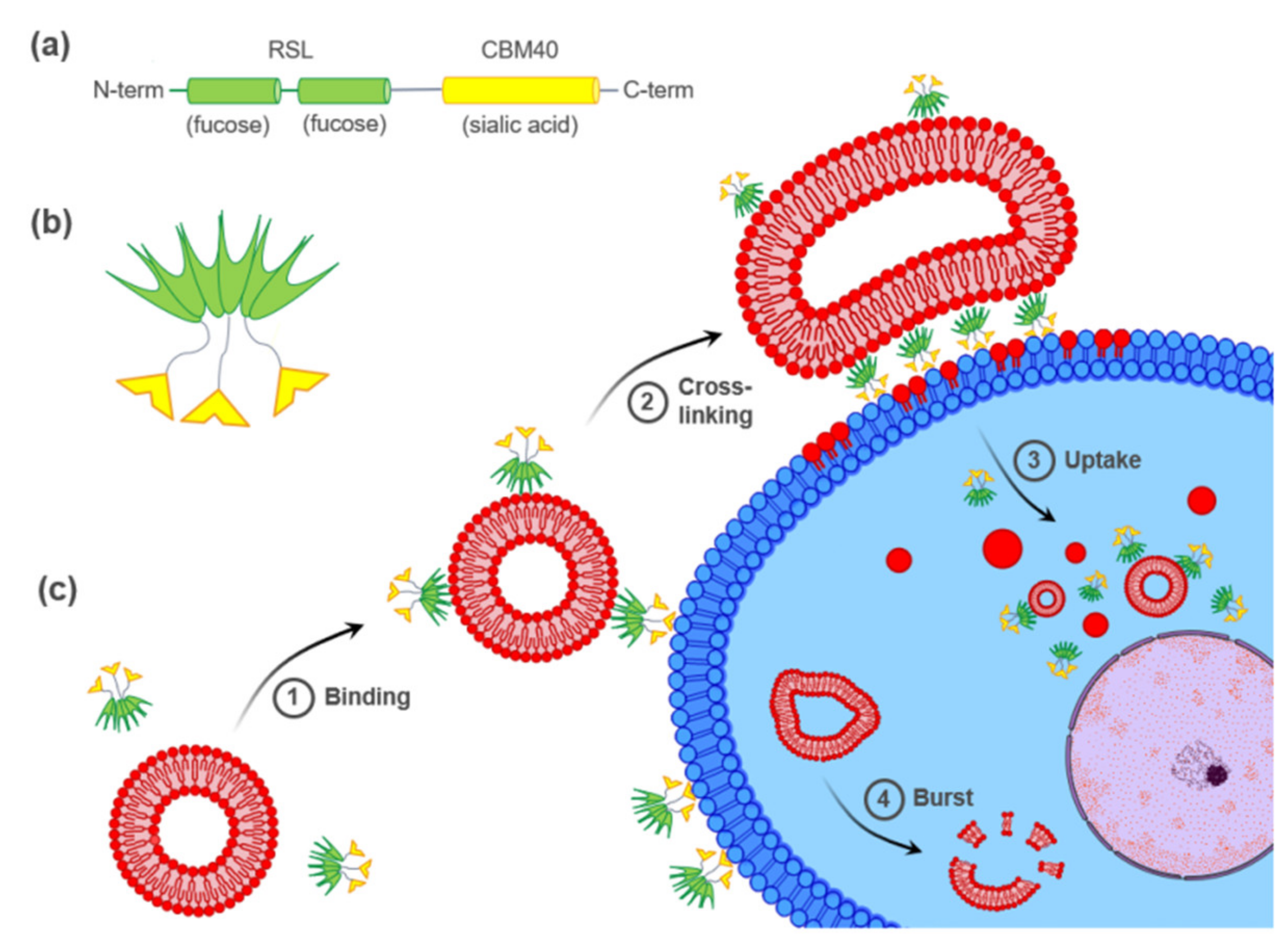
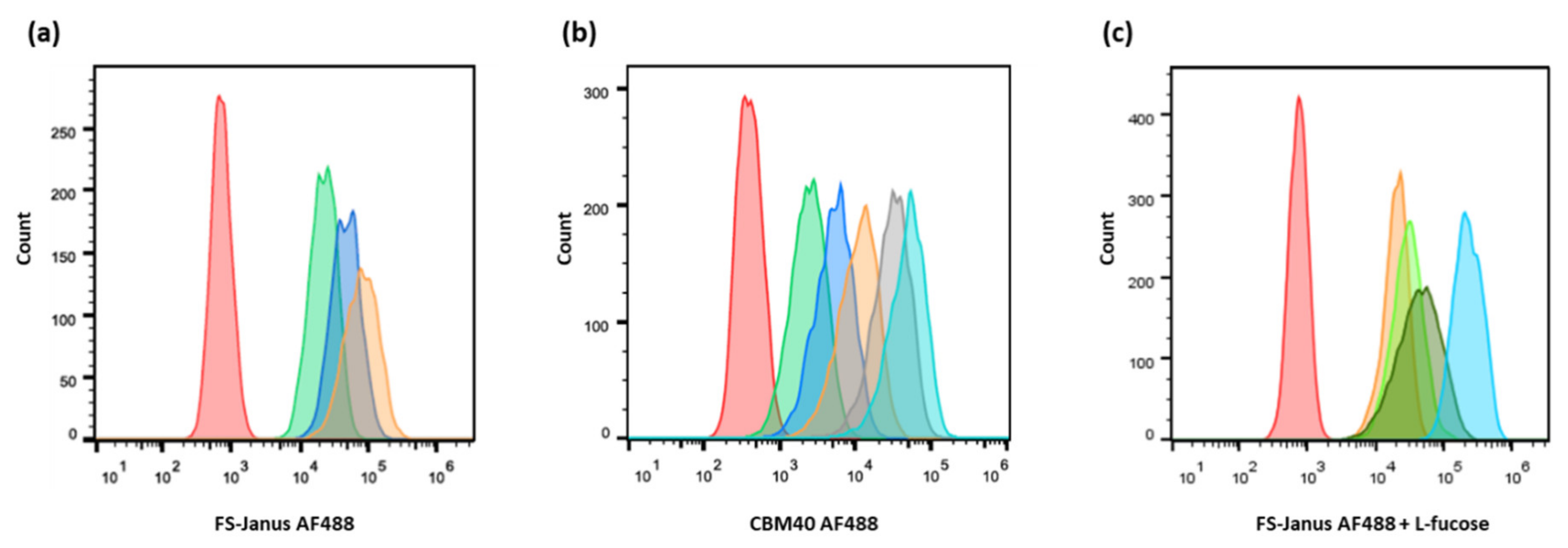
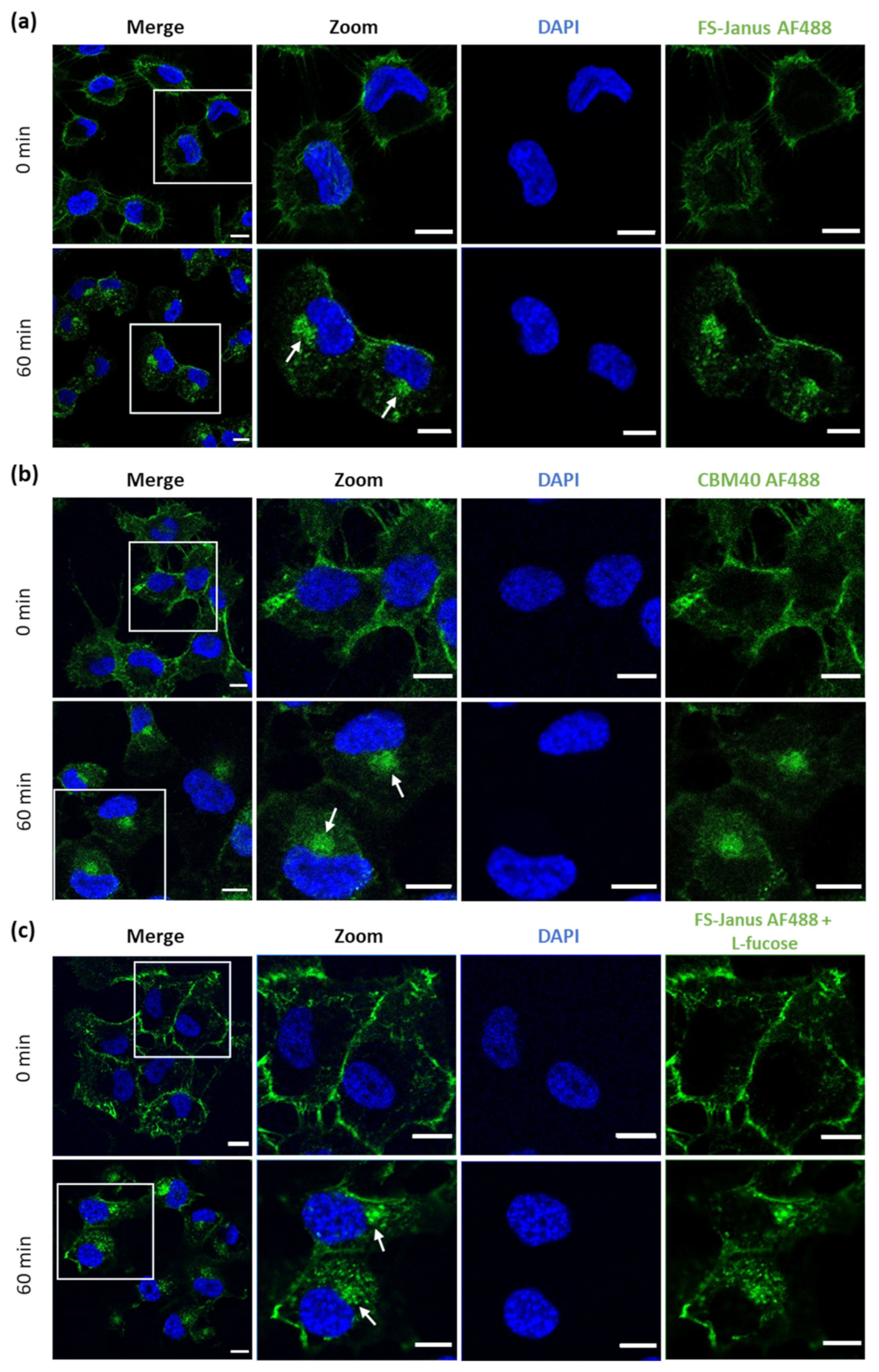
2.2. FS-Janus Lectin and di-CBM40 Bind and Internalize into Human Epithelial Cancer Cells
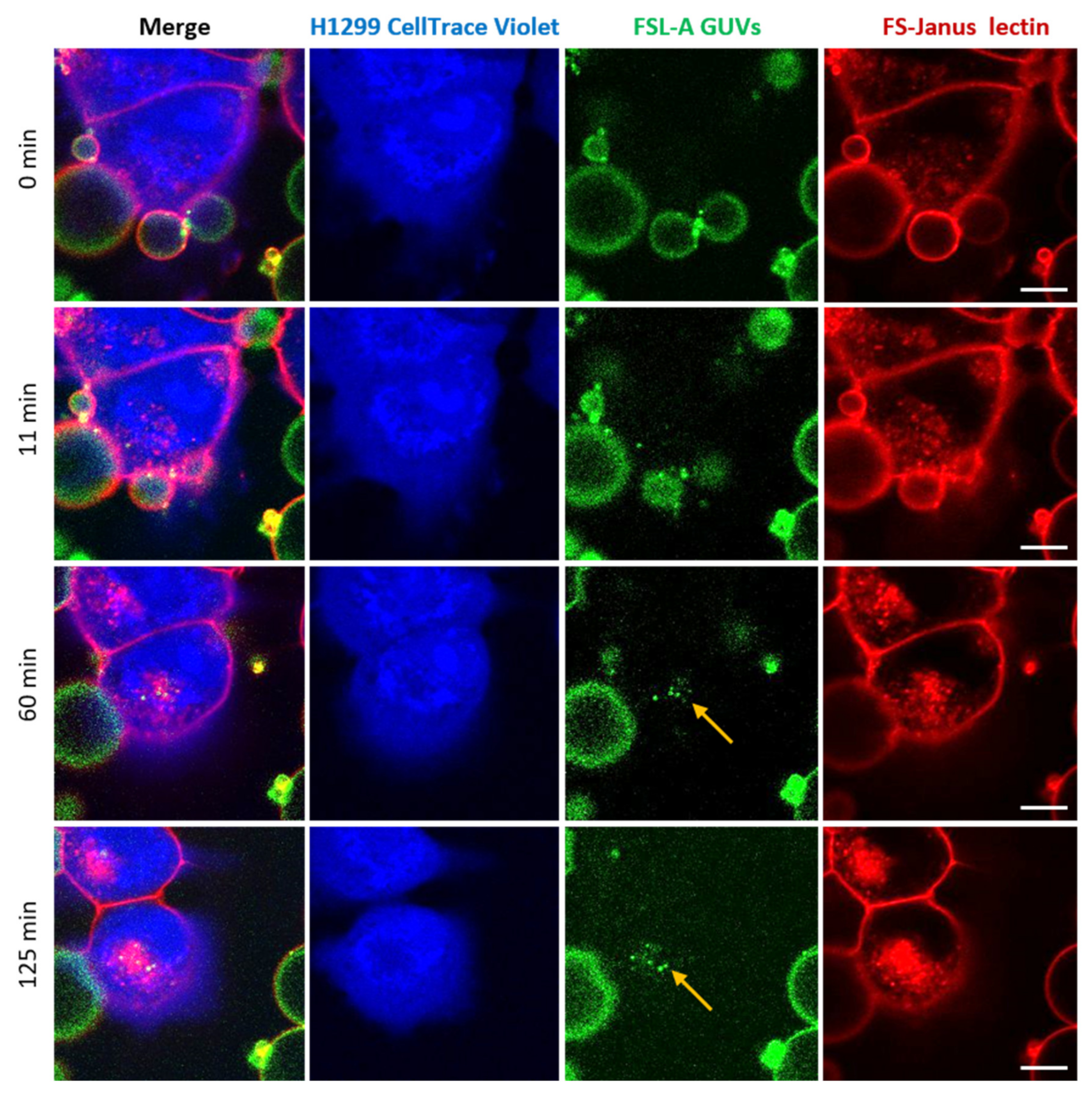
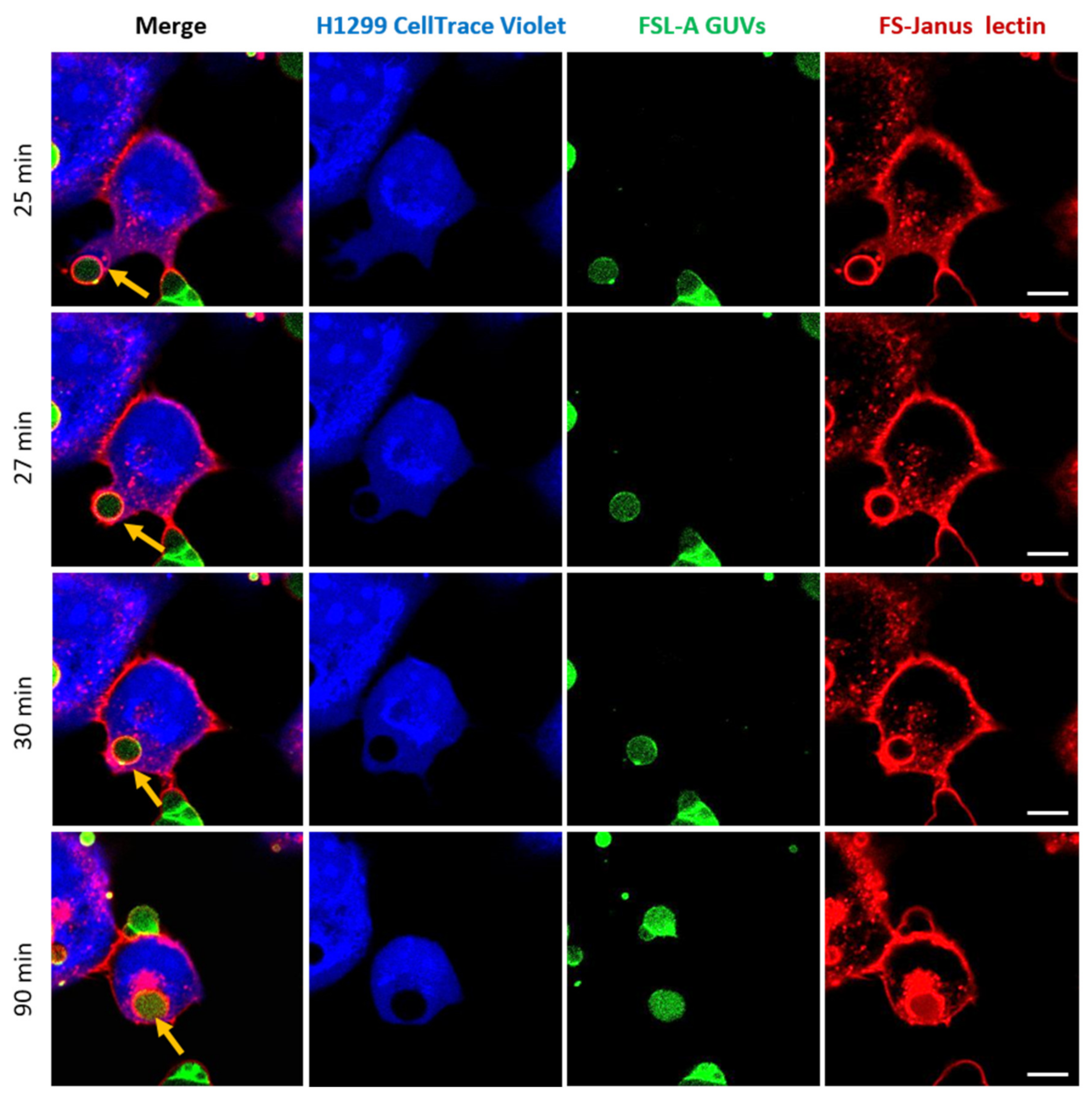
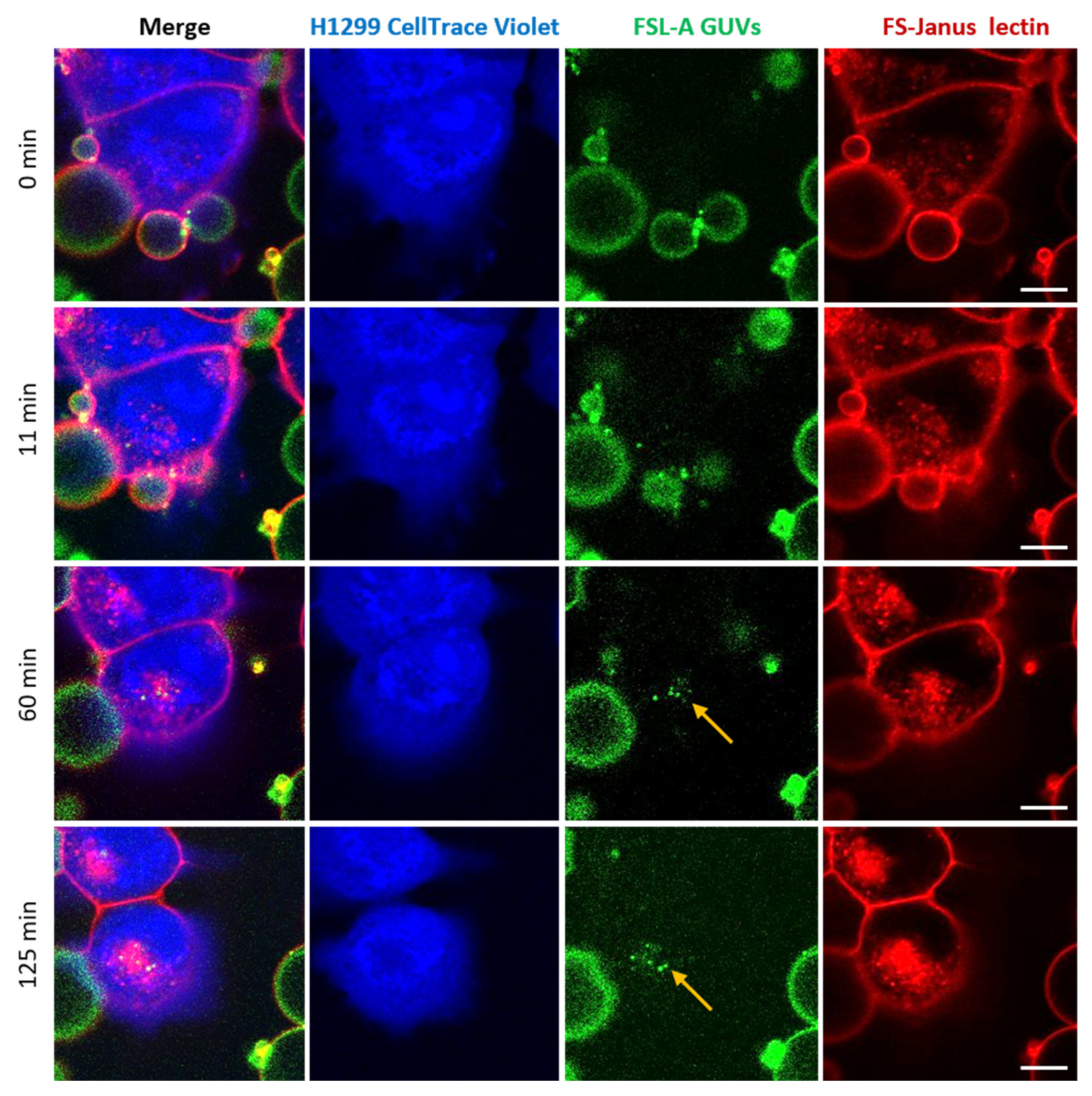
2.3. FS-Janus Lectin Mediates Crosslinking of Liposomes with H1299 Cells and the Cellular Uptake of Liposomal Content
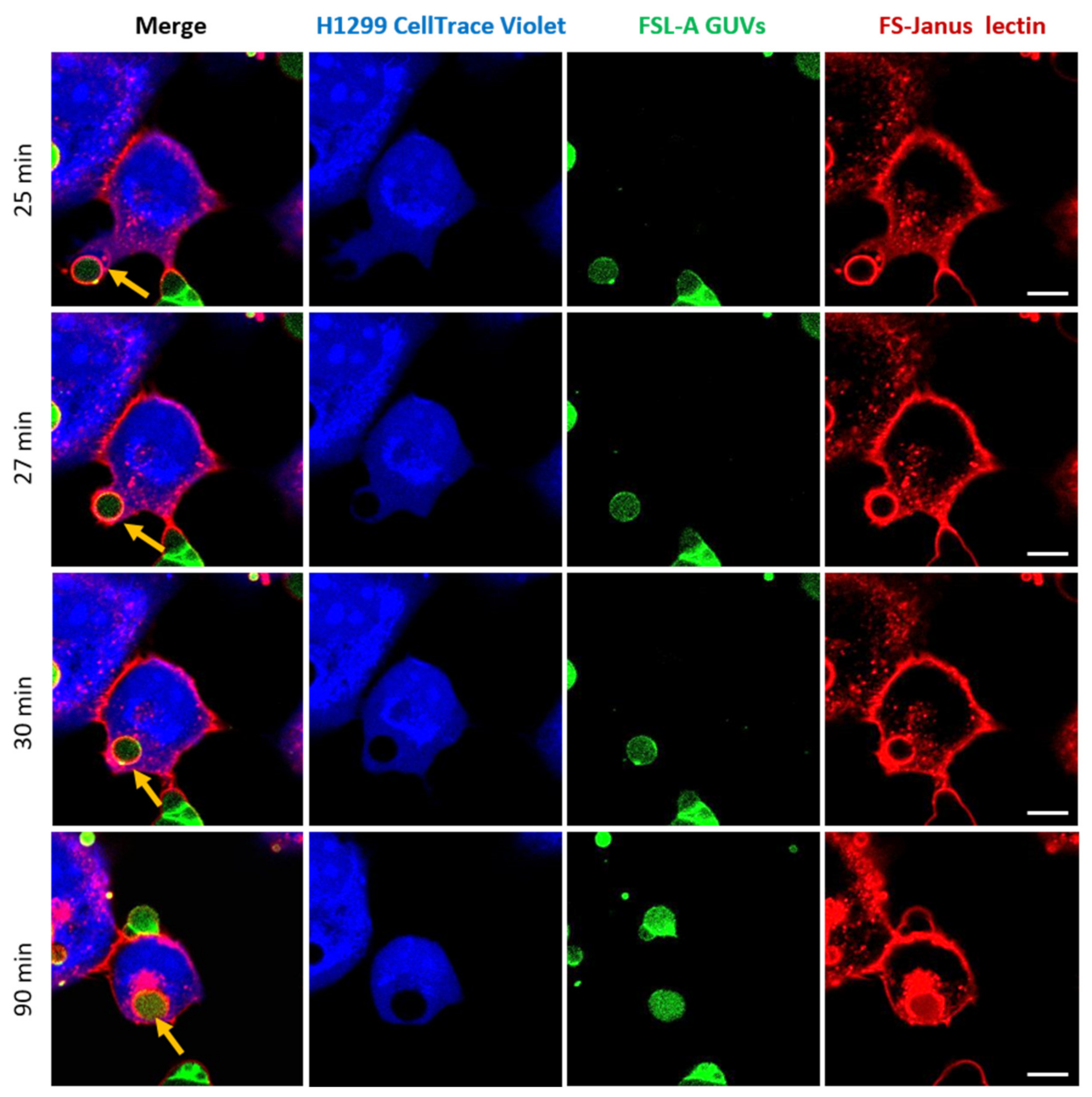
2.4. FS-Janus Lectin Induces the Uptake of Complete Liposomes into H1299 Cells
2.5. Uptaken Liposomes Are Deformed and Burst
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Production of Lectins and Labeling
5.2. Composition and Preparation of GUVs
5.3. Cell Culture
5.4. Flow Cytometry Analysis
5.5. Lectin Stimulation and Fluorescence Microscopy
5.6. Live-Cell Imaging
5.7. Chemical Reagents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mody, R.; Joshi, S.H.; Chaney, W. Use of Lectins as Diagnostic and Therapeutic Tools for Cancer. J. Pharmacol. Toxicol. Methods 1995, 33, 1–10. [Google Scholar] [CrossRef]
- Gorelik, E.; Galili, U.; Raz, A. On the Role of Cell Surface Carbohydrates and Their Binding Proteins (Lectins) in Tumor Metastasis. Cancer Metastasis Rev. 2001, 20, 245–277. [Google Scholar] [CrossRef] [PubMed]
- Bies, C.; Lehr, C.-M.; Woodley, J.F. Lectin-Mediated Drug Targeting: History and Applications. Adv. Drug Deliv. Rev. 2004, 56, 425–435. [Google Scholar] [CrossRef]
- Minko, T. Drug Targeting to the Colon with Lectins and Neoglycoconjugates. Adv. Drug Deliv. Rev. 2004, 56, 491–509. [Google Scholar] [CrossRef] [PubMed]
- Rini, J.M. Lectin Structure. Annu. Rev. Biophys. Biomol. Struct. 1995, 24, 551–577. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Lectins: Carbohydrate-Specific Proteins That Mediate Cellular Recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef]
- Sharon, N. Lectins: Carbohydrate-Specific Reagents and Biological Recognition Molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef] [Green Version]
- Ambrosi, M.; Cameron, N.R.; Davis, B.G. Lectins: Tools for the Molecular Understanding of the Glycocode. Org. Biomol. Chem. 2005, 3, 1593–1608. [Google Scholar] [CrossRef]
- Weis, W.I.; Drickamer, K. Structural Basis of Lectin-Carbohydrate Recognition. Annu. Rev. Biochem. 1996, 65, 441–473. [Google Scholar] [CrossRef]
- Collins, B.E.; Paulson, J.C. Cell Surface Biology Mediated by Low Affinity Multivalent Protein–Glycan Interactions. Curr. Opin. Chem. Biol. 2004, 8, 617–625. [Google Scholar] [CrossRef]
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A Glycobiology Review: Carbohydrates, Lectins and Implications in Cancer Therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Ohtsubo, K.; Marth, J.D. Glycosylation in Cellular Mechanisms of Health and Disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Imberty, A.; Varrot, A. Microbial Recognition of Human Cell Surface Glycoconjugates. Curr. Opin. Struct. Biol. 2008, 18, 567–576. [Google Scholar] [CrossRef]
- Brandley, B.K.; Schnaar, R.L. Cell-Surface Carbohydrates in Cell Recognition and Response. J. Leukoc. Biol. 1986, 40, 97–111. [Google Scholar] [CrossRef]
- Eierhoff, T.; Bastian, B.; Thuenauer, R.; Madl, J.; Audfray, A.; Aigal, S.; Juillot, S.; Rydell, G.E.; Muller, S.; de Bentzmann, S.; et al. A Lipid Zipper Triggers Bacterial Invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 12895–12900. [Google Scholar] [CrossRef] [Green Version]
- Youan, B.-B.C.; Coulibaly, F.S. Current Status of Lectin-Based Cancer Diagnosis and Therapy. AIMS Mol. Sci. 2017, 4, 1–27. [Google Scholar] [CrossRef]
- Müller, S.K.; Wilhelm, I.; Schubert, T.; Zittlau, K.; Imberty, A.; Madl, J.; Eierhoff, T.; Thuenauer, R.; Römer, W. Gb3-Binding Lectins as Potential Carriers for Transcellular Drug Delivery. Expert Opin. Drug Deliv. 2017, 14, 141–153. [Google Scholar] [CrossRef]
- Varki, A.; Kannagi, R.; Toole, B.P. Glycosylation Changes in Cancer. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Pinho, S.S.; Reis, C.A. Glycosylation in Cancer: Mechanisms and Clinical Implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Rodrigues, J.G.; Balmaña, M.; Macedo, J.A.; Poças, J.; Fernandes, Â.; de-Freitas-Junior, J.C.M.; Pinho, S.S.; Gomes, J.; Magalhães, A.; Gomes, C.; et al. Glycosylation in Cancer: Selected Roles in Tumour Progression, Immune Modulation and Metastasis. Cell. Immunol. 2018, 333, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odumosu, O.; Nicholas, D.; Yano, H.; Langridge, W. AB Toxins: A Paradigm Switch from Deadly to Desirable. Toxins 2010, 2, 1612–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johannes, L.; Römer, W. Shiga Toxins—from Cell Biology to Biomedical Applications. Nat. Rev. Microbiol. 2010, 8, 105–116. [Google Scholar] [CrossRef]
- Guan, J.; Zhang, Z.; Hu, X.; Yang, Y.; Chai, Z.; Liu, X.; Liu, J.; Gao, B.; Lu, W.; Qian, J.; et al. Cholera Toxin Subunit B Enabled Multifunctional Glioma-Targeted Drug Delivery. Adv. Healthc. Mater. 2017, 6, 1700709. [Google Scholar] [CrossRef] [PubMed]
- Fort, P.; Sakai, K.; Luppi, P.-H.; Salvert, D.; Jouvet, M. Monoaminergic, Peptidergic, and Cholinergic Afferents to the Cat Facial Nucleus as Evidenced by a Double Immunostaining Method with Unconjugated Cholera Toxin as a Retrograde Tracer. J. Comp. Neurol. 1989, 283, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.; Coimbra, A. Morphological Types of Spinomesencephalic Neurons in the Marginal Zone (Lamina I) of the Rat Spinal Cord, as Shown after Retrograde Labelling with Cholera Toxin Subunit B. J. Comp. Neurol. 1989, 279, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.-H.; Fort, P.; Jouvet, M. Iontophoretic Application of Unconjugated Cholera Toxin B Subunit (CTb) Combined with Immunohistochemistry of Neurochemical Substances: A Method for Transmitter Identification of Retrogradely Labeled Neurons. Brain Res. 1990, 534, 209–224. [Google Scholar] [CrossRef]
- Luppi, P.-H.; Sakai, K.; Salvert, D.; Fort, P.; Jouvet, M. Peptidergic Hypothalamic Afferents to the Cat Nucleus Raphe Pallidus as Revealed by a Double Immunostaining Technique Using Unconjugated Cholera Toxin as a Retrograde Tracer. Brain Res. 1987, 402, 339–345. [Google Scholar] [CrossRef]
- Mo, Y.; Lim, L.-Y. Mechanistic Study of the Uptake of Wheat Germ Agglutinin-Conjugated PLGA Nanoparticles by A549 Cells. J. Pharm. Sci. 2004, 93, 20–28. [Google Scholar] [CrossRef]
- Mo, Y.; Lim, L. Preparation and in Vitro Anticancer Activity of Wheat Germ Agglutinin (WGA)-Conjugated PLGA Nanoparticles Loaded with Paclitaxel and Isopropyl Myristate. J. Control. Release 2005, 107, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Winter, H.C.; Mostafapour, K.; Goldstein, I.J. The Mushroom Marasmius Oreades Lectin Is a Blood Group Type B Agglutinin That Recognizes the Galα1,3Gal and Galα1,3Galβ1,4GlcNAc Porcine Xenotransplantation Epitopes with High Affinity. J. Biol. Chem. 2002, 277, 14996–15001. [Google Scholar] [CrossRef] [Green Version]
- Terada, D.; Kawai, F.; Noguchi, H.; Unzai, S.; Hasan, I.; Fujii, Y.; Park, S.-Y.; Ozeki, Y.; Tame, J.R.H. Crystal Structure of MytiLec, a Galactose-Binding Lectin from the Mussel Mytilus Galloprovincialis with Cytotoxicity against Certain Cancer Cell Types. Sci. Rep. 2016, 6, 28344. [Google Scholar] [CrossRef]
- Dobie, C.; Skropeta, D. Insights into the Role of Sialylation in Cancer Progression and Metastasis. Br. J. Cancer 2021, 124, 76–90. [Google Scholar] [CrossRef]
- Büll, C.; Stoel, M.A.; den Brok, M.H.; Adema, G.J. Sialic Acids Sweeten a Tumor’s Life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Blixt, O.; Wandall, H.H. Mucins as biomarkers in cancer. In Mucins and Cancer; Future Medicine Ltd.: London, UK, 2013. [Google Scholar]
- Drake, P.M.; Cho, W.; Li, B.; Prakobphol, A.; Johansen, E.; Anderson, N.L.; Regnier, F.E.; Gibson, B.W.; Fisher, S.J. Sweetening the Pot: Adding Glycosylation to the Biomarker Discovery Equation. Clin. Chem. 2010, 56, 223–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajaria, B.N.; Patel, K.R.; Begum, R.; Patel, P.S. Sialylation: An Avenue to Target Cancer Cells. Pathol. Oncol. Res. 2016, 22, 443–447. [Google Scholar] [CrossRef]
- Ribeiro, J.P.; Pau, W.; Pifferi, C.; Renaudet, O.; Varrot, A.; Mahal, L.K.; Imberty, A. Characterization of a High-Affinity Sialic Acid-Specific CBM40 from Clostridium Perfringens and Engineering of a Divalent Form. Biochem. J. 2016, 473, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.P.; Villringer, S.; Goyard, D.; Coche-Guerente, L.; Höferlin, M.; Renaudet, O.; Römer, W.; Imberty, A. Tailor-Made Janus Lectin with Dual Avidity Assembles Glycoconjugate Multilayers and Crosslinks Protocells. Chem. Sci. 2018, 9, 7634–7641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostlánová, N.; Mitchell, E.P.; Lortat-Jacob, H.; Oscarson, S.; Lahmann, M.; Gilboa-Garber, N.; Chambat, G.; Wimmerová, M.; Imberty, A. The Fucose-Binding Lectin from Ralstonia Solanacearum. J. Biol. Chem. 2005, 280, 27839–27849. [Google Scholar] [CrossRef] [Green Version]
- Omidvar, R.; Römer, W. Glycan-Decorated Protocells: Novel Features for Rebuilding Cellular Processes. Interface Focus 2019, 9, 20180084. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Chen, X.; Han, Y.; Lei, T.; Wu, Q.; Yu, X.; Wang, L.; Fan, Z.; Wang, S. Modification of A2,6-Sialylation Mediates the Invasiveness and Tumorigenicity of Non-Small Cell Lung Cancer Cells In Vitro and In Vivo via Notch1/Hes1/MMPs Pathway. Int. J. Cancer 2018, 143, 2319–2330. [Google Scholar] [CrossRef] [Green Version]
- Büll, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; Bloemendal, V.R.L.J.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T Cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkley, J.; Scott, E. Targeting Aberrant Sialylation to Treat Cancer. Medicines 2019, 6, 102. [Google Scholar] [CrossRef] [Green Version]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef]
- Đevenica, D.; Čikeš Čulić, V.; Vuica, A.; Markotić, A. Biochemical, Pathological and Oncological Relevance of Gb3Cer Receptor. Med Oncol. 2011, 28, 675–684. [Google Scholar] [CrossRef]
- Miller, K. The Stimulation of Human B and T Lymphocytes by Various Lectins. Immunobiology 1983, 165, 132–146. [Google Scholar] [CrossRef]
- Morgan, D.; Ruscetti, F.; Gallo, R. Selective in Vitro Growth of T Lymphocytes from Normal Human Bone Marrows. Science 1976, 193, 1007–1008. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [Green Version]
- Hua, S.; Wu, S.Y. The Use of Lipid-Based Nanocarriers for Targeted Pain Therapies. Front. Pharmacol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Takeda, T.; Michiue, H.; Abe, T.; Takei, K. Actin Bundling by Dynamin 2 and Cortactin Is Implicated in Cell Migration by Stabilizing Filopodia in Human Non-Small Cell Lung Carcinoma Cells. Int. J. Oncol. 2016, 49, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Gallop, J.L. Filopodia and Their Links with Membrane Traffic and Cell Adhesion. Semin. Cell Dev. Biol. 2020, 102, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Vonna, L.; Wiedemann, A.; Aepfelbacher, M.; Sackmann, E. Micromechanics of Filopodia Mediated Capture of Pathogens by Macrophages. Eur. Biophys. J. 2007, 36, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Underhill, D.M. Mechanisms of Phagocytosis in Macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- Mukai, A.; Kurisaki, T.; Sato, S.B.; Kobayashi, T.; Kondoh, G.; Hashimoto, N. Dynamic Clustering and Dispersion of Lipid Rafts Contribute to Fusion Competence of Myogenic Cells. Exp. Cell Res. 2009, 315, 3052–3063. [Google Scholar] [CrossRef] [PubMed]
- Brück, A.; Abu-dahab, R.; Borchard, G.; Schäfer, U.F.; Lehr, C.-M. Lectin-Functionalized Liposomes for Pulmonary Drug Delivery: Interaction with Human Alveolar Epithelial Cells. J. Drug Target. 2001, 9, 241–251. [Google Scholar] [CrossRef]
- Weber, P.; Dzuricky, M.; Min, J.; Jenkins, I.; Chilkoti, A. Concentration-Independent Multivalent Targeting of Cancer Cells by Genetically Encoded Core-Crosslinked Elastin/Resilin-like Polypeptide Micelles. Biomacromolecules 2021, 22, 4347–4356. [Google Scholar] [CrossRef] [PubMed]
- Fretz, M.M.; Koning, G.A.; Mastrobattista, E.; Jiskoot, W.; Storm, G. OVCAR-3 Cells Internalize TAT-Peptide Modified Liposomes by Endocytosis. Biochim. Biophys. Acta Biomembr. 2004, 1665, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Kusters, R.; Caorsi, V.; Allard, A.; Abou-Ghali, M.; Manzi, J.; di Cicco, A.; Lévy, D.; Lenz, M.; Joanny, J.-F.; et al. Actin Dynamics Drive Cell-like Membrane Deformation. Nat. Phys. 2019, 15, 602–609. [Google Scholar] [CrossRef]
- Madl, J.; Villringer, S.; Römer, W. Delving into Lipid-Driven Endocytic Mechanisms Using Biomimetic Membranes. In Chemical and Synthetic Approaches in Membrane Biology; Humana Press: New York, NY, USA, 2016. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siukstaite, L.; Rosato, F.; Mitrovic, A.; Müller, P.F.; Kraus, K.; Notova, S.; Imberty, A.; Römer, W. The Two Sweet Sides of Janus Lectin Drive Crosslinking of Liposomes to Cancer Cells and Material Uptake. Toxins 2021, 13, 792. https://doi.org/10.3390/toxins13110792
Siukstaite L, Rosato F, Mitrovic A, Müller PF, Kraus K, Notova S, Imberty A, Römer W. The Two Sweet Sides of Janus Lectin Drive Crosslinking of Liposomes to Cancer Cells and Material Uptake. Toxins. 2021; 13(11):792. https://doi.org/10.3390/toxins13110792
Chicago/Turabian StyleSiukstaite, Lina, Francesca Rosato, Anna Mitrovic, Peter Fritz Müller, Katharina Kraus, Simona Notova, Anne Imberty, and Winfried Römer. 2021. "The Two Sweet Sides of Janus Lectin Drive Crosslinking of Liposomes to Cancer Cells and Material Uptake" Toxins 13, no. 11: 792. https://doi.org/10.3390/toxins13110792
APA StyleSiukstaite, L., Rosato, F., Mitrovic, A., Müller, P. F., Kraus, K., Notova, S., Imberty, A., & Römer, W. (2021). The Two Sweet Sides of Janus Lectin Drive Crosslinking of Liposomes to Cancer Cells and Material Uptake. Toxins, 13(11), 792. https://doi.org/10.3390/toxins13110792