Analysis of Cry1Ah Toxin-Binding Reliability to Midgut Membrane Proteins of the Asian Corn Borer



Abstract
1. Introduction
2. Results
2.1. APN and ALP Activities in ACB Larval Midgut
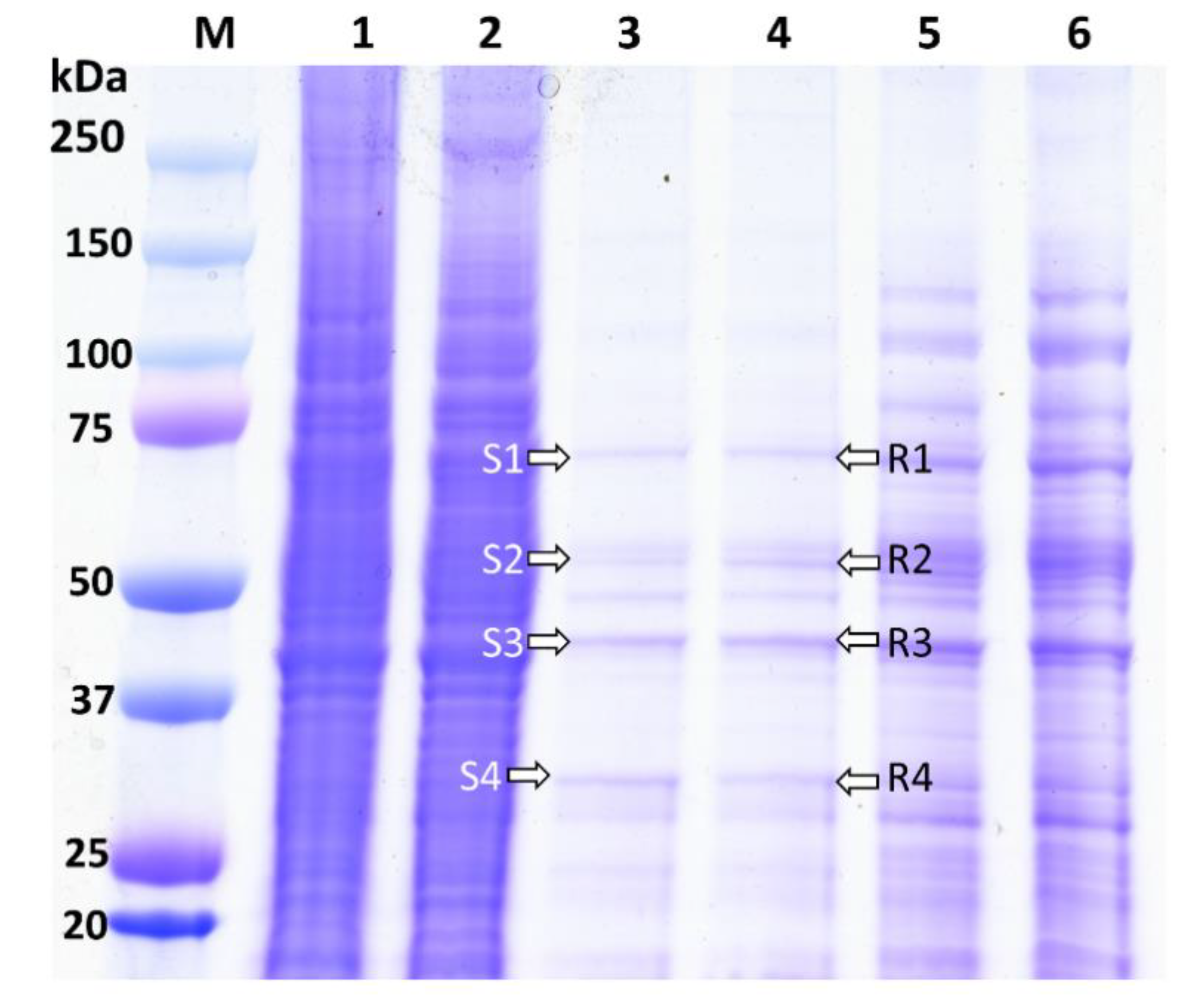
2.2. Cry1Ah-Binding Proteins from BBMV of ACB
2.3. Cry1Ah-Binding Proteins Identified from the BBMVs of the ACB
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Strains
5.2. Preparation of BBMV
5.3. Pull-Down Assay
5.4. Identification of Cry1Ah-Binding Proteins
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crickmore, N.; Baum, J.; Bravo, A.; Lereclus, D.; Narva, K.; Sampson, K.; Schnepf, E.; Sun, M.; Zeigler, D.R. Bacillus Thuringiensis Toxin Nomenclature. 2019. Available online: http://www.btnomenclature.info/ (accessed on 10 February 2020).
- Sanchis, V.; Bourguet, D. Bacillus thuringiensis: Applications in agriculture and insect resistance management. A review. Agron. Sustain. Dev. 2008, 28, 11–20. [Google Scholar] [CrossRef]
- Bagla, P. Hardy cotton-munching pests are latest blow to GM crops. Science 2010, 327, 1439. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Clifton, E.H.; Dunbar, M.W.; Hoffmann, A.M.; Ingber, D.A.; Keweshan, R.S. Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. Proc. Natl. Acad. Sci. USA 2014, 111, 5141–5146. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carriére, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Moar, W.; Roush, R.; Shelton, A.; Ferré, J.; MacIntosh, S.; Leonard, B.R.; Abel, C. Field-evolved resistance to Bt toxins. Nat. Biotechnol. 2008, 26, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Jakka, S.R.K.; Gong, L.; Hasler, J.; Banerjee, R.; Sheets, J.J.; Narva, K. Field evolved mode 1 resistance of the fall armyworm to transgenic Cry1Fa-expressing corn associated with reduced Cry1Fa toxin binding and midgut alkaline phosphatase expression. Appl. Environ. Microbiol. 2015, 82, 1023–1034. [Google Scholar] [CrossRef]
- Ferré, J.; Van Rie, J.; Macintosh, S.C. Insecticidal Genetically Modified Crops and Insect Resistance Management (IRM). In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs; Romeis, J., Shelton, A.M., Kennedy, G.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 5, pp. 41–85. [Google Scholar]
- Soberón, M.; Gill, S.S.; Bravo, A. Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells? Cell. Mol. Life Sci. 2009, 66, 1337–1349. [Google Scholar] [CrossRef]
- Daniel, A.; Sangadala, S.; Dean, D.H.; Adang, M.J. Denaturation of either Manduca sexta aminopeptidase N or Bacillus thuringiensis Cry1A toxins exposes binding epitopes hidden under nondenaturing conditions. Appl. Environ. Microbiol. 2002, 68, 2106–2112. [Google Scholar] [CrossRef]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac {delta}-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. Microbiol. 2005, 71, 948–954. [Google Scholar] [CrossRef]
- Bravo, A.; Soberon, M. How to cope with insect resistance to Bt toxins? Trends Biotechnol. 2008, 26, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Tiewsiri, K.; Wang, P. Differential alteration of two aminopeptidases N associated with resistance to Bacillus thuringiensis toxin Cry1Ac in cabbage looper. Proc. Natl. Acad. Sci. USA 2011, 108, 14037–14042. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [PubMed]
- Candas, M.; Francis, B.R.; Griko, N.B.; Midboe, E.G.; Bulla, L.A., Jr. Proteolytic cleavage of the developmentally important cadherin BT-R1 in the midgut epithelium of Manduca sexta. Biochemistry 2002, 41, 13717–13724. [Google Scholar] [CrossRef]
- Griko, N.; Zhang, X.; Ibrahim, M.; Midboe, E.G.; Bulla, L.A., Jr. Susceptibility of Manduca sexta to Cry1Ab toxin of Bacillus thuringiensis correlates directly to developmental expression of the cadherin receptor BT-R1. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 59–63. [Google Scholar] [CrossRef]
- Hua, G.; Park, Y.; Adang, M.J. Cadherin AdCad1 in Alphitobius diaperinus larvae is a receptor of Cry3Bb toxin from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2014, 45, 11–17. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yaoi, K.; Nagino, Y.; Hara, H.; Kitami, M.; Atsumi, S.; Miura, N.; Sato, R. Aminopeptidase N isoforms from the midgut of Bombyx mori and Plutella xylostella their classification and the factors that determine their binding specificity to Bacillus thuringiensis Cry1A toxin. FEBS Lett. 2002, 519, 215–220. [Google Scholar] [CrossRef]
- Rajagopal, R.; Agrawal, N.; Selvapandiyan, A.; Sivakumar, S.; Ahmad, S.; Bhatnagar, R.K. Recombinantly expressed isoenzymic aminopeptidases from Helicoverpa armigera (American cotton bollworm) midgut display differential interaction with closely related Bacillus thuringiensis insecticidal proteins. Biochem. J. 2003, 370, 971–978. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Conde, J.; Muñoz-Garay, C.; Sánchez, J.; Miranda, R.; Zhuang, M.; Gill, S.S.; Soberón, M. Oligomerization triggers binding of a Bacillus thuringiensis Cry1Ab pore-forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains. Biochim. Biophys. Acta Biomembr. 2004, 1667, 38–46. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Adang, M.J. Characterization of a Cry1Ac-receptor alkaline phosphatase in susceptible and resistant Heliothis virescens larvae. Eur. J. Biochem. 2004, 271, 3127–3135. [Google Scholar] [CrossRef]
- Arenas, I.; Bravo, B.; Soberón, M.; Gómez, I. Role of alkaline phosphatase from Manduca sexta in the mechanism of action of Bacillus thuringiensis Cry1Ab toxin. J. Biol. Chem. 2010, 285, 12497–12503. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Navarrete, F.; Gómez, I.; Peña, G.; Bravo, A.; Soberón, M.A. Tenebrio molitor GPI-anchored alkaline phosphatase is involved in binding of Bacillus thuringiensis Cry3Aa to brush border membrane vesicles. Peptides 2013, 41, 81–86. [Google Scholar] [CrossRef]
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Phys. 2010, 104, 103–110. [Google Scholar] [CrossRef]
- Gahan, L.J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS Genet. 2010, 6, e1001248. [Google Scholar] [CrossRef]
- Park, Y.; Gonza´lez-Martı´nez, R.M.; Navarro-Cerrillo, G.; Chakroun, M.; Kim, Y.; Ziarsolo, P.; Blanca, J.; Cañizares, J.; Ferré, J.; Herrero, S. ABCC transporters mediate insect resistance to multiple Bt toxins revealed by bulk segregant analysis. BMC Biol. 2014, 12, 46. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Sui-Fai, L.; Reineke, A.; Williams, A.K.; Gordon, K.H.J. Insect resistance to Bacillus thuringiensis toxin Cry2Ab is conferred by mutations in an ABC transporter subfamily A protein. PLoS Genet. 2015, 11, e1005534. [Google Scholar] [CrossRef]
- McNall, R.J.; Adang, M.J. Identification of novel Bacillus thuringiensis Cry1Ac binding proteins in Manduca sexta midgut through proteomic analysis. Insect Biochem. Mol. Biol. 2003, 33, 999–1010. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Jurat-Fuentes, J.L.; McNall, R.J.; Andacht, T.; Adang, M.J. Identification of novel Cry1Ac binding proteins in midgut membranes from Heliothis virescens using proteomic analyses. Insect Biochem. Mol. Biol. 2007, 37, 189–201. [Google Scholar] [CrossRef]
- Xu, L.; Ferry, N.; Wang, Z.; Zhang, J.; Edwards, M.G.; Gatehouse, A.M.R.; He, K.L. A proteomic approach to study the mechanism of tolerance to Bt toxins in Ostrinia furnacalis larvae selected for resistance to Cry1Ab. Transgen. Res. 2013, 22, 1155–1166. [Google Scholar] [CrossRef]
- Ma, G.; Roberts, H.; Sarjan, M.; Featherstone, N.; Lahnstein, J.; Akhurst, R.; Schmidt, O. Is the mature endotoxin Cry1Ac from Bacillus thuringiensis inactivated by a coagulation reaction in the gut lumen of resistant Helicoverpa armigera larvae? Insect Biochem. Mol. Biol. 2005, 35, 729–739. [Google Scholar] [CrossRef]
- Liu, S.; Niu, H.; Xiao, T.; Xue, C.; Liu, Z.; Luo, W. Does phenoloxidase contributed to the resistance? Selection with butane-fipronil enhanced its activities from diamondback moths. Open Biochem. J. 2009, 3, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Cunha, F.M.; Wanderley-Teixeira, V.; Torres, J.B.; Álvaro, A.C.; Teixeira, T.J.S.; Alves, T.J.S.; Brayner, F.A. Impact of Bt cotton on the immune system and histology of the midgut of the fall armyworm Spodoptera frugiperda (Smith, J.E.) (Lepidoptera: Noctuidae). Anim. Biol. 2013, 63, 185–197. [Google Scholar] [CrossRef]
- Prabu, S.; Jing, D.; Shabbir, M.Z.; Yuan, W.; Wang, Z.; He, K.L. Contribution of phenoloxidase activation mechanism to Bt insecticidal protein resistance in Asian corn borer. Int. J. Biol. Macromol. 2010, 153, 88–99. [Google Scholar] [CrossRef]
- Shabbir, M.Z.; Zhang, T.; Prabu, S.; Wang, Y.; Wang, Z.; Bravo, A.; Soberón, M.; He, K.L. Identification of Cry1Ah-binding proteins through pull down and gene expression analysis in Cry1Ah-resistant and susceptible strains of Ostrinia furnacalis. Pestic. Biochem. Phys. 2020, 163, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Ren, Y.; Liu, Y.; Liang, G.; Song, F.; Bai, S.; Wang, J.; Wang, G. Overexpression of a novel Cry1Ie gene confers resistance to Cry1Ac-resistant cotton bollworm in transgenic lines of maize. Plant Cell Tissue Organ Cult. 2013, 115, 151–158. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, T.; Bai, S.; Wang, Z.; He, K.L. Evaluation of Bt corn with pyramided genes on efficacy and insect resistance management for the Asian corn borer in China. PLoS ONE 2016, 11, e0168442. [Google Scholar] [CrossRef]
- Shabbir, M.Z.; Quan, Y.; Wang, Z.; Bravo, A.; Soberón, M.; He, K.L. Characterization of the Cry1Ah resistance in Asian corn borer and its cross-resistance to other Bacillus thuringiensis toxins. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- He, K.L.; Wang, Z.Y.; Zhou, D.; Wen, L.; Song, Y. Evaluation of transgenic Bt corn for resistance to the Asian corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 2003, 96, 935–940. [Google Scholar] [CrossRef]
- Zhou, D.R.; He, K.L.; Wang, Z.Y.; Ye, Z.H.; Wen, L.P.; Gao, Y.X.; Song, Y.Y. Asian Corn Borer and Its Integrated Management; Golden Shield Press: Beijing, China, 1995; p. 102. (In Chinese) [Google Scholar]
- Zhuang, M.; Oltean, D.I.; Gómez, I.; Pullikuth, A.K.; Soberón, M.; Bravo, A.; Gill, S.S. Heliothis virescens and Manduca sexta lipid rafts are involved in Cry1A toxin binding to the midgut epithelium and subsequent pore formation. J. Biol. Chem. 2002, 277, 13863–13872. [Google Scholar] [CrossRef]
- Ferré, J.; Van Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein vip3a differs from that of Cry1ab δ-endotoxin. Biochem. Pharmacol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, H.A.A.; González-Cabrera, J.; Ferré, J.; Flannagan, R.; Siegfried, B.D. Analyses of Cry1Ab binding in resistant and susceptible strains of the European corn borer, Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae). Appl. Environ. Microbiol. 2006, 72, 5318–5324. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.J.G.; Siqueira, H.A.A.; Zhuang, M.; Storer, N.P.; Siegfried, B.D. Measurements of Cry1F binding and activity of luminal gut proteases in susceptible and Cry1F resistant Ostrinia nubilalis larvae (Lepidoptera: Crambidae). J. Invertebr. Pathol. 2010, 103, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Crespo, A.L.B.; Rodrigo-simón, A.; Siqueira, H.A.A.; Pereira, E.J.G.; Ferré, J.; Siegfried, B.D. Cross-resistance and mechanism of resistance to Cry1Ab toxin from Bacillus thuringiensis in a field-derived strain of European corn borer, Ostrinia nubilalis. J. Invertebr. Pathol. 2011, 107, 185–192. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2013, 6, 17–26. [Google Scholar] [CrossRef]
- Liang, Y.; Patel, S.S.; Dean, D.H. Irreversible binding kinetics of Bacillus thuringiensis CryIA δ-endotoxins to gypsy moth brush border membrane vesicles is directly correlated to toxicity. J. Biol. Chem. 1995, 270, 24719–24724. [Google Scholar] [CrossRef]
- Wu, S.J.; Dean, D.H. Functional Significance of Loops in The Receptor Binding Domain of Bacillus thuringiensis CryIIIA δ-Endotoxin. J. Mol. Biol. 1996, 255, 628–640. [Google Scholar] [CrossRef]
- Wolfersberger, M.G. The toxicity of two Bacillus thuringiensis δ-endotoxins to gypsy moth larvae is inversely related to the affinity of binding sites on midgut brush border membranes for the toxins. Experientia 1990, 46, 475–477. [Google Scholar] [CrossRef]
- Lee, M.K.; Dean, D.H. Inconsistencies in determining Bacillus thuringiensis toxin binding sites relationship by comparing competition assays with ligand blotting. Biochem. Biophys. Res. Commun. 1996, 220, 575–580. [Google Scholar] [CrossRef]
- Sangadala, S.; Walters, F.S.; English, L.H.; Adang, M.J. A mixture of Manduca sexta aminopeptidase and phosphatase enhances Bacillus thuringiensis insecticidal CryIA(c) toxin binding and 86Rb-K1 efflux in vitro. J. Biol. Chem. 1994, 269, 10088–10092. [Google Scholar]
- Masson, L.; Lu, J.Y.; Mazza, A.; Brousseau, R.; Adang, M.J. The CryIA(c) receptor purified from Manduca sexta displays multiple specificities. J. Biol. Chem. 1995, 270, 20309–20315. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.L.; Lu, Y.J.; Söhnlein, P.; Brousseau, R.; Laprade, R.; Masson, L.; Adang, M.J. Ion channels formed in planar lipid bilayers by Bacillus thuringiensis toxins in the presence of Manduca sexta midgut receptors. FEBS Lett. 1997, 412, 270–276. [Google Scholar] [CrossRef]
- Keeton, T.P.; Francis, B.R.; Maaty, W.S.; Bulla, L.A., Jr. Effects of midgut-protein-preparative and ligand binding procedures on the toxin binding characteristics of BT-R1, a common high-affinity receptor in Manduca sexta for Cry1A Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 1998, 64, 2158–2165. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Karumbaiah, L.; Jakka, S.R.; Ning, C.; Liu, C.; Wu, K.; Jackson, J.; Gould, F.; Blanco, C.; Portilla, M.; et al. Reduced levels of membrane-bound alkaline phosphatase are common to lepidopteran strains resistant to cry toxins from Bacillus thuringiensis. PLoS ONE 2001, 6, e17606. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, C.; Xiao, Y.; Zhang, D.; Zhang, Y.; Li, X.; Tabashnik, B.E.; Wu, K. A toxin-binding alkaline phosphatase fragment synergizes Bt toxin Cry1Ac against susceptible and resistant Helicoverpa armigera. PLoS ONE 2015, 10, e0126288. [Google Scholar] [CrossRef]
- Gahan, L.J.; Gould, F.; Heckel, D.G. Identification of a gene associated with Bt resistance in Heliothis virescens. Science 2001, 293, 857–860. [Google Scholar] [CrossRef]
- Baxter, S.W.; Badenes-Pérez, F.R.; Morrison, A.; Vogel, H.; Crickmore, N.; Kain, W.; Wang, P.; Heckel, W.G.; Jiggins, C.D. Parallel evolution of Bacillus thuringiensis toxin resistance in lepidoptera. Genetics 2011, 189, 675–679. [Google Scholar] [CrossRef]
- Aronson, A.I.; Wu, D.; Zhang, C. Mutagenesis of specificity and toxicity regions of a Bacillus thuringiensis protoxin gene. J. Bacteriol. 1995, 177, 4059–4065. [Google Scholar] [CrossRef]
- Atsumi, S.; Miyamato, K.; Yamamoto, K.; Narukawa, J.; Kawai, S.; Sezutsu, H.; Kobayashi, I.; Uchino, K.; Tamura, T.; Mita, K.; et al. Single amino acid mutation in an ATP-binding cassette transporter causes resistance to Bt toxin Cry1Ab in the silkworm, Bombyx mori. Proc. Natl Acad. Sci. USA 2012, 109, 1591–1598. [Google Scholar] [CrossRef]
- Valadez-Lira, J.A.; Alcocer-Gonzalez, J.M.; Damas, G.; Nuñez-Mejía, G.; Oppert, B.; Rodriguez-Padilla, C.; Tamez-Guerra, P. Comparative evaluation of phenoloxidase activity in different larval stages of four lepidopteran pests after exposure to Bacillus thuringiensis. J. Insect Sci. 2012, 12, 1–11. [Google Scholar] [CrossRef]
- Ashida, M.; Dohke, K. Activation of pro-phenol oxidase by the activating enzyme of the silkworm, Bombyx mori. Insect Biochem. 1980, 10, 37–47. [Google Scholar] [CrossRef]
- Nappi, A.J.; Vass, E. Cytotoxic reactions associated with insect immunity. Adv. Exp. Med. Biol. 2001, 484, 329–348. [Google Scholar] [PubMed]
- Bidla, G.; Hauling, T.; Dushay, M.S.; Theopold, U. Activation of insect phenoloxidase after injury: Endogenous versus foreign elicitors. J. Innate Immun. 2009, 1, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Wolfersberger, M.; Lüthy, P.; Maurer, A.; Parenti, P.; Sacchi, F.; Giordana, B.; Hanozet, G. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Phys. A Comp. Phys. 1987, 86, 301–308. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rodrigo-Simón, A.; Caccia, S.; Ferré, J. Bacillus thuringiensis Cry1Ac toxin-binding and pore-forming activity in brush border membrane vesicles prepared from anterior and posterior midgut regions of lepidopteran larvae. Appl. Environ. Microbiol. 2008, 74, 1710–1716. [Google Scholar] [CrossRef]


{kind=link}
| Enzymatic Assays (μmol/min/mL) | ACB-BtS | ACB-AhR | ||
|---|---|---|---|---|
| Midgut Homogenate | BBMV | Midgut Homogenate | BBMV | |
| APN | 16.92 | 52.40 | 26.13 | 98.45 |
| ALP | 9.42 | 32.42 | 16.68 | 62.15 |
| ACB-BtS Midgut Fractions | ACB-AhR Midgut Fractions | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fraction | Score | Accession Number | Protein Description | Sequence Coverage % | Fraction | Score | Accession Number | Protein Description | Sequence Coverage % |
| S1 | 65.30 | tr|B5A8 × 9 | Aminopeptidase | 7.2 | R1 | 22.56 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 9.6 |
| 20.09 | tr|D2Y440 | Actin | 19.9 | 19.68 | tr|E5LEV4 | Heat shock 70 | 4.7 | ||
| 17.09 | tr|Q8WB29 | Cytochrome c oxidase subunit 3 | 2.7 | 15.00 | tr|A0A0F7QEA3 | Actin | 20.4 | ||
| 16.87 | tr|A0A0A7BYG4 | Elongation factor 1 alpha (Fragment) | 7.6 | 21.40 | tr|B5A8X9 | Aminopeptidase | 1.2 | ||
| 15.68 | tr|E5LEV4 | Heat shock 70 kDa | 3.0 | 6.05 | tr|A0A1B4ZBI6 | Odorant binding protein | 5.9 | ||
| 14.34 | tr|B5A8Y0 | Aminopeptidase | 4.1 | 3.26 | tr|Q2PQR1 | Ubiquitin | 10.6 | ||
| 12.45 | tr|Q8W7D1 | ATP synthase subunit a | 3.4 | 2.90 | tr|A0A0A7DWF3 | Arginine kinase | 2.0 | ||
| 6.48 | tr|E3W6T9 | V-ATPase subunit A | 3.9 | 2.44 | tr|A0A0E4B3V9 | Odorant receptor | 4.2 | ||
| 5.28 | tr|A0A0A7BYG7 | Glyceraldehyde-3-phosphate dehydrogenase | 8.6 | 59.08 | tr|A7LIA2 | Cadherin-like protein | 3.7 | ||
| 5.10 | tr|A0A0F7QEE5 | Carboxylic ester hydrolase | 1.3 | 11.56 | tr|A0A0B6VK98 | Uncharacterized protein | 2.0 | ||
| 4.66 | tr|Q8WB23 | Cytochrome | 2.4 | 6.99 | tr|A0A0F7QIG2 | Ionotropic receptor | 1.1 | ||
| 3.97 | tr|A0A0F7QIG2 | Ionotropic receptor | 2.4 | 1.57 | tr|A0A1Q1MKI0 | Prophenoloxidase PPO1b | 4.2 | ||
| 3.84 | tr|Q2PQR1 | Ubiquitin | 24.3 | 13.74 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 14.7 | ||
| 3.69 | tr|A0A1B4ZBI6 | Odorant binding protein | 6.0 | 12.40 | tr|Q8W7D1 | synthase subunit a | 3.1 | ||
| 2.95 | tr|A0A0A7BYS7 | Glutathione-S transferase (Fragment) | 4.3 | 10.17 | tr|Q8WB24 | NADH dehydrogenase subunit 6 | 5.6 | ||
| 2.90 | tr|A0A0A7DWF3 | Arginine kinase | 2.0 | 9.14 | tr|Q8WB23 | Cytochrome b | 9.4 | ||
| 2.34 | tr|A0A0A7BYM5 | Glutathione-S transferase | 6.0 | 8.85 | tr|Q2V6H3 | Chitin synthase | 0.5 | ||
| 2.13 | tr|E0XJK4 | Heat shock protein 90 | 1.0 | 8.45 | tr|A0A0F7QJX1 | Carboxylic ester hydrolase | 2.0 | ||
| 7.59 | tr|A0A1Q1MKI5 | Prophenoloxidase PPO1a | 3.1 | ||||||
| 1.65 | tr|A0A0B6VK98 | Uncharacterized protein | 2.0 | ||||||
| 1.44 | tr|B4YIR0 | Chitin synthase | 2.2 | ||||||
| S2 | 240.08 | tr|D0UYB1 | Aminopeptidase | 44.2 | R2 | 64.86 | tr|B5A8X9 | Aminopeptidase | 15.0 |
| 181.29 | tr|A7YAH7 | Aminopeptidase | 27.4 | 45.15 | tr|A0A0S2C6J0 | Actin | 14.1 | ||
| 123.85 | tr|B5A8Y0 | Aminopeptidase | 19.9 | 59.08 | tr|A7LIA2 | Cadherin-like protein | 3.2 | ||
| 49.61 | tr|A7LIA2 | Cadherin-like protein | 7.6 | 28.02 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 7.1 | ||
| 29.97 | tr|E3W6T9 | V-ATPase subunit A | 17.7 | 6.46 | tr|B5A8X9 | Aminopeptidase | 1.5 | ||
| 21.42 | tr|Q7K403 | Acyl-CoA delta-9 desaturase | 5.1 | 17.85 | tr|A0A0A7BYG7 | Glyceraldehyde-3-phosphate dehydrogenase | 4.5 | ||
| 18.31 | tr|A5JJU1 | Aminopeptidase | 23.9 | 14.97 | tr|A7YAH7 | Aminopeptidase | 2.3 | ||
| 14.21 | tr|E5LEV4 | Heat shock 70 kDa | 2.8 | 13.21 | tr|E5LEV4 | Heat shock 70 kDa | 1.7 | ||
| 14.18 | tr|A0A0S2C6J0 | Actin | 18.5 | 11.56 | tr|A0A0B6VK98 | Uncharacterized protein | 2.0 | ||
| 6.24 | tr|Q8WB29 | Cytochrome c oxidase subunit | 5.3 | 7.31 | tr|A0A0A7DWF3 | Arginine kinase | 2.0 | ||
| 5.90 | tr|D0UYB2 | Aminopeptidase | 40.2 | 6.72 | tr|Q2PQR1 | Ubiquitin | 11.8 | ||
| 5.18 | tr|A0A0F7QEE5 | Carboxylic ester hydrolase | 1.3 | 5.95 | tr|L7QRW6 | Period | 2.2 | ||
| 2.88 | tr|M4Q143 | NADH-ubiquinone oxidoreductase | 3.1 | 5.87 | tr|A0A1B2AQF4 | Trehalase | 1.1 | ||
| 1.70 | tr|Q2PQR1 | Ubiquitin | 28.9 | 5.77 | tr|E3W6T9 | V-ATPase subunit A | 1.5 | ||
| 1.68 | tr|A0A0A7BYG7 | Glyceraldehyde-3-phosphate dehydrogenase | 2.1 | 5.73 | tr|A0A0F7QEC7 | Aldehyde oxidase | 1.4 | ||
| 1.60 | tr|A0A1B4ZBI6 | Odorant binding protein 9 | 5.9 | 5.71 | tr|A0A1Q1MKI5 | Prophenoloxidase PPO1a | 3.6 | ||
| 1.30 | tr|M1RM07 | Elongation factor-1 alpha | 14.1 | 5.68 | tr|A0A0E4B3V9 | Odorant receptor | 2.0 | ||
| 1.04 | tr|Q8WB31 | Cytochrome c oxidase subunit 1 | 1.6 | ||||||
| 0.95 | tr|A0A0E4B5I4 | Odorant receptor | 1.7 | ||||||
| 0.57 | tr|A0A0A7BYS7 | Glutathione-S transferase | 3.2 | ||||||
| 0.49 | tr|L7QRW6 | Period | 2.2 | ||||||
| 0.49 | tr|A0A0E3VLQ5 | Odorant receptor | 2.3 | ||||||
| 0.45 | tr|M1RTC5 | Elongation factor-1 alpha | 16.7 | ||||||
| S3 | 323.31 | tr|E3W6T9 | V-ATPase subunit A | 71.5 | R3 | 244.29 | tr|B5A8X9 | Aminopeptidase | 28.9 |
| 95.44 | tr|B5A8X9 | Aminopeptidase | 21.7 | 66.82 | tr|A7YAH7 | Aminopeptidase | 9.2 | ||
| 50.77 | tr|E5LEV4 | Heat shock 70 kDa | 28.2 | 65.03 | tr|E3W6T9 | V-ATPase subunit A | 14.3 | ||
| 37.45 | tr|A7YAH7 | Aminopeptidase | 12.2 | 54.05 | tr|B5A8Y0 | Aminopeptidase | 10.6 | ||
| 30.53 | tr|E0XJK4 | Heat shock protein 90 | 17.3 | 27.18 | tr|D2Y440 | Actin | 30.6 | ||
| 17.39 | tr|B5A8Y0 | Aminopeptidase | 8.0 | 25.37 | tr|D0UYB2 | Aminopeptidase | 7.7 | ||
| 14.82 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 14.7 | 21.40 | tr|B2LS41 | Aminopeptidase | 26.9 | ||
| 9.53 | tr|A0A1L7B974 | Serpin 5 | 12.2 | 21.13 | tr|A0A0S2C6J0 | Actin | 23.9 | ||
| 12.66 | tr|A0A0F7QJX1 | Carboxylic ester hydrolase | 9.6 | 14.45 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 7.1 | ||
| 9.12 | tr|B2LS41 | Aminopeptidase | 21.7 | 13.98 | tr|E5LEV4 | Heat shock 70 kDa | 2.8 | ||
| 7.98 | tr|D0UYB2 | Aminopeptidase | 5.9 | 13.91 | tr|A0A0A0YWU7 | Arginine kinase | 5.6 | ||
| 7.98 | tr|D2Y440 | Actin (Fragment) | 19.1 | 6.62 | tr|A0A1Q1MKI0 | Prophenoloxidase PPO1b | 1.7 | ||
| 7.83 | tr|A0A0A7DWF3 | Arginine kinase | 8.5 | 6.29 | tr|A0A0F7QIG2 | Ionotropic receptor | 1.1 | ||
| 7.72 | tr|Q06GJ0 | Beta-hexosaminidase | 2.7 | 6.09 | tr|Q8WB29 | Cytochrome c oxidase subunit 3 | 2.7 | ||
| 4.99 | tr|M1RTC5 | Elongation factor-1 alpha | 22.3 | 5.71 | tr|J7FBQ4 | Odorant receptor | 2.4 | ||
| 4.83 | tr|A0A0S2C6J0 | Actin | 14.1 | 5.60 | tr|M1RTC5 | Elongation factor-1 alpha | 5.6 | ||
| 3.62 | tr|M1RM07 | Elongation factor-1 alpha | 19.4 | 5.60 | tr|A0A0F7QEJ4 | Sensory neuron membrane protein 1 | 1.5 | ||
| 2.96 | tr|Q2PQR1 | Ubiquitin | 24.9 | 2.43 | tr|A0A1L7B973 | Serine proteinase inhibitor 2 | 17.5 | ||
| 2.64 | tr|Q8WB31 | Cytochrome c oxidase subunit 1 | 1.6 | 14.59 | tr|A0A1L7B974 | Serpin 5 | 12.4 | ||
| 1.84 | tr|G1JT78 | Alkaline phosphatase | 1.3 | ||||||
| S4 | 258.25 | tr|E3W6T9 | V-ATPase subunit A | 49.2 | R4 | 231.15 | tr|E3W6T9 | V-ATPase subunit A | 27.1 |
| 49.45 | tr|B5A8X9 | Aminopeptidase | 14.4 | 164.37 | tr|B2LS41 | Aminopeptidase | 16.6 | ||
| 24.74 | tr|A7YAH7 | Aminopeptidase | 8.7 | 48.18 | tr|D2Y440 | Actin | 30.2 | ||
| 23.70 | tr|A0A1L7B973 | Serine proteinase inhibitor 2 | 14.6 | 46.51 | tr|B5A8X9 | Aminopeptidase | 16.8 | ||
| 19.10 | tr|A0A0F7QJX1 | Carboxylic ester hydrolase | 13.3 | 43.97 | tr|A0A0S2C6J0 | Actin | 28.3 | ||
| 17.18 | tr|A0A0F7QIF1 | Carboxylic ester hydrolase | 15.0 | 37.32 | tr|E0XJK4 | Heat shock protein 90 | 6.3 | ||
| 10.56 | tr|E0XJK4 | Heat shock protein 90 | 7.8 | 30.01 | tr|E5LEV4 | Heat shock 70 kDa | 6.3 | ||
| 6.84 | tr|A0A0S2C6J0 | Actin | 17.9 | 24.14 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 7.1 | ||
| 6.06 | tr|A0A0B6VQ49 | Uncharacterized protein | 42.7 | 22.07 | tr|F6MEP1 | Storage protein | 6.1 | ||
| 5.24 | tr|D2Y440 | Actin | 19.1 | 8.94 | tr|Q8WB29 | Cytochrome c oxidase subunit 3 | 2.7 | ||
| 4.90 | tr|A0A0A7BYG4 | Elongation factor 1 alpha | 11.2 | 8.02 | tr|M1RTC5 | Elongation factor-1 alpha | 9.4 | ||
| 4.58 | tr|M4T4G3 | Ryanodine receptor | 1.3 | 7.72 | tr|A0A0F7QEE5 | Carboxylic ester hydrolase | 1.3 | ||
| 4.37 | tr|A0A0A7DWF3 | Arginine kinase | 8.5 | 6.74 | tr|A0A1B4ZBI6 | Odorant binding protein 9 | 5.9 | ||
| 4.23 | tr|B5A8Y0 | Aminopeptidase | 2.2 | 6.65 | tr|A0A0A7BYG7 | Glyceraldehyde-3-phosphate dehydrogenase | 2.1 | ||
| 3.00 | tr|M1RM07 | Elongation factor-1 alpha | 14.1 | 6.40 | tr|A0A1Q1MKI5 | Prophenoloxidase PPO1a | 1.2 | ||
| 1.81 | tr|Q8WB29 | Cytochrome c oxidase subunit 3 | 2.7 | 6.28 | tr|A2TIK8 | Integrin beta | 1.1 | ||
| 1.67 | tr|E5LEV4 | Heat shock 70 kDa | 1.1 | 5.98 | tr|A0A0A7BYS7 | Glutathione-S transferase | 3.2 | ||
| 1.48 | tr|Q2V6H3 | Chitin synthase | 3.0 | ||||||
| 1.33 | tr|D2KWQ3 | Odorant receptor | 2.5 | ||||||
| 1.20 | tr|G1JT78 | Alkaline phosphatase | 1.3 | ||||||
| 1.08 | tr|B2LS41 | Aminopeptidase | 12.7 | ||||||
| 0.89 | tr|A0A1L7B974 | Serpin 5 | 22.9 | ||||||
| 0.87 | tr|A0A1U8ZSW0 | Storage protein | 1.2 | ||||||
| 0.85 | tr|A0A0F7QEC9 | Carboxylesterase | 1.4 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prabu, S.; Shabbir, M.Z.; Wang, Z.; He, K. Analysis of Cry1Ah Toxin-Binding Reliability to Midgut Membrane Proteins of the Asian Corn Borer. Toxins 2020, 12, 418. https://doi.org/10.3390/toxins12060418
Prabu S, Shabbir MZ, Wang Z, He K. Analysis of Cry1Ah Toxin-Binding Reliability to Midgut Membrane Proteins of the Asian Corn Borer. Toxins. 2020; 12(6):418. https://doi.org/10.3390/toxins12060418
Chicago/Turabian StylePrabu, Sivaprasath, Muhammad Zeeshan Shabbir, Zhenying Wang, and Kanglai He. 2020. "Analysis of Cry1Ah Toxin-Binding Reliability to Midgut Membrane Proteins of the Asian Corn Borer" Toxins 12, no. 6: 418. https://doi.org/10.3390/toxins12060418
APA StylePrabu, S., Shabbir, M. Z., Wang, Z., & He, K. (2020). Analysis of Cry1Ah Toxin-Binding Reliability to Midgut Membrane Proteins of the Asian Corn Borer. Toxins, 12(6), 418. https://doi.org/10.3390/toxins12060418