Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily


Abstract
1. Introduction
2. Structural Features of Cysteine-Rich Secretory Proteins (CRISPs)
2.1. CAP/PR-1 Domain
2.2. Hinge Region
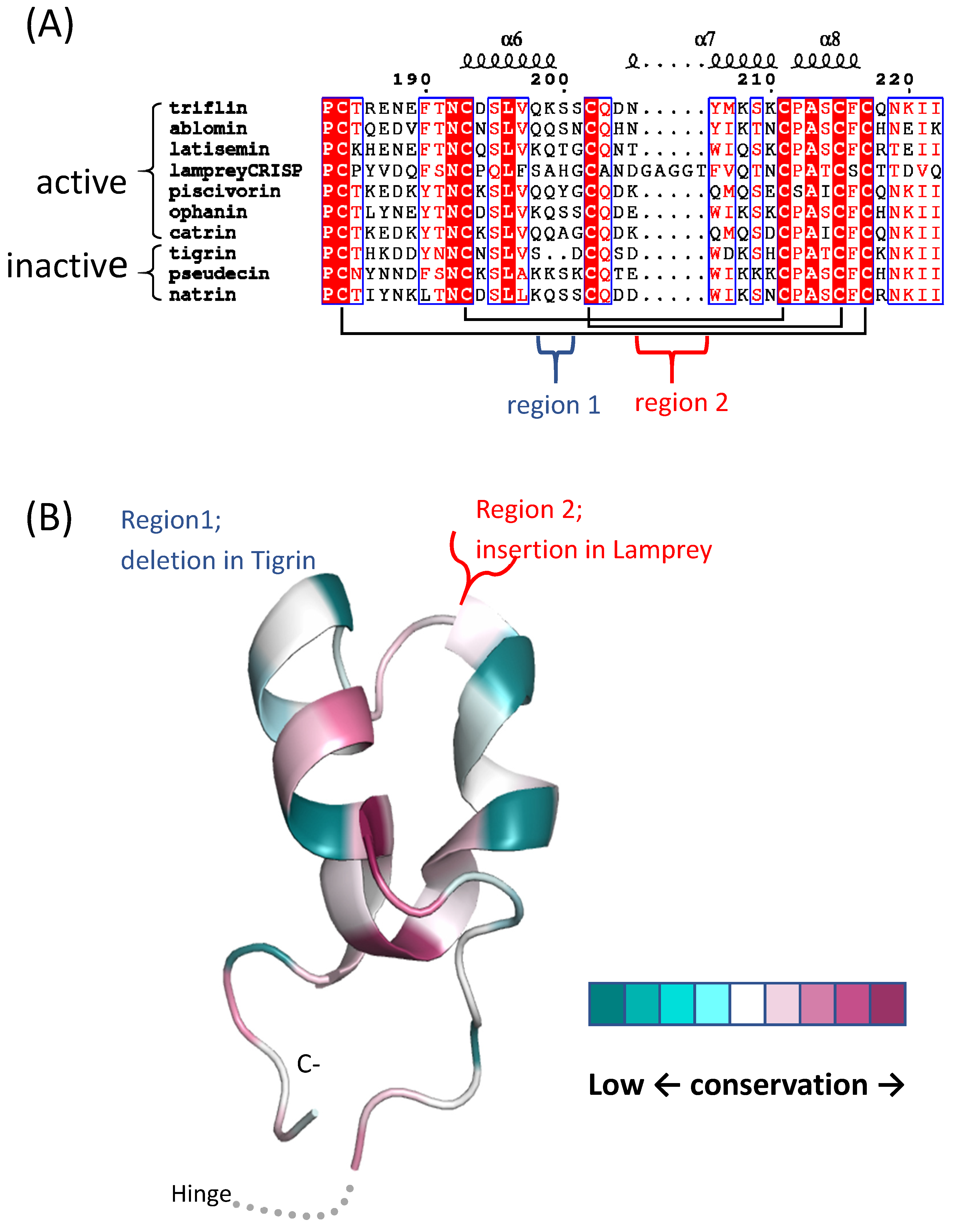
2.3. Cysteine-Rich Domain (CRD)/Ion Channel Regulatory (ICR) Domain
3. CRISP Co-Factors
4. Proteins That Bind to CRISPs
5. Isolation and Characterization of Snake Venom CRISPs
5.1. Ancestral CRISP Activity
5.2. Myotoxicity
5.3. Ion Channel Inhibition
5.4. Anti-Protozoal Activity
5.5. Anti-Angiogenic Activity
5.6. Vascular Permeability Regulator
5.7. Inflammation Regulator and Protease Activity
6. Functional Sites Identified in CRISPs
6.1. Potential Functional Sites Responsible for Protease Activity
6.2. Potential Domains and Functional Sites Responsible for Ion Channel Inhibition
7. svCRISP Evolution
8. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cantacessi, C.; Gasser, R.B. SCP/TAPS proteins in helminths–Where to from now? Mol. Cell. Probes 2012, 26, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Van Kammen, A. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. ‘Samsun’ and ‘Samsun NN’. Virology 1970, 40, 199–211. [Google Scholar] [CrossRef]
- Alexander, D.; Goodman, R.M.; Gut-Rella, M.; Glascock, C.; Weymann, K.; Friedrich, L.; Maddox, D.; Ahl-Goy, P.; Luntz, T.; Ward, E. Increased tolerance to two oomycete pathogens in transgenic tobacco expressing pathogenesis-Related protein 1a. Proc. Natl. Acad. Sci. USA 1993, 90, 7327–7331. [Google Scholar] [CrossRef] [PubMed]
- Kiba, A.; Nishihara, M.; Nakatsuka, T.; Yamamura, S. Pathogenesis-Related protein 1 homologue is an antifungal protein in Wasabia japonica leaves and confers resistance to Botrytis cinerea in transgenic tobacco. Plant Biotechnol. 2007, 24, 247–253. [Google Scholar] [CrossRef]
- Sarowar, S.; Kim, Y.J.; Kim, E.N.; Kim, K.D.; Hwang, B.K.; Islam, R.; Shin, J.S. Overexpression of a pepper basic pathogenesis-Related protein 1 gene in tobacco plants enhances resistance to heavy metal and pathogen stresses. Plant Cell Rep. 2005, 24, 216–224. [Google Scholar] [CrossRef]
- Shin, S.H.; Park, J.-H.; Kim, M.-J.; Kim, H.-J.; Oh, J.S.; Choi, H.K.; Jung, H.W.; Chung, Y.S. An Acidic PATHOGENESIS-RELATED1 Gene of Oryza grandiglumis is Involved in Disease Resistance Response Against Bacterial Infection. Plant Pathol. J. 2014, 30, 208–214. [Google Scholar] [CrossRef]
- Carr, J.P.; Beachy, R.N.; Klessig, D.F. Are the PR1 proteins of tobacco involved in genetically engineered resistance to TMV? Virology 1989, 169, 470–473. [Google Scholar] [CrossRef]
- Hon, W.C.; Griffith, M.; Mlynarz, A.; Kwok, Y.C.; Yang, D. Antifreeze Proteins in Winter Rye Are Similar to Pathogenesis-Related Proteins. Plant Physiol. 1995, 109, 879–889. [Google Scholar] [CrossRef]
- Zeier, J.; Pink, B.; Mueller, M.; Berger, S. Light conditions influence specific defence responses in incompatible plant-pathogen interactions: Uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta 2004, 219. [Google Scholar] [CrossRef]
- Seo, P.J.; Lee, A.-K.; Xiang, F.; Park, C.-M. Molecular and Functional Profiling of Arabidopsis Pathogenesis-Related Genes: Insights into Their Roles in Salt Response of Seed Germination. Plant Cell Physiol. 2008, 49, 334–344. [Google Scholar] [CrossRef]
- Liu, W.-X.; Zhang, F.-C.; Zhang, W.-Z.; Song, L.-F.; Wu, W.-H.; Chen, Y.-F. Arabidopsis Di19 Functions as a Transcription Factor and Modulates PR1, PR2, and PR5 Expression in Response to Drought Stress. Mol. Plant 2013, 6, 1487–1502. [Google Scholar] [CrossRef] [PubMed]
- Kothari, K.S.; Dansana, P.K.; Giri, J.; Tyagi, A.K. Rice Stress Associated Protein 1 (OsSAP1) Interacts with Aminotransferase (OsAMTR1) and Pathogenesis-Related 1a Protein (OsSCP) and Regulates Abiotic Stress Responses. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Memelink, J.; Linthorst, H.J.M.; Schilperoort, R.A.; Hoge, J.H.C. Tobacco genes encoding acidic and basic isoforms of pathogenesis-Related proteins display different expression patterns. Plant Mol. Biol. 1990, 14, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef] [PubMed]
- King, T.P.; Spangfort, M.D. Structure and Biology of Stinging Insect Venom Allergens. Int. Arch. Allergy Immunol. 2000, 123, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Andersen, J.F.; Francischetti, I.M.B.; Valenzuela, J.G.; Schwan, T.G.; Pham, V.M.; Garfield, M.K.; Hammer, C.H.; Ribeiro, J.M.C. Comparative sialomics between hard and soft ticks: Implications for the evolution of blood-Feeding behavior. Insect Biochem. Mol. Biol. 2008, 38, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Charlab, R.; Valenzuela, J.G.; Rowton, E.D.; Ribeiro, J.M.C. Toward an understanding of the biochemical and pharmacological complexity of the saliva of a hematophagous sand fly Lutzomyia longipalpis. Proc. Natl. Acad. Sci. USA 1999, 96, 15155–15160. [Google Scholar] [CrossRef]
- Calvo, E.; Dao, A.; Pham, V.M.; Ribeiro, J.M.C. An insight into the sialome of Anopheles funestus reveals an emerging pattern in anopheline salivary protein families. Insect Biochem. Mol. Biol. 2007, 37, 164–175. [Google Scholar] [CrossRef]
- Henriksen, A.; King, T.P.; Mirza, O.; Monsalve, R.I.; Meno, K.; Ipsen, H.; Larsen, J.N.; Gajhede, M.; Spangfort, M.D. Major venom allergen of yellow jackets, Ves v 5: Structural characterization of a pathogenesis-Related protein superfamily. Proteins Struct. Funct. Genet. 2001, 45, 438–448. [Google Scholar] [CrossRef]
- Müller, U.R.; Johansen, N.; Petersen, A.B.; Fromberg-Nielsen, J.; Haeberli, G. Hymenoptera venom allergy: Analysis of double positivity to honey bee and Vespula venom by estimation of IgE antibodies to species-Specific major allergens Api m1 and Ves v5. Allergy 2009, 64, 543–548. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C.; Francischetti, I.M.B. Role of arthropod saliva in blood feeding: Sialome and Post-Sialome Perspectives. Annu. Rev. Entomol. 2003, 48, 73–88. [Google Scholar] [CrossRef] [PubMed]
- dos Santos-Pinto, J.R.A.; dos Santos, L.D.; Andrade Arcuri, H.; Castro, F.M.; Kalil, J.E.; Palma, M.S. Using Proteomic Strategies for Sequencing and Post-Translational Modifications Assignment of Antigen-5, a Major Allergen from the Venom of the Social Wasp Polybia paulista. J. Proteome Res. 2014, 13, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Assumpção, T.C.F.; Ma, D.; Schwarz, A.; Reiter, K.; Santana, J.M.; Andersen, J.F.; Ribeiro, J.M.C.; Nardone, G.; Yu, L.L.; Francischetti, I.M.B. Salivary Antigen-5/CAP Family Members Are Cu2+-Dependent Antioxidant Enzymes That Scavenge O2− and Inhibit Collagen-induced Platelet Aggregation and Neutrophil Oxidative Burst. J. Biol. Chem. 2013, 288, 14341–14361. [Google Scholar] [CrossRef] [PubMed]
- Cameo, M.S.; Blaquier, J.A. Androgen-Controlled specific proteins in rat epididymis. J. Endocrinol. 1976, 69, 47–55. [Google Scholar] [CrossRef]
- Kierszenbaum, A.L.; Lea, O.; Petrusz, P.; French, F.S.; Tres, L.L. Isolation, culture, and immunocytochemical characterization of epididymal epithelial cells from pubertal and adult rats. Proc. Natl. Acad. Sci. USA 1981, 78, 1675–1679. [Google Scholar] [CrossRef]
- Kratzschmar, J.; Haendler, B.; Eberspaecher, U.; Roosterman, D.; Donner, P.; Schleuning, W.-D. The Human Cysteine-Rich Secretory Protein (CRISP) Family. Primary Structure and Tissue Distribution of CRISP-1, CRISP-2 and CRISP-3. Eur. J. Biochem. 1996, 236, 827–836. [Google Scholar] [CrossRef]
- Udby, L.; Cowland, J.B.; Johnsen, A.H.; Sørensen, O.E.; Borregaard, N.; Kjeldsen, L. An ELISA for SGP28/CRISP-3, a cysteine-Rich secretory protein in human neutrophils, plasma, and exocrine secretions. J. Immunol. Methods 2002, 263, 43–55. [Google Scholar] [CrossRef]
- Koppers, A.J.; Reddy, T.; O′Bryan, M.K. The role of cysteine-Rich secretory proteins in male fertility. Asian J. Androl. 2011, 13, 111–117. [Google Scholar] [CrossRef]
- Gibbs, G.M.; Roelants, K.; O′Bryan, M.K. The CAP Superfamily: Cysteine-Rich Secretory Proteins, Antigen 5, and Pathogenesis-Related 1 Proteins—Roles in Reproduction, Cancer, and Immune Defense. Endocr. Rev. 2008, 29, 865–897. [Google Scholar] [CrossRef]
- Gibbs, G.M.; O′Bryan, M.K. Cysteine rich secretory proteins in reproduction and venom. Soc. Reprod. Fertil. Suppl. 2007, 65, 261–267. [Google Scholar]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-Rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Milne, T.J.; Abbenante, G.; Tyndall, J.D.A.; Halliday, J.; Lewis, R.J. Isolation and Characterization of a Cone Snail Protease with Homology to CRISP Proteins of the Pathogenesis-Related Protein Superfamily. J. Biol. Chem. 2003, 278, 31105–31110. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.; Jackson, T.N.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A proteomics and transcriptomics investigation of the venom from the barychelid spider Trittame loki (brush-Foot trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gutierrez, T.; Peguero-Sanchez, E.; Cevallos, M.A.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. A Deeper Examination of Thorellius atrox Scorpion Venom Components with Omic Techonologies. Toxins 2017, 9, 399. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Mita, M.; Takahashi, Y.; Matsushima, A.; Watanabe, Y.G.; Hirano, S.; Odani, S. Novel cysteine-Rich secretory protein in the buccal gland secretion of the parasitic lamprey, Lethenteron japonicum. Biochem. Biophys. Res. Commun. 2007, 358, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Casewell, N.R.; Wuster, W.; Vidal, N.; Young, B.; Jackson, T.N. The structural and functional diversification of the Toxicofera reptile venom system. Toxicon 2012, 60, 434–448. [Google Scholar] [CrossRef]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.R.; Kuruppu, S.; Fung, K.; Blair Hedges, S.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef]
- Barua, A.; Mikheyev, A.S. Many Options, Few Solutions: Over 60 My Snakes Converged on a Few Optimal Venom Formulations. Mol. Biol. Evol. 2019, 36, 1964–1974. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Morrissette, J.; Krätzschmar, J.; Haendler, B.; el-Hayek, R.; Mochca-Morales, J.; Martin, B.M.; Patel, J.R.; Moss, R.L.; Schleuning, W.D.; Coronado, R. Primary structure and properties of helothermine, a peptide toxin that blocks ryanodine receptors. Biophys. J. 1995, 68, 2280–2288. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Heyborne, W.H. Cysteine-Rich secretory proteins in reptile venoms. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2010; pp. 325–334. [Google Scholar]
- Niderman, T.; Genetet, I.; Bruyere, T.; Gees, R.; Stintzi, A.; Legrand, M.; Fritig, B.; Mosinger, E. Pathogenesis-Related PR-1 Proteins Are Antifungal (Isolation and Characterization of Three 14-Kilodalton Proteins of Tomato and of a Basic PR-1 of Tobacco with Inhibitory Activity against Phytophthora infestans). Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Anklesaria, J.H.; Pandya, R.R.; Pathak, B.R.; Mahale, S.D. Purification and characterization of CRISP-3 from human seminal plasma and its real-Time binding kinetics with PSP94. J. Chromatogr. B 2016, 1039, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Crest, M.; Jacquet, G.; Gola, M.; Zerrouk, H.; Benslimane, A.; Rochat, H.; Mansuelle, P.; Martin-Eauclaire, M.F. Kaliotoxin, a novel peptidyl inhibitor of neuronal BK-type Ca(2+)-activated K+ channels characterized from Androctonus mauretanicus mauretanicus venom. J. Biol. Chem. 1992, 267, 1640–1647. [Google Scholar] [PubMed]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed]
- Gupte, M.; Kulkarni, P.; Ganguli, B. Antifungal antibiotics. Appl. Microbiol. Biotechnol. 2002, 58, 46–57. [Google Scholar] [CrossRef]
- Schneiter, R.; Di Pietro, A. The CAP protein superfamily: Function in sterol export and fungal virulence. Biomol. Concepts 2013, 4. [Google Scholar] [CrossRef]
- Darwiche, R.; El Atab, O.; Cottier, S.; Schneiter, R. The function of yeast CAP family proteins in lipid export, mating, and pathogen defense. FEBS Lett. 2018, 592, 1304–1311. [Google Scholar] [CrossRef]
- Suzuki, N.; Yamazaki, Y.; Brown, R.L.; Fujimoto, Z.; Morita, T.; Mizuno, H. Structures of pseudechetoxin and pseudecin, two snake-Venom cysteine-Rich secretory proteins that target cyclic nucleotide-Gated ion channels: Implications for movement of the C-terminal cysteine-Rich domain. Acta Crystallogr. Sect. D Biol. Crystallogr. 2008, 64, 1034–1042. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Kuo, J.-H.; Lee, S.-C.; Liu, J.-S.; Hsieh, Y.-C.; Shih, Y.-T.; Chen, C.-J.; Chiu, J.-J.; Wu, W.-G. Cobra CRISP Functions as an Inflammatory Modulator via a Novel Zn2+- and Heparan Sulfate-Dependent Transcriptional Regulation of Endothelial Cell Adhesion Molecules. J. Biol. Chem. 2010, 285, 37872–37883. [Google Scholar] [CrossRef]
- Xiao, R.; Li, Q.-W.; Perrett, S.; He, R.-Q. Characterisation of the fibrinogenolytic properties of the buccal gland secretion from Lampetra japonica. Biochimie 2007, 89, 383–392. [Google Scholar] [CrossRef]
- Jiang, Q.; Liu, Y.; Duan, D.; Gou, M.; Wang, H.; Wang, J.; Li, Q.; Xiao, R. Anti-Angiogenic activities of CRBGP from buccal glands of lampreys (Lampetra japonica). Biochimie 2016, 123, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.; Xiao, R.; Li, Q.; Zhou, L.; He, R.; Qi, Z. Suppression of neuronal excitability by the secretion of the lamprey (Lampetra japonica) provides a mechanism for its evolutionary stability. Pflügers Arch.-Eur. J. Physiol. 2009, 458, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Liu, Y.; Gou, M.; Han, J.; Wang, J.; Li, Q.; Xiao, R. Data for the inhibition effects of recombinant lamprey CRBGP on the tube formation of HUVECs and new blood vessel generation in CAM models. Data Brief 2016, 6, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Bai, J.; Sun, J.; Wu, Y.; Yu, S.Y.; Guo, R.Y.; Liu, X.; Li, Q.W. Novel neutrophil inhibitory factor homologue in the buccal gland secretion of Lampetra japonica. Biol. Chem. 2011, 392. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Pang, Y.; Li, Q.W. The buccal gland of Lampetra japonica is a source of diverse bioactive proteins. Biochimie 2012, 94, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Nobile, M.; Noceti, F.; Prestipino, G.; Possani, L. Helothermine, a lizard venom toxin, inhibits calcium current in cerebellar granules. Exp. Brain Res. 1996, 110. [Google Scholar] [CrossRef]
- Nobile, M.; Magnelli, V.; Lagostena, L.; Mochca-Morales, J.; Possani, L.D.; Prestipino, G. The toxin helothermine affects potassium currents in newborn rat cerebellar granule cells. J. Membr. Biol. 1994, 139. [Google Scholar] [CrossRef]
- Mochca-Morales, J.; Martin, B.M.; Possani, L.D. Isolation and characterization of Helothermine, a novel toxin from Heloderma horridum horridum (Mexican beaded lizard) venom. Toxicon 1990, 28, 299–309. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Brown, R.L.; Morita, T. Purification and Cloning of Toxins from Elapid Venoms that Target Cyclic Nucleotide-Gated Ion Channels. Biochemistry 2002, 41, 11331–11337. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Guo, M.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; Hao, Q. Crystal Structure of the Cysteine-Rich Secretory Protein Stecrisp Reveals That the Cysteine-Rich Domain Has a K + Channel Inhibitor-Like Fold. J. Biol. Chem. 2005, 280, 12405–12412. [Google Scholar] [CrossRef] [PubMed]
- Lodovicho, M.E.; Costa, T.R.; Bernardes, C.P.; Menaldo, D.L.; Zoccal, K.F.; Carone, S.E.; Rosa, J.C.; Pucca, M.B.; Cerni, F.A.; Arantes, E.C.; et al. Investigating possible biological targets of Bj-CRP, the first cysteine-Rich secretory protein (CRISP) isolated from Bothrops jararaca snake venom. Toxicol. Lett. 2017, 265, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Suntravat, M.; Cromer, W.E.; Marquez, J.; Galan, J.A.; Zawieja, D.C.; Davies, P.; Salazar, E.; Sánchez, E.E. The isolation and characterization of a new snake venom cysteine-Rich secretory protein (svCRiSP) from the venom of the Southern Pacific rattlesnake and its effect on vascular permeability. Toxicon 2019, 165, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Lecht, S.; Chiaverelli, R.A.; Gerstenhaber, J.; Calvete, J.J.; Lazarovici, P.; Casewell, N.R.; Harrison, R.; Lelkes, P.I.; Marcinkiewicz, C. Anti-Angiogenic activities of snake venom CRISP isolated from Echis carinatus sochureki. Biochim. Biophys. Acta BBA Gen. Subj. 2015, 1850, 1169–1179. [Google Scholar] [CrossRef]
- Adade, C.M.; Carvalho, A.L.O.; Tomaz, M.A.; Costa, T.F.R.; Godinho, J.L.; Melo, P.A.; Lima, A.P.C.A.; Rodrigues, J.C.F.; Zingali, R.B.; Souto-Padrón, T. Crovirin, a Snake Venom Cysteine-Rich Secretory Protein (CRISP) with Promising Activity against Trypanosomes and Leishmania. PLoS Negl. Trop. Dis. 2014, 8, e3252. [Google Scholar] [CrossRef]
- Brown, R.L.; Haley, T.L.; West, K.A.; Crabb, J.W. Pseudechetoxin: A peptide blocker of cyclic nucleotide-Gated ion channels. Proc. Natl. Acad. Sci. USA 1999, 96, 754–759. [Google Scholar] [CrossRef]
- Wang, J.; Shen, B.; Guo, M.; Lou, X.; Duan, Y.; Cheng, X.P.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; et al. Blocking Effect and Crystal Structure of Natrin Toxin, a Cysteine-Rich Secretory Protein from Naja atra Venom that Targets the BK Ca Channel. Biochemistry 2005, 44, 10145–10152. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, Q.-L.; Meng, X.; Shu, Y.; Jiang, T.; Wagenknecht, T.; Yin, C.-C.; Sui, S.-F.; Liu, Z. Structural and Functional Characterization of Ryanodine Receptor-Natrin Toxin Interaction. Biophys. J. 2008, 95, 4289–4299. [Google Scholar] [CrossRef]
- Wang, F.; Li, H.; Liu, M.-N.; Song, H.; Han, H.-M.; Wang, Q.-L.; Yin, C.-C.; Zhou, Y.-C.; Qi, Z.; Shu, Y.-Y.; et al. Structural and functional analysis of natrin, a venom protein that targets various ion channels. Biochem. Biophys. Res. Commun. 2006, 351, 443–448. [Google Scholar] [CrossRef]
- Peichoto, M.E.; Mackessy, S.P.; Teibler, P.; Tavares, F.L.; Burckhardt, P.L.; Breno, M.C.; Acosta, O.; Santoro, M.L. Purification and characterization of a cysteine-Rich secretory protein from Philodryas patagoniensis snake venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 79–84. [Google Scholar] [CrossRef]
- Magdaleno, L.; Gasset, M.A.; Varea, J.; Schambony, A.M.; Urbanke, C.; Raida, M.; Töpfer-Petersen, E.; Calvete, J.J. Biochemical and conformational characterisation of HSP-3, a stallion seminal plasma protein of the cysteine-Rich secretory protein (CRISP) family. FEBS Lett. 1997, 420, 179–185. [Google Scholar] [CrossRef]
- Kosari, F.; Asmann, Y.W.; Cheville, J.C.; Vasmatzis, G. Cysteine-Rich secretory protein-3: A potential biomarker for prostate cancer. Cancer Epidemiol. Prev. Biomark. 2002, 11, 1419–1426. [Google Scholar]
- Udby, L.; Lundwall, Å.; Johnsen, A.H.; Fernlund, P.; Valtonen-André, C.; Blom, A.M.; Lilja, H.; Borregaard, N.; Kjeldsen, L.; Bjartell, A. β-Microseminoprotein binds CRISP-3 in human seminal plasma. Biochem. Biophys. Res. Commun. 2005, 333, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Udby, L.; Sørensen, O.E.; Pass, J.; Johnsen, A.H.; Behrendt, N.; Borregaard, N.; Kjeldsen, L. Cysteine-Rich Secretory Protein 3 Is a Ligand of α1B-Glycoprotein in Human Plasma †. Biochemistry 2004, 43, 12877–12886. [Google Scholar] [CrossRef]
- Reeves, J.R.; Xuan, J.W.; Arfanis, K.; Morin, C.; Garde, S.V.; Ruiz, M.T.; Wisniewski, J.; Panchal, C.; Tanner, J.E. Identification, purification and characterization of a novel human blood protein with binding affinity for prostate secretory protein of 94 amino acids. Biochem. J. 2005, 385, 105–114. [Google Scholar] [CrossRef]
- Anklesaria, J.H.; Jagtap, D.D.; Pathak, B.R.; Kadam, K.M.; Joseph, S.; Mahale, S.D. Prostate Secretory Protein of 94 Amino Acids (PSP94) Binds to Prostatic Acid Phosphatase (PAP) in Human Seminal Plasma. PLoS ONE 2013, 8, e58631. [Google Scholar] [CrossRef]
- Ghasriani, H.; Fernlund, P.; Udby, L.; Drakenberg, T. A model of the complex between human β-Microseminoprotein and CRISP-3 based on NMR data. Biochem. Biophys. Res. Commun. 2009, 378, 235–239. [Google Scholar] [CrossRef]
- Breed, A.A.; Gomes, A.; Roy, B.S.; Mahale, S.D.; Pathak, B.R. Mapping of the binding sites involved in PSP94–CRISP-3 interaction by molecular dissection of the complex. Biochim. Biophys. Acta BBA Gen. Subj. 2013, 1830, 3019–3029. [Google Scholar] [CrossRef]
- Shioi, N.; Tadokoro, T.; Shioi, S.; Okabe, Y.; Matsubara, H.; Kita, S.; Ose, T.; Kuroki, K.; Terada, S.; Maenaka, K. Crystal structure of the complex between venom toxin and serum inhibitor from Viperidae snake. J. Biol. Chem. 2019, 294, 1250–1256. [Google Scholar] [CrossRef]
- Aoki, N.; Sakiyama, A.; Deshimaru, M.; Terada, S. Identification of novel serum proteins in a Japanese viper: Homologs of mammalian PSP94. Biochem. Biophys. Res. Commun. 2007, 359, 330–334. [Google Scholar] [CrossRef]
- Hansson, K.; Kjellberg, M.; Fernlund, P. Cysteine-Rich secretory proteins in snake venoms form high affinity complexes with human and porcine β-Microseminoproteins. Toxicon 2009, 54, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, Y.; Yamazaki, Y.; Hyodo, F.; Sugiyama, Y.; Nozaki, M.; Morita, T. Structural Divergence of Cysteine-Rich Secretory Proteins in Snake Venoms. J. Biochem. 2009, 145, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lu, Q.; Zhou, X.; Zhu, S.; Li, R.; Wang, W.; Xiong, Y. Purification and cloning of cysteine-Rich proteins from Trimeresurus jerdonii and Naja atra venoms. Toxicon 2003, 42, 539–547. [Google Scholar] [CrossRef]
- Aoki, N.; Sakiyama, A.; Kuroki, K.; Maenaka, K.; Kohda, D.; Deshimaru, M.; Terada, S. Serotriflin, a CRISP family protein with binding affinity for small serum protein-2 in snake serum. Biochim. Biophys. Acta BBA Proteins Proteom. 2008, 1784, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Del Tordello, E.; Vacca, I.; Ram, S.; Rappuoli, R.; Serruto, D. Neisseria meningitidis NalP cleaves human complement C3, facilitating degradation of C3b and survival in human serum. Proc. Natl. Acad. Sci. USA 2014, 111, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Evolution of CRISPs Associated with Toxicoferan-Reptilian Venom and Mammalian Reproduction. Mol. Biol. Evol. 2012, 29, 1807–1822. [Google Scholar] [CrossRef] [PubMed]
- Manceau, M.; Marin, J.; Morlon, H.; Lambert, A. Model-Based Inference of Punctuated Molecular Evolution. BioRxiv 2019. [Google Scholar] [CrossRef]
- Perry, B.W.; Card, D.C.; McGlothlin, J.W.; Pasquesi, G.I.M.; Adams, R.H.; Schield, D.R.; Hales, N.R.; Corbin, A.B.; Demuth, J.P.; Hoffmann, F.G.; et al. Molecular Adaptations for Sensing and Securing Prey and Insight into Amniote Genome Diversity from the Garter Snake Genome. Genome Biol. Evol. 2018, 10, 2110–2129. [Google Scholar] [CrossRef]
- Suryamohan, K.; Krishnankutty, S.P.; Guillory, J.; Jevit, M.; Schröder, M.S.; Wu, M.; Kuriakose, B.; Mathew, O.K.; Perumal, R.C.; Koludarov, I.; et al. The Indian cobra reference genome and transcriptome enables comprehensive identification of venom toxins. Nat. Genet. 2020, 52, 106–117. [Google Scholar] [CrossRef]
- Schield, D.R.; Card, D.C.; Hales, N.R.; Perry, B.W.; Pasquesi, G.M.; Blackmon, H.; Adams, R.H.; Corbin, A.B.; Smith, C.F.; Ramesh, B.; et al. The origins and evolution of chromosomes, dosage compensation, and mechanisms underlying venom regulation in snakes. Genome Res. 2019, 29, 590–601. [Google Scholar] [CrossRef]
- Shibata, H.; Chijiwa, T.; Oda-Ueda, N.; Nakamura, H.; Yamaguchi, K.; Hattori, S.; Matsubara, K.; Matsuda, Y.; Yamashita, A.; Isomoto, A.; et al. The habu genome reveals accelerated evolution of venom protein genes. Sci. Rep. 2018, 8, 11300. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Animals | Name | Species | Target (Interaction Molecules) | Biological Effect (or Related Investigation) | Accession Numbers | Ref. |
|---|---|---|---|---|---|---|
| Lamprey | Lamprey CRISP Buccal gland secretion protein-2 (BGSP-2) Cysteine-rich buccal gland protein (CRBGP) | Lethenteron japonicum (Lampetra japonica) | ・Voltage-dependent Na+ channels ・Integrin β2 (CD11s/CD18) | Ca2+ channel blocker-like properties Anti-angiogenic activities Permeability Inhibition of adhesion, proliferation, migration, and invasion of cells (HUVEC; IC50 = 4.0 μM and Hela cell; IC50 = 6.7 μM) Non-fibrinogenolytic activity Activity of immunosuppressant (neutrophil inhibitory factor) Inhibition of Na+ channels in hippocampal neurons (12 μM) Inhibition K+ channels in hippocampal neurons (120 μM) | A4PIZ5 | [35,51,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56] |
| Cone snail | Tex31 | Conus textile | N.D. | Proteinase | Q7YT83 | [32] |
| Lizard | Helothermine | Heloderma horridum salivary secretion | ・Ryanodine receptors ・Ca2+ channels ・K+ channels | Lethargy, partial paralysis of rear limbs and lowering of body temperature Blockage of receptors (Cerebellar Granule Cells) Inhibition of K+ channels (IC50 = 0.52 μM) Inhibition of Ca2+ channels (IC50 = 0.25 μM) Inhibition of skeletal ryanodine receptors (about 1.0 μM ) | Q91055 | [40,57,58,59] |
| Snake | ||||||
| (Vipers) | Ablomin | Gloydius blomhoffi. | N.D. | Ca2+ channel blocker-like properties | Q8JI40 | [60] |
| Piscivorinc | Agkistrodon piscivorus | N.D. | Ca2+ channel blocker-like properties | AY181982 | [61] | |
| Catrin | Crotalus atrox | N.D. | Ca2+ channel blocker-like properties | AY181983 | [61] | |
| Triflin | Protobothrops flavoviridis | N.D. | Ca2+ channel blocker-like properties | Q8JI39 1WVR, 6IMF (with inhibitor) | [60] | |
| Stecrisp | Trimeresurus stejnegeri (Viridovipera stejnegeri) | N.D. | No proteolysis activity (unlikeTex31) | P60623 1RC9 | [62] | |
| Bj-CRP | Bothrops jararaca | Component C3 and C4 | Bind and cleaved to component C3 and C4 Lack of effect of K+ channel blockage activity (1.0 μM) (Kv1.1 to Kv1.6, Shaker-IR, Kv3.1, Kv7.1, Kv7.2, Kv7.4 and Kv10.1) | N.D. (partial sequence) | [63] | |
| Hellerin | Crotalus oreganus helleri | N.D. | To increase trans-epithelial permeability Cytotoxicity against HUVEC (cytotoxic concentration CC50 = 2.3 μM) | G9DCH4 | [64] | |
| EC-CRISP | Echis Carinatus Sochureki | N.D. | Binding to HUVEC cell Anti-angiogenic Activities (10-20 μg/ml, <1 μM) | P0DMT4 | [65] | |
| Crovirin | Crotalus viridis viridis | N.D. | Anti-protozoan activity against Trypanosoma cruzi and Leishmania amazonensis | N.D (partial sequence) | [66] | |
| (Elapid) | Pseudechetoxin (PsTx) | Pseudechis australis | Cyclic nucleotide- gated ion channels | Inhibition of CNGA1 subunit (apparent Ki = 70 nM) Inhibition of CNGA2 subunits (apparent Ki = 15 nM) | Q8AVA4 2DDA | [49,67] |
| Pseudecin | Pseudechis porphyriacus | Cyclic nucleotide-gated ion channels | Ca2+ channel blocker-like properties | Q8AVA3 2DDB, 2EPF(Zn2+) | [49,60] | |
| Latisemin | Laticauda semifasciata | N.D. | Ca2+ channel blocker-like properties | Q8JI38 | [60] | |
| Ophanin | Ophiophagus hannah | N.D. | Ca2+ channel blocker-like properties | AY181984 | [61] | |
| Natrin | Naja Naja atra | Bkca Kv1.3 Ryanodine receptors Heparin | High-conductance calcium-activated potassium (BKCa) channel (34.4 nM) Inhibition of Kv1.3 (10–200 nM) Inhibition of ryanodine receptors (1 μM, Kd = 1.5–2.3 nM) Inflammatory Modulator (<1.0 μM) Non-proteolytic activity (BSA, neurotensin, Tex31 substrate, kenetensin) | Q7T1K6 1XX5, 1XTA, 2GIZ 3MZ8 (Zn2+) | [68,69,70] | |
| (Colubrid) | Tigrin | Rhabdophis tigrinus | N.D. | Non-Ca2+ channel blocker-like properties | Q8JGT9 | [60] |
| Patagonin | Philodryas patagoniensis | N.D. | Non-Ca2+ channel blocker-like properties Non-fibrinogenolytic activity Skeletal myotoxic activity (43 and 87 μM) | N.D. (only N-terminal sequence) | [71] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadokoro, T.; M. Modahl, C.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins 2020, 12, 175. https://doi.org/10.3390/toxins12030175
Tadokoro T, M. Modahl C, Maenaka K, Aoki-Shioi N. Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins. 2020; 12(3):175. https://doi.org/10.3390/toxins12030175
Chicago/Turabian StyleTadokoro, Takashi, Cassandra M. Modahl, Katsumi Maenaka, and Narumi Aoki-Shioi. 2020. "Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily" Toxins 12, no. 3: 175. https://doi.org/10.3390/toxins12030175
APA StyleTadokoro, T., M. Modahl, C., Maenaka, K., & Aoki-Shioi, N. (2020). Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins, 12(3), 175. https://doi.org/10.3390/toxins12030175