Isolation, Molecular Identification, and Mycotoxin Production of Aspergillus Species Isolated from the Rhizosphere of Sugarcane in the South of Iran

 ,
,  ,
, 

Abstract
1. Introduction
2. Results
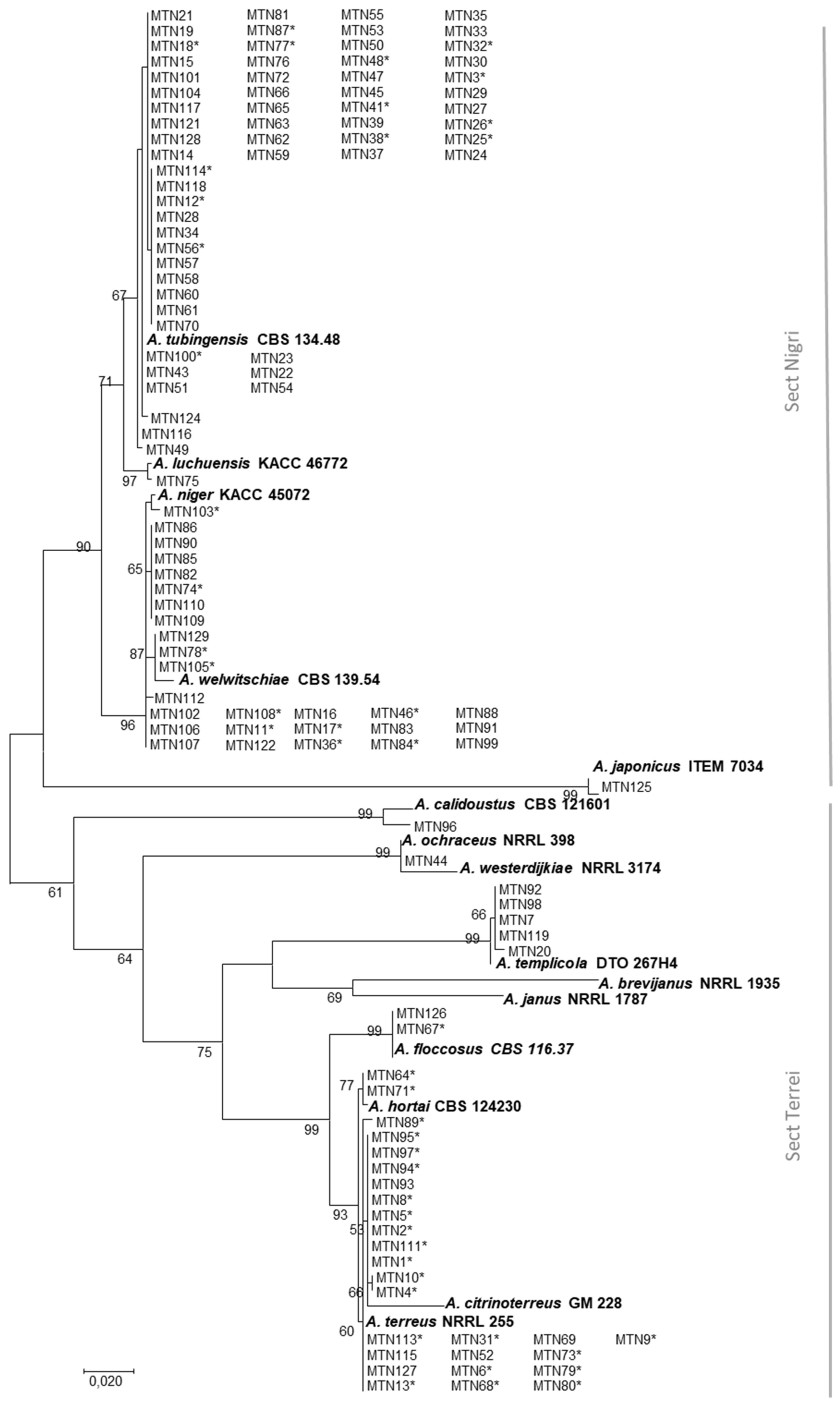
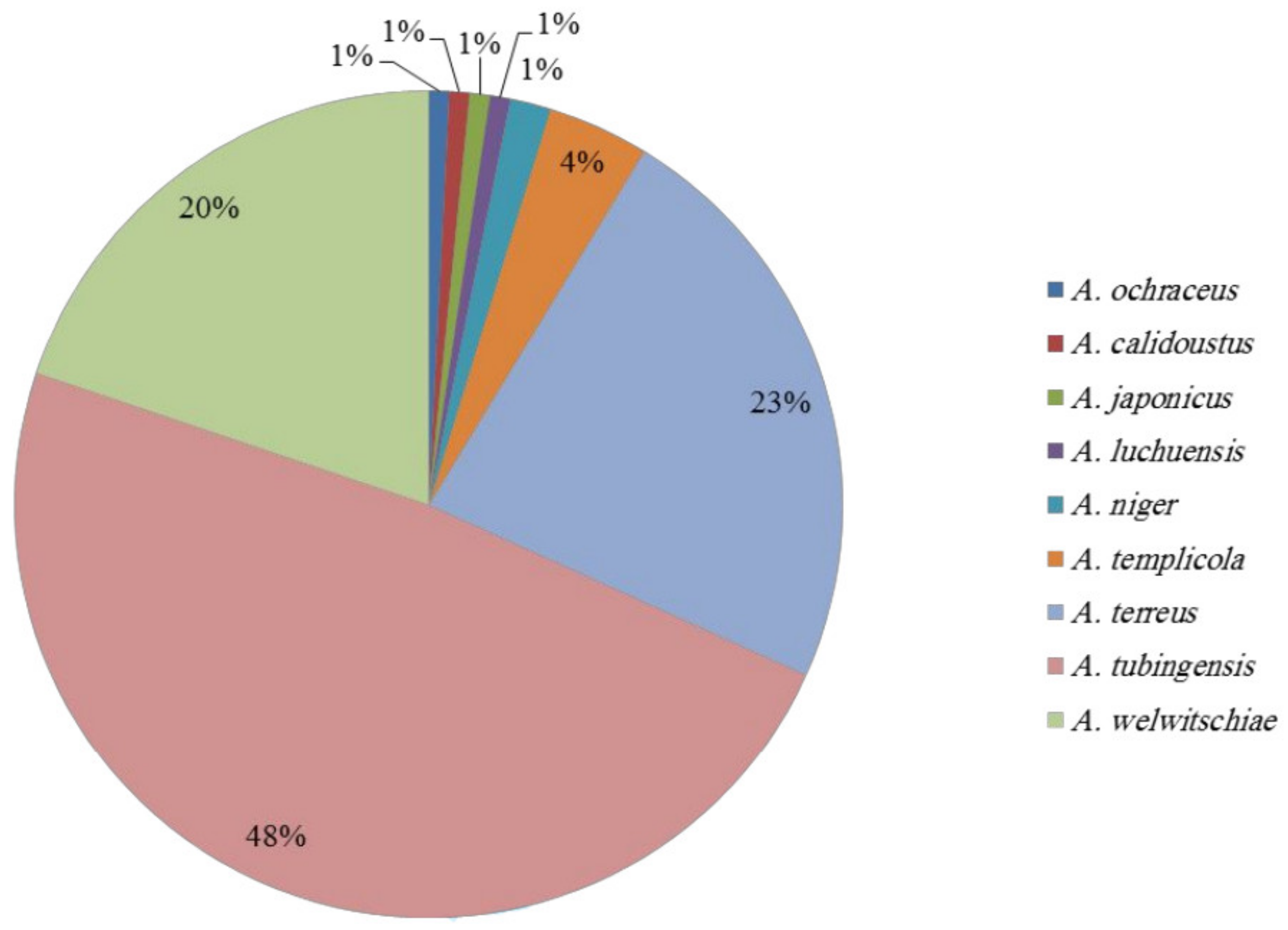
2.1. Species Identification
2.2. Mycotoxin Production
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples
5.2. Fungal Isolation and Morphological Identification
5.3. Molecular Identification
5.4. Sequence Data Analysis
5.5. Mycotoxin Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization of the United Nations). FAOSTAT. Available online: http://faostat.fao.org/faostat/ (accessed on 21 November 2010).
- Lozano, F.J.; Lozano, R. Assessing the potential sustainability benefits of agricultural residues: Biomass conversion to syngas for energy generation or to chemicals production. J. Clean. Prod. 2018, 172, 4162–4169. [Google Scholar] [CrossRef]
- Kumar, D.; Jain, V.K.; Shanker, G.; Srivastava, A. Citric acid production by solid state fermentation using sugarcane bagasse. Process Biochem. 2003, 38, 1731–1738. [Google Scholar] [CrossRef]
- Varga, J.; Kevei, F.; Hamari, Z.; Toth, B.; Teren, J.; Croft, H.; Kozakiewicz, Z. Genotypic and phenotypic variability among black aspergilli. In Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 397–411. [Google Scholar]
- Abarca, M.L.; Accensi, F.; Cano, J.; Cabañes, F.J. Taxonomy and significance of black aspergilli. Antonie Van Leeuwenhoek 2004, 86, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Oliván, A.; Marín, S.; Sancis, V.; Ramos, A.J. Effect of intra and interspecific interaction on OTA production by A. section Nigri in grapes during dehydration. Food Microbiol. 2007, 24, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Chávez, R.; Bull, P.; Eyzaguirre, J. The xylanolytic enzyme system from the genus Penicillium. J. Biotechnol. 2006, 123, 413–433. [Google Scholar]
- Ichishima, E. Development of enzyme technology for Aspergillus oryzae, A. sojae, and A. luchuensis, the national microorganisms of Japan. Biosci. Biotechnol. Biochem. 2016, 80, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.Y.; Demey, J.R.; Fuchs, M.; González, V.; Rea, R.; De Sousa, O.; Gutiérrez, Z. Selection of sugarcane plants resistant to SCMV. Plant. Sci. 2003, 165, 221–225. [Google Scholar] [CrossRef]
- Abdallah, M.; Krska, R.; Sulyok, M. Mycotoxin contamination in sugarcane grass and juice: First report on detection of multiple mycotoxins and exposure assessment for aflatoxins B1 and G1 in humans. Toxins 2016, 8, 343. [Google Scholar] [CrossRef]
- Graham, M.H.; Haynes, R.J. Organic matter status and the size, activity and metabolic diversity of the soil microbial community in the row and inter-row of sugarcane under burning and trash retention. Soil Biol. Biochem. 2006, 38, 21–31. [Google Scholar] [CrossRef]
- De Souza, R.S.; Okura, V.K.; Armanhi, J.S.; Jorrín, B.; Lozano, N.; da Silva, M.J.; González-Guerrero, M.; de Araújo, L.M.; Verza, N.C.; Bagheri, H.C.; et al. Unlocking the bacterial, and fungal communities assemblages of sugarcane microbiome. Sci. Rep. 2016, 6, 28774. [Google Scholar] [CrossRef] [PubMed]
- Kamei, K.; Watanabe, A. Aspergillus mycotoxins and their effect on the host. Med. Mycol. 2005, 43, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Herbrecht, R.; Letscher-Bru, V.; Oprea, C.; Lioure, B.; Waller, J.; Campos, F.; Villard, O.; Liu, K.-L.; Natarajan-Amé, S.; Lutz, P.; et al. Aspergillus galactomannan detection in the diagnosis of invasive aspergillosis in cancer patients. J. Clin. Oncol. 2002, 20, 1898–1906. [Google Scholar] [CrossRef] [PubMed]
- Schuster, E.; Dunn-Coleman, N.; Frisvad, J.C.; Van Dijck, P.W. On the safety of Aspergillus niger—A review. Appl. Microbiol. Biotechnol. 2002, 59, 426–435. [Google Scholar]
- Daiani, M.S.; Batista, L.R.; Rezende, E.F.; Fungaro, M.H.P.; Sartori, D.; Alves, E. Identification of fungi of the genus Aspergillus section Nigri using polyphasic taxonomy. Braz. J. Microbiol. 2011, 42, 761–773. [Google Scholar]
- Nielsen, K.F.; Mogensen, J.M.; Johansen, M.; Larsen, T.O.; Frisvad, J.C. Review of secondary metabolites and mycotoxins from the Aspergillus niger group. Annal. Bioanal. Chem. 2009, 395, 1225–1242. [Google Scholar] [CrossRef]
- Jahromi, M.F.; Liang, J.B.; Ho, Y.W.; Mohamad, R.; Goh, Y.M.; Shokryazdan, P. Lovastatin production by Aspergillus terreus using agro-biomass as substrate in solid state fermentation. J. Biomed. Biotechnol. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Bizukojc, M.; Pawlak, M.; Boruta, T.; Gonciarz, J. Effect of pH on biosynthesis of lovastatin and other secondary metabolites by Aspergillus terreus ATCC 20542. J. Biotechnol. 2012, 162, 253–261. [Google Scholar] [CrossRef]
- Samson, R.A.; Peterson, S.W.; Frisvad, J.C.; Varga, J. New species in Aspergillus section Terrei. Stud. Mycol. 2011, 69, 39–55. [Google Scholar] [CrossRef]
- Luque, M.I.; Rodríguez, A.; Andrade, M.J.; Gordillo, R.; Rodríguez, M.; Córdoba, J.J. Development of a PCR protocol to detect patulin producing moulds in food products. Food Control 2011, 22, 831–1838. [Google Scholar] [CrossRef]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. Food Saf. 2005, 4, 8–21. [Google Scholar] [CrossRef]
- Baumgardner, D.J. Soil-related bacterial and fungal infections. J. Am. Board Fam. Med. 2012, 25, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Eberl, L.; Hartmann, A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 2005, 7, 1673–1685. [Google Scholar] [CrossRef]
- Leger, R.J.S.; Screen, S.E.; Shams-Pirzadeh, B. Lack of host specialization in Aspergillus flavus. Appl. Environ. Microbiol. 2000, 66, 320–324. [Google Scholar] [CrossRef]
- Klich, M.A. Identification of Common Aspergillus Species; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002. [Google Scholar]
- Hariprasad, P.; Vipin, A.V.; Karuna, S.; Raksha, R.K.; Venkateswaran, G. Natural aflatoxin uptake by sugarcane (Saccharum officinaurum L.) and its persistence in jaggery. Environ. Sci. Pollut. Res. 2015, 22, 6246–6253. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; Baker, J.L.; Mahoney, N.E. Isolation of bacterial antagonists of Aspergillus flavus from almonds. Microb. Ecol. 2006, 52, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Djossou, O.; Perraud-Gaime, I.; Mirleau, F.L.; Rodriguez-Serrano, G.; Karou, G.; Niamke, S.; Ouzari, I.; Boudabous, A.; Roussos, S. Robusta coffee beans post-harvest microflora: Lactobacillus plantarum sp. as potential antagonist of Aspergillus carbonarius. Anaerobe 2011, 17, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H.P.; Vakharia, D.N. Molecular and biochemical characterization of Trichoderma isolates inhibiting a phytopathogenic fungi Aspergillus niger Van Tieghem. Physiol. Mol. Plant. Pathol. 2010, 74, 274–282. [Google Scholar] [CrossRef]
- Hill, R.A.; Blankenship, P.D.; Cole, R.J.; Sanders, T.H. Effects of soil moisture and temperature on preharvest invasion of peanuts by the Aspergillus flavus group and subsequent aflatoxin development. Appl. Environ. Microbiol. 1983, 45, 628–633. [Google Scholar] [CrossRef]
- Susca, A.; Proctor, R.H.; Morelli, M.; Haidukowski, M.; Gallo, A.; Logrieco, A.F.; Moretti, A. Variation in fumonisin and ochratoxin production associated with differences in biosynthetic gene content in Aspergillus niger and A. welwitschiae isolates from multiple crop and geographic origins. Front. Microbiol. 2016, 7, 1412. [Google Scholar] [CrossRef]
- Medina, A.; Mateo, R.; López-Ocana, L.; Valle-Algarra, F.M.; Jiménez, M. Study of Spanish grape mycobiota and ochratoxin A production by isolates of Aspergillus tubingensis and other members of Aspergillus section Nigri. Appl. Environ. Microbiol. 2005, 71, 4696–4702. [Google Scholar] [CrossRef] [PubMed]
- Susca, A.; Proctor, R.H.; Butchko, R.A.; Haidukowski, M.; Stea, G.; Logrieco, A.F.; Moretti, A. Variation in the fumonisin biosynthetic gene cluster in fumonisin-producing and nonproducing black aspergilli. Fungal Genet. Biol. 2014, 73, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Fiorentino, C.; Spataro, F.; Nostro, A. Effect of temperature on production of ochratoxin A by Aspergillus niger in orange juice. J. Toxins 2014, 2014, 1–5. [Google Scholar] [CrossRef]
- Abarca, M.L.; Bragulat, M.R.; Castella, G.; Cabañes, F.J. Ochratoxin A production by strains of Aspergillus niger var. niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar] [CrossRef] [PubMed]
- Singleton, L.L.; Mihailm, J.D.; Rush, C.M. Methods for research on soilborne phytopathogenic fungi. In The American Phytopathological Society; APS Press: St Paul, MI, USA, 1993; Volume 85, pp. 140–141. [Google Scholar]
- Bills, G.F.; Christensen, M.; Powell, M.; Thorn, G. Saprobic soil fungi. In Biodiversity of Fungi: Inventory and Monitoring Methods; Muller, G.M., Bills, G.F., Foster, M.S., Eds.; Elsevier Academic Press: Burlington, MA, USA, 2004; pp. 271–302. [Google Scholar]
- Johnson, L.F.; Curl, E.A.; Bond, J.H.; Fribourg, H.A. Methods for Studying Soil Mycoflora: Plant Disease Relationships; Burgess Publishing: Minneapolis, CO, USA, 1960; p. 179. [Google Scholar]
- Warcup, I.M. Method for Isolation and Estimation of Activity of Fungi in Soil. In The Ecology of Soil. An International Symposium; Liverpool University Press: Liverpool, UK, 1960; pp. 3–21. [Google Scholar]
- Gams, W.; Howekstra, E.S.; Aptroot, A. CBS Course of Mycology; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 1998; pp. 1–165. [Google Scholar]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; Williams & Wilkins: Baltimore, MD, USA, 1965; p. 114. [Google Scholar]
- Pitt, J.; Hocking, A. The ecology of Fungal Food Spoilage. In Fungi and Food Spoilage; Blackie Academic and Professional: London, UK, 2009; pp. 3–9. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS KNAW Biodiversity Center: Utrecht, The Netherlands, 2010. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A Multigene phylogeny of the Gibberellafujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC) Official Method 2008-08. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&cPath=1&products_id=2816 (accessed on 16 August 2016).
- CEN (European Committee for Standardization). Foodstuffs-Determination of patulin in fruit juice and fruit based puree for young children—HPLC method with liquid/liquid partition clean-up and solid phase extraction and UV determination. CEN/TC 275/WG5. In Food analysis–Biotoxins–Foodstuffs–EN 15890; CEN: Brussels, Belgium, 2002. [Google Scholar]
- Sewram, V.; Nair, J.J.; Nieuwoudt, T.W.; Leggott, N.L.; Shephard, G.S. Determination of patulin in apple juice by high-performance liquid chromatography–atmospheric pressure chemical ionization mass spectrometry. J. Chromat A 2000, 897, 365–374. [Google Scholar] [CrossRef]
- Veršilovskis, A.; De Saeger, S.; Mikelsone, V. Determination of sterigmatocystin in beer by high performance liquid chromatography with ultraviolet detection. W. Mycotox. J. 2008, 1, 161–166. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | No. of Sugarcane Rhizosphere Samples | No. of Aspergillus Isolates | Aspergillus spp. Occurrence (%) | |||
|---|---|---|---|---|---|---|
| A. terreus | A. tubingensis | A. welwitschiae | Others | |||
| Amir Kabir | 8 | 8 | - - | 75 | 12.5 | 12.5 |
| Imam Khomeini | 8 | 4 | 25 | 50 | 25 | - - |
| Debal Khazaei | 8 | 11 | - - | 73 | 18 | 9 |
| Dehkhoda | 8 | 10 | 80 | 10 | - - | 10 |
| Karun | 8 | 23 | 25 | 29 | 42 | 4 |
| Mirza K. Khan | 8 | 11 | 9 | 82 | 9 | - - |
| Salman Farsi | 8 | 26 | 19 | 81 | - - | - - |
| Haft Tappeh | 8 | 32 | 21 | 24 | 30 | 24 |
| Total | 64 | 125 | 23 | 48 | 20 | 9 |
| ITEM Collection Acc. Number | IRAN Collection Acc. Number | Isolate ID | Species | Mycotoxin Production (μg/g) | ||
|---|---|---|---|---|---|---|
| FB2 | OTA | PAT * | ||||
| 18460 | 3629 | MTN11 | A. welwitschiae | 1.0 | 106 | n.t. |
| 18461 | 3533 | MTN17 | A. welwitschiae | 1.0 | 32 | n.t. |
| 18462 | 3550 | MTN36 | A. welwitschiae | <LOD | <LOD | n.t. |
| 18464 | 3632 | MTN46 | A. welwitschiae | 1.0 | <LOD | n.t. |
| 18465 | 3583 | MTN74 | A. welwitschiae | <LOD | <LOD | n.t. |
| 18466 | 3587 | MTN78 | A. welwitschiae | 0.1 | <LOD | n.t. |
| 18467 | 3606 | MTN100 | A. tubingensis | n.t. | <LOD | n.t. |
| 18468 | 3608 | MTN103 | A. welwitschiae | 4.4 | <LOD | n.t. |
| 18469 | 3609 | MTN105 | A. welwitschiae | 0.2 | 49 | n.t. |
| 18470 | 3612 | MTN108 | A. welwitschiae | 2.2 | 142 | n.t. |
| 18471 | 3521 | MTN3 | A. tubingensis | n.t. | <LOD | n.t. |
| 18472 | 3528 | MTN12 | A. tubingensis | n.t. | <LOD | n.t. |
| 18473 | 3530 | MTN14 | A. tubingensis | n.t. | <LOD | n.t. |
| 18474 | 3534 | MTN18 | A. tubingensis | n.t. | <LOD | n.t. |
| 18475 | 3541 | MTN25 | A. tubingensis | n.t. | <LOD | n.t. |
| 18476 | 3542 | MTN26 | A. tubingensis | n.t. | <LOD | n.t. |
| 18477 | 3547 | MTN32 | A. tubingensis | n.t. | <LOD | n.t. |
| 18478 | 3552 | MTN38 | A. tubingensis | n.t. | <LOD | n.t. |
| 18479 | 3554 | MTN41 | A. tubingensis | n.t. | <LOD | n.t. |
| 18480 | 3559 | MTN48 | A. tubingensis | n.t. | <LOD | n.t. |
| 18481 | 3567 | MTN56 | A. tubingensis | n.t. | <LOD | n.t. |
| 18482 | 3586 | MTN77 | A. tubingensis | n.t. | <LOD | n.t. |
| 18483 | 3595 | MTN87 | A. tubingensis | <LOD | <LOD | <LOD |
| 18484 | 3616 | MTN114 | A. tubingensis | <LOD | <LOD | <LOD |
| 18485 | 3520 | MTN1 | A. terreus | n.t. | n.t. | 523.56 |
| 18486 | 3627 | MTN2 | A. terreus | n.t. | n.t. | 18.39 |
| 18487 | 3628 | MTN4 | A. terreus | n.t. | n.t. | n.t. |
| 18488 | 3522 | MTN5 | A. terreus | n.t. | n.t. | n.t. |
| 18489 | 3523 | MTN6 | A. terreus | n.t. | n.t. | 172.39 |
| 18490 | 3525 | MTN8 | A. terreus | n.t. | n.t. | 212.65 |
| 18491 | 3526 | MTN9 | A. terreus | n.t. | n.t. | n.t. |
| 18492 | 3527 | MTN10 | A. terreus | n.t.. | n.t. | n.t. |
| 18493 | 3529 | MTN13 | A. terreus | n.t. | n.t. | n.t. |
| 18494 | 3546 | MTN31 | A. terreus | n.t. | n.t. | 124.57 |
| 18495 | 3574 | MTN64 | A. terreus | n.t. | n.t. | 7.88 |
| 18496 | 3577 | MTN67 | A. terreus | n.t. | n.t. | n.t. |
| 18497 | 3578 | MTN68 | A. terreus | n.t. | n.t. | 62.05 |
| 18498 | 3634 | MTN71 | A. terreus | n.t. | n.t. | 7.16 |
| 18499 | 3582 | MTN73 | A. terreus | n.t. | n.t. | n.t. |
| 18500 | 3588 | MTN79 | A. terreus | n.t. | n.t. | 154.65 |
| 18501 | 3589 | MTN80 | A. terreus | n.t. | n.t. | 10.70 |
| 18502 | 3636 | MTN89 | A. terreus | n.t. | n.t. | n.t. |
| 18503 | 3601 | MTN94 | A. terreus | n.t. | n.t. | 0.20 |
| 18504 | 3602 | MTN95 | A. terreus | n.t. | n.t. | 510.56 |
| 18505 | 3603 | MTN97 | A. terreus | n.t. | n.t. | n.t. |
| 18506 | 3639 | MTN111 | A. terreus | n.t. | n.t. | n.t. |
| 18507 | 3615 | MTN113 | A. terreus | n.t. | n.t. | 59.46 |
| Location Name | Latitude | Longitude |
|---|---|---|
| Karun | 32.290077 | 48.601034 |
| Haft Tappeh | 32.104158 | 48.769371 |
| Imam Khomeini | 31.816312 | 48.835920 |
| Dehkhoda | 31.363858 | 48.754828 |
| Debal Khazaei | 31.151123 | 48.511743 |
| Amir Kabir | 31.030506 | 48.278718 |
| Salman Farsi | 30.974513 | 48.657927 |
| Mirza. K. Khan | 30.739912 | 48.066038 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavakol Noorabadi, M.; Babaeizad, V.; Zare, R.; Asgari, B.; Haidukowski, M.; Epifani, F.; Stea, G.; Moretti, A.; Logrieco, A.F.; Susca, A. Isolation, Molecular Identification, and Mycotoxin Production of Aspergillus Species Isolated from the Rhizosphere of Sugarcane in the South of Iran. Toxins 2020, 12, 122. https://doi.org/10.3390/toxins12020122
Tavakol Noorabadi M, Babaeizad V, Zare R, Asgari B, Haidukowski M, Epifani F, Stea G, Moretti A, Logrieco AF, Susca A. Isolation, Molecular Identification, and Mycotoxin Production of Aspergillus Species Isolated from the Rhizosphere of Sugarcane in the South of Iran. Toxins. 2020; 12(2):122. https://doi.org/10.3390/toxins12020122
Chicago/Turabian StyleTavakol Noorabadi, Maryam, Valiollah Babaeizad, Rasoul Zare, Bita Asgari, Miriam Haidukowski, Filomena Epifani, Gaetano Stea, Antonio Moretti, Antonio Francesco Logrieco, and Antonia Susca. 2020. "Isolation, Molecular Identification, and Mycotoxin Production of Aspergillus Species Isolated from the Rhizosphere of Sugarcane in the South of Iran" Toxins 12, no. 2: 122. https://doi.org/10.3390/toxins12020122
APA StyleTavakol Noorabadi, M., Babaeizad, V., Zare, R., Asgari, B., Haidukowski, M., Epifani, F., Stea, G., Moretti, A., Logrieco, A. F., & Susca, A. (2020). Isolation, Molecular Identification, and Mycotoxin Production of Aspergillus Species Isolated from the Rhizosphere of Sugarcane in the South of Iran. Toxins, 12(2), 122. https://doi.org/10.3390/toxins12020122