The Genomic Regions That Contain Ochratoxin A Biosynthetic Genes Widely Differ in Aspergillus Section Circumdati Species




Department of Genetics, Physiology, and Microbiology, Faculty of Biology, University Complutense of Madrid, Jose Antonio Nováis 12, 28040 Madrid, Spain
*
Author to whom correspondence should be addressed.
Toxins 2020, 12(12), 754; https://doi.org/10.3390/toxins12120754
Submission received: 30 October 2020
/
Revised: 22 November 2020
/
Accepted: 28 November 2020
/
Published: 29 November 2020
(This article belongs to the Section Mycotoxins)
Abstract
:Aspergillus section Circumdati includes 27 species, some of which are considered ochratoxin A (OTA) producers. However, there is considerable controversy about their potential OTA synthesis ability. In this work, the complete genomes of 13 species of Aspergillus section Circumdati were analyzed in order to study the cluster of OTA biosynthetic genes and the region was compared to those previously reported in A. steynii and A. westerdijkiae. The results obtained reveal that the genomes of some species in this section, including A. affinis, A. cretensis, A. elegans, A. muricatus, A. pulvericola, A. roseoglobulosus, and A. subramanianii, contain a potentially functional OTA biosynthetic cluster. Therefore, they might be able to synthesize the toxin. On the contrary, A. melleus, A. ochraceus, A. ostianus, A. persii, A. sclerotiorum, A. sesamicola, and A. westlandensis contain a truncated version of the cluster that lacks many of the genes involved in OTA biosynthesis, which might be related to their inability to produce OTA. The gain/loss pattern is different in all species, which suggests that the genetic evolution of this region might be due to independent events.
Keywords:
Aspergillus ochraceus; Aspergillus steynii; Aspergillus westerdijkiae; biosynthetic cluster; ochratoxin AKey Contribution: The potential of Aspergillus section Circumdati species to produce OTA has been attributed to the presence/absence of the genes involved in its biosynthesis. Some species that were considered OTA producers, such as A. ochraceus, might not form the toxin because they lack the biosynthetic cluster in their genomes.

1. Introduction
Ochratoxin A (OTA) is one of the most important mycotoxins due to its ubiquity as a natural contaminant in food and feed, and it exerts several toxic effects that pose clear risks to human and animal health [1]. Traditionally, cereals [2], grapes and their derivatives [3], and coffee [4] have been considered the main sources of OTA in the human diet. However, new insights into OTA occurrence have revealed that dietary exposure to this toxin might also be of importance in other products, such as preserved meat and cheese [5].
The evidence of OTA production was first reported in a culture of Aspergillus ochraceus (section Circumdati) [6], and for a long time, this species was considered the main OTA producer worldwide. Since then, many other Aspergillus and Penicillium species have been reported to be capable of synthesizing OTA, and currently, the list of OTA-producing species is rapidly increasing. In an extensive morphological, metabolic, and phylogenetic study, Frisvad et al. [7] described new species included in Aspergillus section Circumdati, and some of them were not included in the A. ochraceus clade. Several studies have indicated that two of the new species, A. westerdijkiae and A. steynii, are by far the most important OTA producers in this section, and several isolates previously identified as A. ochraceus, including the type species and the original OTA-producing strain, were reidentified as A. westerdijkiae [7,8,9]. Since then, different studies have extended the knowledge regarding the taxonomy of this section, and currently, 27 species are accepted in Aspergillus section Circumdati [10]. Recently, Houbraken et al. [11] included new taxonomical groups in the genus Aspergillus and separated section Circumdati into three series: Circumdati, Sclerotiorum, and Steyniorum. However, there is considerable controversy about the potential of all Aspergillus section Circumdati species to produce OTA. This ability has been revised in several studies, but it seems to be inconsistent in many species, and no clear conclusions have been reached except for the abovementioned cases (A. steynii and A. westerdijkiae) [10,12]. Aspergillus section Nigri also contains important OTA-producing species, such as A. carbonarius, A. niger, and A. welwitschiae [13], whereas, in the Penicillium genus, the most important OTA producers are P. nordicum and P. verrucosum [14]. The species Aspergillus alliaceus, included in section Flavi is also able to synthetize this toxin [15].
Despite the importance of OTA in food safety, its biosynthetic pathway is not entirely clear and some enzymatic steps are not fully understood [16]. This fact has complicated studies on the molecular and genetic basis of OTA production. For a long time, only a few isolated genes were demonstrated to be involved in OTA biosynthesis in Aspergillus species, as well as two small clustered regions in Penicillium [17]. However, in the last few years, the rapid evolution of molecular biology techniques, including next-generation sequencing, has significantly increased the number of available genome sequences of OTA-producing species, most of which have been deposited in databases. Therefore, in the last decade, many studies have focused on unraveling the genetic basis of OTA production. A clustered region of five genes is now known to be involved in OTA biosynthesis, and the same conserved synteny is maintained in both Aspergillus species (A. westerdijkiae, A. steynii, A. niger, A. welwitschiae, and A. carbonarius) and Penicillium nordicum [18,19]. These five genes encode different enzymes, including a halogenase (HAL), a cytochrome P450 monooxygenase (p450), a non-ribosomal peptide synthetase (NRPS), and a polyketide synthase (PKS) that can catalyze most of the steps of the OTA biosynthetic pathway, as well as a bZIP transcription factor (bZIP) that can regulate the expression of the rest of the clustered genes [20,21]. However, more studies are necessary to resolve existing contradictions on some processes of the biosynthetic pathway, to identify genes that might also be involved in OTA biosynthesis, and to explore how the route is fully regulated [20,21].
As mentioned before, the significance of the Aspergillus section Circumdati species as OTA producers has not been clearly established. Thus, the aim of this work was to perform an in-depth analysis of the molecular basis of OTA production in the most relevant Aspergillus section Circumdati species. The sequences of the complete biosynthetic clusters of A. westerdijkiae and A. steynii were obtained by our group in a previously published paper [18]. In this work, these regions were compared to those present in some relevant Aspergillus section Circumdati species. The genomes of A. affinis, A. cretensis, A. muricatus, A. ostianus, A. persii, A. roseoglobulosus, A. sclerotiorum, A. sesamicola, A. subramanianii, and A. westlandensis were retrieved from the JGI database, whereas A. elegans and A. melleus complete genomes were obtained in this work. The putative OTA biosynthetic cluster region of five A. ochraceus strains was also sequenced.
2. Results
2.1. Analysis of the OTA Biosynthetic Cluster in Aspergillus Section Circumdati Species Containing the Complete Cluster
The Aspergillus elegans genome was completely sequenced in this work and deposited in the NCBI database (BioProject number PRJNA679179). The total genome size was 36.78 Mb, and it was assembled into 8 contigs with a mean length of 4,597,187 bp. The BLAST search performed revealed a long region of 23680 bp that shared very high identity (99%) with the complete OTA biosynthetic cluster of A. steynii. When both sequences were aligned, only 120 bp differed between them, and most of these changes occurred in intergenic regions or gene introns. The presence of the complete biosynthetic OTA cluster was also demonstrated in other Aspergillus section Circumdati species whose genomes, including A. affinis, A. cretensis, A. muricatus, A. pulvericola, A. roseoglobulosus, and A. subramanianii, are available in the JGI database. Table 1 includes the most important features of these regions. The sequences of these clusters were compared with the OTA biosynthetic clusters described in the other section Circumdati species. In general, all of the regions found in this work shared higher identity with the corresponding cluster from A. steynii than that from A. westerdijkiae. The comparison performed revealed that most of the changes found in DNA sequences occurred in gene introns or intergenic sequences that, theoretically, do not affect gene expression. The bZIP binding sites required to activate gene transcription were also found upstream of the four genes in all of the studied species. The sequences of the HAL, BZIP, P450, NRPS, and PKS proteins were predicted from all of the genes of the biosynthetic cluster in all seven species. The changes in DNA sequences were neither modifications in the reading frames nor internal stop codons, and no important changes in the predicted proteins were found that might have affected any of the essential motifs. The sequences of the complete biosynthetic clusters, together with their corresponding gene features, were deposited in the NCBI databases, and their accession numbers are indicated in Table 1. Taking all of these aspects into account, the OTA biosynthetic cluster in A. affinis, A. cretensis, A. elegans, A. muricatus, A. pulvericola, A. roseoglobulosus, and A. subramanianii seems to be perfectly functional. Therefore, these species might be capable of producing OTA. Moreover, in all cases, the flanking regions corresponded to genes that encoded an alpha/beta hydrolase and an oxidoreductase, and therefore, the OTA biosynthetic cluster was in the same location in all species.
2.2. Analysis of the OTA Biosynthetic Cluster in Aspergillus Section Circumdati Species Containing the Truncated Cluster
The primer pair designed on the basis of the flanking regions of the OTA biosynthetic cluster of A. westerdijkiae and A. steynii yielded a clear band using the DNA of all A. ochraceus strains tested. The fragment was first sequenced in A. ochraceus CECT 2093, which is known to be a nonproducing strain [9]. After four rounds, a sequence of 3330 bp was obtained. This approach was also used to sequence the fragment amplified in A. ochraceus CECT 2969, CECT 2970, PDF2, and R1, and a similar length was obtained in all cases, with high identity values with the sequence obtained in A. ochraceus CECT 2093 (97–99%). When the sequence of this region was compared to the biosynthetic cluster of A. westerdijkiae, high identity values were observed in some regions (78–85%), although many parts of the cluster were absent in the A. ochraceus amplified region. A scheme that compares both regions is shown in Figure 1. Only four small parts of the biosynthetic cluster remained in A. ochraceus and corresponded to some parts of the HAL- and PKS-encoding genes. Moreover, a sequence gain seems to have occurred, since two additional regions were also found that shared no identity with any sequence available in databases. The same pattern of sequence gain/loss was found in the A. ostianus genome, although the length of each fragment was slightly different (Figure 1).
The complete genome of A. melleus was sequenced in this work in order to study the putative cluster involved in OTA biosynthesis in this species. The accession number of the BioProject in the NCBI database is PRJNA679179. A genome size of 38.85 Mb was found, and the genome was assembled into 11 contigs (mean length of 3,531,893 bp). After a BLAST search in the NCBI database, a region of 9384 bp was found to show identity with the OTA biosynthetic cluster in A. westerdijkiae. A comparison of both regions is shown in Figure 1. In this case, the region also appears to have lost some parts of the biosynthetic cluster, although the deletion pattern is different from that observed in A. ochraceus. In that case, the complete CDS of the PKS-encoding gene remains in the genome, together with some small parts of the HAL-encoding gene and a short sequence of the intergenic spacer between this gene and the bZIP transcription factor-encoding one. Three unknown small sequences also appear in this region in the A. melleus genome. This gain/loss pattern was also observed in the putative OTA cluster region found in the A. sesamicola genome (Figure 1).
The remaining parts of the OTA biosynthetic cluster found in A. westlandensis widely differ from those reported in the other species, and it is the only one that has not maintained the 3′-end of the PKS-encoding gene. In this case, the complete BZIP-encoding gene is present, as well as other intergenic regions that are located close to this gene. The gain of some regions was also observed (Figure 1).
All of the aforementioned species that contain a truncated version of the OTA biosynthetic cluster are classified in series Circumdati and share higher identity with the corresponding region of the A. westerdijkiae genome. However, the rest of the truncated versions of the OTA cluster found in Aspergillus section Circumdati species show the highest identity with the genomic region of A. steynii, and therefore, comparisons of this sequence were performed. This second group of species that contains the truncated cluster belongs to series Sclerotiorum. The presence of the OTA biosynthetic cluster within the genome of A. sclerotiorum was also studied. The A. sclerotiorum genome was found to contain a region of 9426 bp with different sequences that are similar to the OTA biosynthetic cluster of A. steynii, with identity values that range between 68 and 82% (Figure 2). These regions correspond to the complete CDS of the HAL-encoding gene, parts of the genes encoding BZIP and P450, and three short sequences of the PKS-encoding gene, including its 3′-end. The intergenic regions between HAL and bZIP as well as between BZIP and P450 are slightly longer, and no identity at all was found between them and those present in the A. steynii OTA cluster.
The partial OTA cluster found in A. persii maintained the same regions of the BZIP-, P450-, and PKS-encoding genes as A. sclerotiorum, although a phenomenon of genetic inversion was observed in the HAL-encoding gene (Figure 2). Whereas the 3′-end of the CDS was conserved in the correct direction, the 5′-end was found to be inverted from its original form, indicating that, apart from gain/loss events, the OTA cluster of A. persii underwent a rearrangement event.
The flanking regions of the partial OTA cluster were studied in the genomes of the species analyzed in this work that contained either the complete or the truncated version. In general, both genes encoding an alpha/beta hydrolase and an oxidoreductase were maintained, indicating that the location of the cluster is the same in all of the species. The only exception was found in the A. sclerotiorum genome, which maintained the alpha/beta hydrolase-encoding gene in one of the flanks, but it also lost the gene corresponding to the oxidoreductase-encoding gene.
2.3. Phylogenetic Analysis of Aspergillus Section Circumdati Related to the Presence of the OTA Biosynthetic Cluster
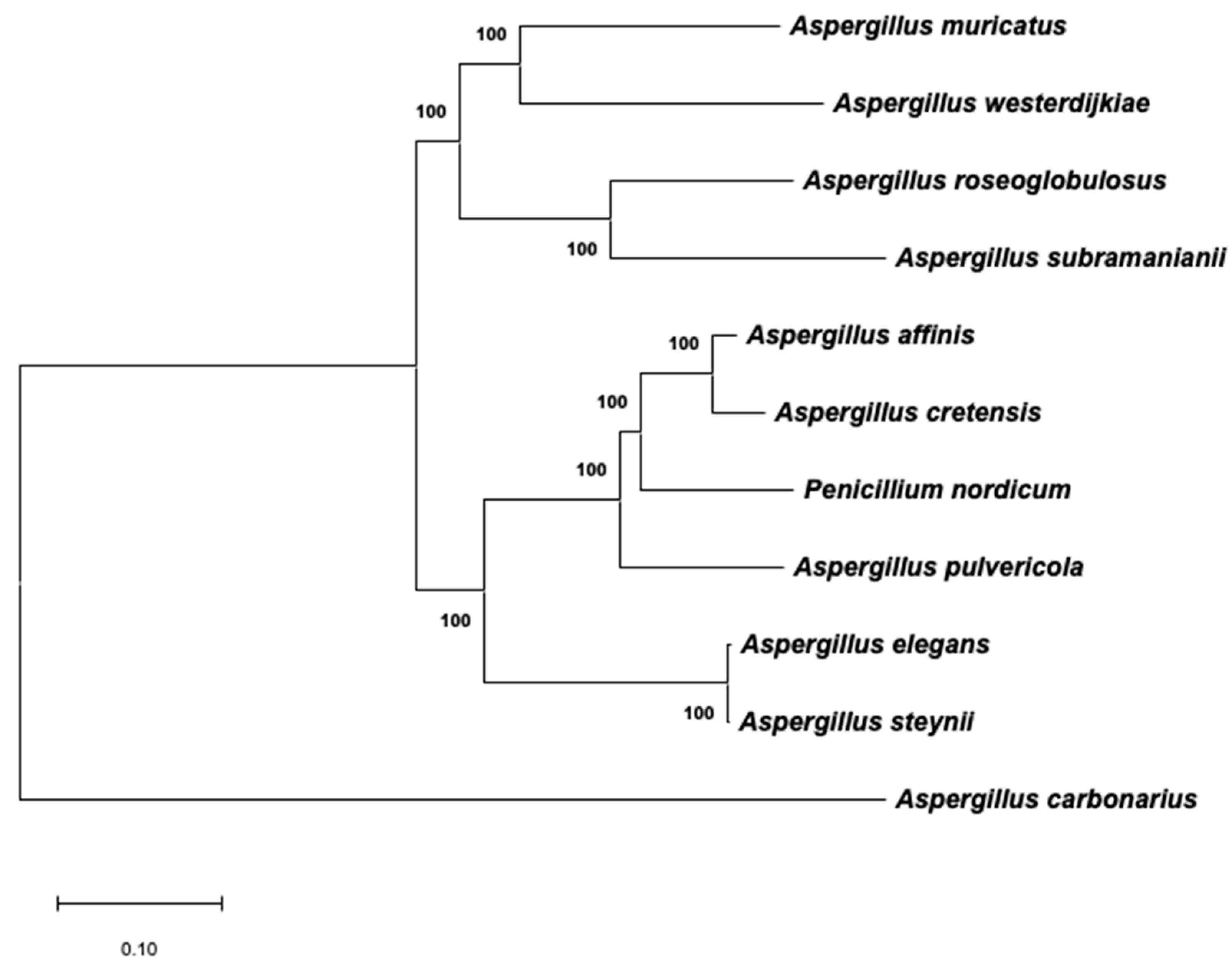
The evolutionary history, which was inferred using a maximum likelihood (ML) approach with the combined dataset of β-tubulin/ITS1-5.8S-ITS2/calmodulin, is shown in Figure 3. The best model to describe the substitution pattern of the sequences was Tamura Nei, and the rate variation among sites was modeled with a gamma distribution (shape parameter = 0.31). This phylogeny, based on the ML method, is a reflection of the real phylogeny of Aspergillus section Circumdati that was previously reported by other authors [7,22], and the branches are strongly supported by high bootstrap values. Near the branches, Figure 3 also shows a schematic that represents the sequences of the biosynthetic cluster present in the genome of the corresponding species. It is clear that the presence/absence of the complete cluster does not completely reflect the phylogeny of the section. Therefore, the gain/loss of the genes involved in OTA biosynthesis might be due to independent events.
Whereas the complete OTA biosynthetic region was maintained in all analyzed species of series Steyniorum, the presence of a partial cluster was frequent in series Circumdati and Sclerotiorum. Moreover, similar gain/loss patterns were observed in closely related species, such as A. sesamicola and A. melleus, A. ochraceus and A. ostianus, and A. persii and A. sclerotiorum.
A similar phylogeny using the same species was constructed on the basis of the partial 3′-end of the PKS-encoding gene that was found to remain in all of the species evaluated, except in the case of A. westlandensis. The corresponding ML tree is shown in Figure 4. Tamura 3-parameter was the best evolutionary model to describe the substitution pattern, and the rate variation among sites also followed a gamma distribution with a shape parameter of 0.61. As mentioned in the Materials and Methods section, it was not possible to include an appropriate outgroup taxon. Therefore, this unrooted phylogeny is not strictly comparable to that constructed using the β-tubulin/ITS1-5.8S-ITS2/calmodulin dataset. However, some conclusions can be made independently for the two main branches. It is clear that the topology of this tree reveals a higher evolution rate in the species that contain a nonfunctional version of the OTA cluster. In series Circumdati, the last branches corresponded to A. sesamicola, A. melleus, A. ostianus, and A. ochraceus, and the same situation arose in series Sclerotiorum, in which A. persii and A. sclerotiorum clustered together in the last branch. All six of the mentioned species contain a truncated version of the OTA cluster. It is important to highlight the cluster formed by A. affinis and A. cretensis, which were classified as series Circumdati but grouped together with the members of series Steyniorum in this tree. Moreover, the OTA-producing species P. nordicum was also included in this cluster.
The complete sequence of the OTA biosynthetic cluster found in Aspergillus section Circumdati species was also analyzed using an ML phylogeny. The corresponding sequence of P. nordicum was also included in the analysis. The ML tree obtained is shown in Figure 5. The General Time-Reversible model had the best ability to describe the substitution pattern. A discrete gamma distribution was used to model the evolutionary rate differences among sites (alpha parameter = 2.01) and allowed for some sites to be evolutionarily invariable (27.66%). This phylogenetic analysis revealed that the OTA clusters in A. westerdijkiae and A. muricatus were more related to that of series Sclerotiorum, since they diverged together from the same cluster. However, as occurred in the case of the analysis performed with the 3′-end of the PKS-encoding gene, A. affinis and A. cretensis were closer to the members of series Steyniorum than to their closely related species A. westerdijkiae and A. muricatus. Moreover, the two species clustered together with A. pulvericola and P. nordicum in a branch supported by high bootstrap values. This fact supports the hypothesis that P. nordicum acquired the biosynthetic cluster via horizontal transfer from an Aspergillus species included in section Circumdati [17] and reduces the number of the most probable donors to these three species.
3. Discussion
Aspergillus section Circumdati includes 27 species, which are able to produce a large variety of secondary metabolites [10]. As mentioned above, there is a lot of controversy regarding OTA production by Aspergillus section Circumdati species. In this work, the study of fungal genomes of different species revealed interesting data about the presence of the complete or truncated OTA biosynthetic cluster that might be related to their ability/inability to synthesize the toxin.
The inability to produce mycotoxins due to the deletion of some genes of the biosynthetic cluster has been frequently reported in the Aspergillus genus by several authors. Some strains of A. oryzae are not able to produce aflatoxins, and the loss of several genes involved in their synthesis has been reported [23]. Similarly, Adhikari et al. [24] reported that a number of A. flavus strains were not able to produce aflatoxins due to deletions of some genes involved in early biosynthesis stages. A similar situation occurred in the case of the fumonisin B2 biosynthetic cluster in Aspergillus niger aggregate species [19,25]. Therefore, deletion of parts of the biosynthetic cluster seems to be a common path related to the loss of mycotoxigenic ability in these species.
The partial deletion of the cluster of genes involved in OTA biosynthesis has also been recently studied in Aspergillus niger aggregate species [19,25]. In all cases, the presence of this truncated cluster is related to the inability of the isolates to produce the toxin, and, to our knowledge, it seems to occur in most A. niger and A. welwitschiae strains and all Aspergillus tubingensis. In all cases, this long deletion in Aspergillus niger aggregate species is accompanied by the maintenance of a region that is approximately 600 bp long, at the end of the biosynthetic cluster, which corresponds to the 3′-region of the PKS gene. This aspect might be assumed to be related to a single deletion event that eliminated almost the complete cluster. However, in the case of the species of Aspergillus section Circumdati, some parts of the genes or intergenic regions of the biosynthetic cluster remain in the genome, and these parts are different in each species, which could be related to subsequent gains and losses over time. These multiple independent loss events have also been described in the deletion of the bikaverin biosynthetic cluster in Botrytis spp. [26].
Aspergillus ochraceus was the first reported OTA-producing species and, for a long time, it was considered an important contributor to OTA levels in foodstuffs. However, due to frequent changes in taxonomy, its relative importance drastically dropped with the emergence of A. westerdijkiae and A. steynii as the main producers in section Circumdati [7,9]. In this work, A. ochraceus was analyzed to elucidate the genes involved in OTA production in this species. However, surprisingly, the typical clusters that are present in both Aspergillus and Penicillium species, as reported previously by our group [18], were not found, and only small remaining parts of some genes were present in the A. ochraceus genome. Due to the importance of this species as an OTA producer, these results were confirmed in five different strains from different origins. The same deletion patterns were found in all of the isolates studied, indicating a consistent lack of the ability to produce OTA in this species. All of these isolates had been previously classified as nonproducing strains of Aspergillus ochraceus [9]. However, some recently published papers have described some A. ochraceus isolates that are able to produce OTA, although it is true that they do so inconsistently and yield very low levels. As mentioned before, identification of Aspergillus section Circumdati species is especially difficult and often requires the application of molecular techniques [8,12]. Some papers that have recently described these OTA-producing isolates have performed their identification based only on morphological characteristics, which has been demonstrated to be insufficient [27,28,29,30,31,32,33]. Therefore, it might be possible that they were misidentified and were not really A. ochraceus, but instead other species, such as A. steynii or A. westerdijkiae, that are more relevant OTA producers in foodstuffs. Similarly, Wang et al. [34] recently published the genome of A. ochraceus fc-1 and reported the presence of the complete OTA biosynthetic cluster and the ability of this strain to produce the toxin. However, the analysis of this genome, which is available on the NCBI database, revealed a misidentification of the strain fc-1 that should be reclassified as A. westerdijkiae on the basis of β-tubulin and ITS1-5.8S-ITS2 sequences. On the other hand, other papers that recently described A. ochraceus as an OTA producer evaluated toxin concentrations using HPLC methods and have detected extremely low levels of OTA in the medium, which have been very close to detection limits [35]. It is known that some fungi are able to form some compounds that might be misidentified as OTA using HPLC. This was the case for A. tubingensis, which led to the species being considered an OTA producer for a long time [25,36].
Apart from A. ochraceus, six other Aspergillus section Circumdati species contain a truncated version of the OTA biosynthetic cluster: A. melleus, A. ostianus, A. persii, A. sclerotiorum, A. sesamicola, and A. westlandensis. The ability of these species to produce OTA has been frequently described as inconsistent, and previous studies have indicated that only trace levels of the toxin could be detected, even in highly permissive conditions [7,10,12]. Our results demonstrate that these species may not be OTA producers since they do not contain the biosynthetic cluster in their genomes.
Several hypotheses on the reason why mycotoxin biosynthetic clusters are lost over evolution have been proposed, and their common tenet is that adaptation to new situations might be decisive. It is well known that gene loss has been crucial to fungal evolution and, in some cases, is correlated with shifts in ecological niches or association with a host [37]. Campbell et al. [26] suggested that the acquisition of the bikaverin cluster might have provided a selective advantage that allowed Botrytis spp. to survive as saprotrophs in the soil. However, as Botrytis genomes evolved toward virulent, endophytic strains, bikaverin production was no longer retained. A similar fact was observed in the case of Penicillium roqueforti and its inability to produce mycophenolic acid due to a short deletion of 174 bp in the biosynthetic cluster [38]. All of the strains that do not harbor the capacity to produce this toxin are isolated from blue cheese. The authors hypothesized that these strains may no longer require this trait, which has a high energetic cost, as an evolutionary advantage to compete with other organisms. For this reason, they do not maintain the ability to produce mycophenolic acid. This idea of ecological adaptation due to the domestication of strains was also proposed by Martín and Liras [39] regarding the inability of P. roqueforti to produce meleagrin and neoxalin. These authors affirmed that the production of these toxins did not confer an ecological advantage to the fungus, although they asserted that the deletion of the genes is not a recent event, since the deletion is consistent in isolates from different origins and types of cheese.
In the last few years, different studies have been published regarding the relative significance of Aspergillus spp. in human pathologies. In this context, some Aspergillus section Circumdati species, including A. melleus, A. ochraceus, A. persii, and A. sclerotiorum, were described as causal agents of onychomycosis [40]. All of these species contain only a short fragment of the OTA cluster in their genomes, and adaptation to this unusual habitat might be related to the loss of their ability to produce the toxin because it does not have a useful function. Because they were recently described as new species in Aspergillus section Circumdati, there are no reports on the occurrence of A. westlandensis or A. sesamicola, and no hypothesis addresses their inability to produce OTA. However, it is important to highlight A. westlandensis, since it presents a unique pattern in the remaining regions of the OTA cluster that might be related to independent events during its speciation.
The analysis of fungal genomes revealed the presence of the complete OTA biosynthetic cluster in seven Aspergillus section Circumdati species, which, together with A. steynii and A. westerdijkiae, should be considered as possible OTA producers within this section. As mentioned above, several studies have reported the ability of these species to produce OTA. In the most recently published study, Visagie et al. [10] indicated that A. affinis, A. cretensis, A. muricatus, A. pulvericola, and A. roseoglobulosus are consistent OTA producers, since the authors detected large amounts of OTA in fungal cultures. This fact is in agreement with the results of this work, in which the presence of a putative functional OTA cluster was detected in their genomes, which indicates their potential ability to synthesize the toxin. To date, few reports have studied the natural occurrence of these species, which, in some cases, is due to their recent description. For example, A. pulvericola has only been reported in old, deteriorated manuscripts [41], whereas A. affinis has been detected in decomposing leaves and in a cave environment [42,43]. Therefore, it is essential to perform exhaustive surveys to study the presence of these potential OTA-producing species in foodstuffs to establish the risk that they might pose to food security and human health.
Aspergillus subramanianii and A. elegans are considered inconsistent OTA producers because the toxin has been detected in only some cases and at low levels in fungal cultures [9,10]. The in silico analysis performed in this work suggests that the enzymes encoded by the clustered genes that are involved in toxin production might be functional. However, some atoxigenic strains have also been described in A. carbonarius, which contains no large deletions in the OTA biosynthetic cluster. However, all of the genes are downregulated, even under highly permissive conditions [44]. Even containing the complete cluster in their genomes, some atoxigenic strains of A. westerdijkiae and A. steynii have also been described, and no expression has been detected in the five genes involved in OTA biosynthesis [9,18]. Furthermore, some authors have suggested that post-transcriptional modifications might also interfere in the regulation of mycotoxin biosynthesis. These mechanisms include microRNA and virus-mediated modulation, among others [45,46]. Thus, the ability of A. subramanianii and A. elegans to produce OTA should be studied in detail to unravel whether some regulatory mechanisms are affected.
As mentioned before, Adhikari et al. [24] related the inability to produce aflatoxins to deletions of some genes in the biosynthetic cluster. The deletion patterns allowed A. flavus isolates to be classified into six different groups that revealed a consistent evolutionary relationship between most of the genotypes, suggesting distinct deletion events for each group. However, in our study, no evident evolutionary relationships were found between Aspergillus section Circumdati species and the loss of the OTA cluster. In comparing the presence/absence of the biosynthetic cluster in Aspergillus section Circumdati species with the real phylogeny constructed based on β-tubulin, ITS1-5.8S-ITS2, and calmodulin regions, we observed no obvious patterns, which suggests that the evolution of this region might be due to independent gain/loss events during speciation. However, it is interesting to highlight that all tested species of series Steyniorum have maintained the complete OTA biosynthetic cluster, and therefore, it seems to be a common threat in this series. In this work, only one isolate was tested in most of the species. Therefore, it would be interesting to confirm whether deletion patters present intraspecific variability. Another relevant hypothesis that could be made regarding phylogenetic analysis is the origin of the P. nordicum cluster, which is known to have acquired the cluster by horizontal transfer from Aspergillus species [18]. To date, the highest identity found is between P. nordicum and A. steynii OTA clusters, and the hypothesis indicates that this section Circumdati species might have been the donor to the Penicillium species. However, phylogenetic analysis clustered P. nordicum with A. affinis and A. cretensis, showing that they share extremely high identity in their OTA biosynthetic regions (83% and 80%, respectively). Therefore, these results suggest that the transference of the OTA biosynthetic cluster to the Penicillium genus might have occurred from one of these two Aspergillus section Circumdati species.
The study of the gain/loss pattern of the OTA biosynthetic cluster revealed that, except for A. westlandensis, a short region of the 3′-end of the cluster was maintained in all species of Aspergillus section Circumdati and in section Nigri. The phylogenetic analysis performed using the sequences of this region in Aspergillus section Circumdati reflects that the evolutionary history of the section, as revealed by the analysis of β-tubulin/ITS1-5.8S-ITS2, is similar in some groups but widely different in others. This aspect might suggest that the cluster was present in the genome of the common ancestor of Aspergillus section Circumdati species and then evolved independently during speciation. The reason that both nonproducing Aspergillus section Nigri and Circumdati species have maintained the 3′-end of the PKS-encoding gene remains unknown, and its importance needs to be addressed.
As mentioned before, many taxonomical revisions of Aspergillus section Circumdati have been published in the last few years in which new species were described and some isolates were reclassified. Therefore, currently, it is difficult to accurately determine the habitat of each species. However, many authors have suggested that A. westerdijkiae and A. steynii are probably the most frequent species that occur in foodstuffs in the section [12,47], whereas the rest of the species occur in other ecological environments [48]. It is not completely clear why fungi produce mycotoxins, but some evidence indicates that they provide a competitive advantage to defend their niche from other species [49]. It might be thought that A. steynii and A. westerdijkiae have retained their ability to produce OTA to effectively colonize food products, whereas the species more adapted to other less-specialized environments might have lost the cluster to avoid extreme energy expenses due to OTA production.
4. Conclusions
The results presented here provide insight into the genetic basis of OTA production in the most important Aspergillus section Circumdati species. The Aspergillus ochraceus strains analyzed in this work did not contain the OTA biosynthetic cluster in their genome. Therefore, this species might be excluded as an OTA producer, in which case studies on the occurrence of this fungus are no longer relevant. Moreover, it is crucial to carry out large surveys regarding the occurrence of species that are demonstrated to be potential OTA producers in food products to unravel their relative importance in food safety, such as A. affinis, A. cretensis, A. elegans, A. muricatus, A. pulvericola, and A. subramanianii. It would be also interesting to confirm the lack of intraspecific variation and whether all the isolates of these species are potential OTA producers. Due to the fact that the loss of the cluster seems to be the most frequent mechanism of the inability to produce OTA in Aspergillus species, this region might be used to design molecular detection methods based on PCR for the early detection of OTA producers in foodstuffs.
5. Materials and Methods
5.1. Putative OTA Cluster Determination in Aspergillus ochraceus Strains
Five Aspergillus ochraceus strains were used to study the presence of the OTA biosynthetic cluster in fungal genomes. These strains included the type strain CECT 2093, other isolates obtained from Spanish Type Culture Collection (CECT 2969 and CECT 2970), and two A. ochraceus obtained from previous works in our laboratory (PDF2 and R1). The inability of these strains to produce has been demonstrated before [9].
Genomic DNA was isolated from 3-day-old cultures in Sabouraud Broth. Mycelia were harvested by filtration using Whatman Paper Nº1, frozen with liquid nitrogen, and ground using a mortar and a pestle. DNA isolation was performed using the protocol described elsewhere [50]. DNA concentrations were determined using a NanoDrop® ND-1000 spectrophotometer (Nanodrop Technologies, Wilmington, NC, USA).
A new set of degenerate primers were designed on the basis of the sequences of flanking genes of the OTA biosynthetic cluster in A. steynii and A. westerdijkiae, and they amplified the region located between the 3′-end of the alpha/beta hydrolase and the 5′-end of the oxidoreductase. Therefore, the primer pair interHID-F (5′-CGTGTGATTGGGAKCGG-3′) and interOXI-R (5′-CTGGSGATGGRAGTCGA-3′) was used to amplify the region in A. ochraceus strains using the following PCR conditions: 1 cycle of 5 min at 95 °C; 30 cycles of 1 min at 94 °C, 1 min at 63 °C, and 2 min at 72 °C; and a final extension of 5 min at 72 °C. The amplification reactions were carried out in volumes of 25 μL, containing 100 ng of sample DNA, 1 μL of each primer (20 μM; Metabion, Planegg, Germany), and 12.5 μL of NZYTaq II 2x Green Master Mix (Nzytech, Lisbon, Portugal).
PCR products were detected in 1.5% agarose gels in TAE 1X buffer (40 mM Tris-acetate and 1.0 mM EDTA) with 0.002% GreenSafe Premium stain (Nzytech, Lisbon, Portugal). The NZYDNA Ladder VII (Nzytech, Lisbon, Portugal) was used as a molecular size marker. PCR products were purified using the kit NZYGelpure (Nzytech, Lisbon, Portugal).
Due to the length of the PCR products obtained and the impossibility of obtaining a sequence of quality using the primer interOXI-R, different rounds of sequencing had to be performed. The first sequencing step was carried out using the primer interHID-F, and the complete sequence was subsequently obtained using interOCRAseq1 (5′-GACACCGAGTCATGCAGCGTC-3′), interOCRAseq2 (5’-GGCTGTTGTTTATCTGATCTGACA-3′), and interOCRAseq3 (5′-GACTCGTTAGTGGCAGTGGACTT-3′). PCR products were sequenced in Macrogen Facilities (Madrid, Spain) using an ABI 3730XL DNA sequencer (Applied Biosystems, Foster City, CA, USA). Sequences were edited with Unipro UGENE bioinformatics software version 1.25.0 (Unipro, Novosibirsk, Russia) [47].
5.2. Genome Sequencing of Aspergillus melleus and Aspergillus elegans
The complete genome sequence of A. elegans IMI 345568 and A. melleus CBS 546.65 were obtained by next-generation sequencing. Genomic DNA was isolated as described in the previous subsection. A hybrid sequencing approach was applied to obtain the highest quality results. This method combined the PacBio long-read platform and Illumina sequencing. The DNA sample was used for library construction using the PacBio SMRTbell Library Preparation Kit (Pacific Biosciences, Menlo Park, CA, USA) with a 20 kb insert size and was subsequently analyzed in a PacBio RSII sequencer (Pacific Biosciences, Menlo Park, CA, USA). In the case of Illumina data, the sequencing library was prepared using the TruSeq DNA Whole genome library preparation kit (Applied Biosystems, Foster City, CA, USA), and sequencing was performed in a NovaSeq 6000 system (Applied Biosystems, Foster City, CA, USA). The de novo assembly of PacBio reads was performed using the Hierarchical Genome Assembly Process (HGAP4). Then, Illumina reads were applied for accurate genome sequence using Pilon.
5.3. Sequence Analysis and Putative Cluster Identification in Other Aspergillus Section Circumdati Species
The complete genome sequences of Aspergillus affininis CBS 129190, Aspergillus cretensis CBS 112802, Aspergillus muricatus CBS 112808, Aspergillus ostianus CBS 103.07, Aspergillus persii CBS 112795, Aspergillus pulvericola CBS 137327, Aspergillus roseoglobulosus CBS 112800, A. sclerotiorum CBS 549.65, Aspergillus sesamicola CBS 137324, Aspergillus subramanianii CBS 138230, and Aspergillus westlandensis CBS 123905 are available on the JGI Genome Portal website and are a part of the Aspergillus whole-genome sequencing project started in 2013 by Scott E. Baker. The correct links to access these genomes are included in Table S1. The presence of the putative region containing the OTA cluster in these fungal genomes was tested using the JGI Alignment Search BLAST tool, available on the website, using the sequence of the OTA biosynthetic cluster of A. steynii CBS 112.812 (accession number MG701897) as the query.
5.4. Bioinformatics Analysis for Ochratoxin A Biosynthetic Cluster Comparisons
The putative regions corresponding to the biosynthetic cluster in A. steynii and A. westerdijkiae and the truncated cluster obtained in A. melleus, A. ochraceus, A. ostianus, A. persii, A. sclerotiorum, and A. sesamicola were compared using the bl2seq tool available at the NCBI website. The percentage of identity of the different regions of the sequences was also obtained using this tool.
Other bioinformatics analyses were performed if the complete cluster was observed in fungal genomes. Identity calculations and intron locations were carried out using Unipro UGENE 1.25.0 [51]. Protein sequence predictions were carried out using the Translate tool available at the Bioinformatics Resource Portal of the Swiss Institute of Bioinformatics (https://web.expasy.org/translate/), whereas domain analyses were performed with the PFAM 31.0 software available on the EMBL webpage (http://pfam.xfam.org).
5.5. Phylogenetic Analysis
DNA sequences were aligned using the iterative MUSCLE algorithm. Subsequently, the relationships among the species were visualized using a maximum likelihood (ML) tree. Three datasets were constructed to study the relationships among the taxa. In all cases, the corresponding sequences of Aspergillus carbonarius were used as the outgroup taxon. The first one included a combined dataset with the partial β-tubulin gene, the ITS1-5.8S-ITS2 region, and the partial sequence of the calmodulin gene, because they were reported as phylogenetically informative in evaluating interspecific relationships in Aspergillus section Circumdati. The sequences used to construct this phylogenetic tree are available in the NCBI database, and their accession numbers are included in Table 2. There were a total of 1233 positions in the final dataset.
The second dataset included a short sequence of the 3′-end of the PKS gene, which was found to be conserved in the genome of all the species analyzed except in A. westlandensis. This dataset was constructed with the sequences obtained in this work, as described above, together with others used in previous works in our laboratory [18]. In this case, it was not possible to find an appropriate outgroup taxon because of the high divergence between the sequences in Aspergillus section Circumdati and Nigri, which include other species that are able to produce OTA. Therefore, the tree constructed from the 3′-end of the PKS gene was unrooted. There was a total of 624 positions in this final dataset.
The last dataset corresponds to the sequences of the OTA biosynthetic genes and includes all of the species that were found to contain the complete cluster in their genomes. The corresponding sequence of A. carbonarius was used as the outgroup taxon. This final dataset was formed by 19,747 positions.
The data were first evaluated in order to establish the best evolutionary model. Subsequently, an ML tree was constructed using MEGA X [52]. The trees were drawn at scale, with branch lengths measured in the number of substitutions per site. Initial trees for the heuristic search were obtained automatically by Neighbor-Join and BioNJ algorithms applied to a matrix of pairwise distances estimated using the corresponding model. Then, the topology with the best log-likelihood value was selected. All positions containing gaps and missing data were eliminated from the analysis. Bootstrap values were calculated from 1000 replications of the bootstrap analysis to determine the support for the clades.
The analyses involved 17, 16, and 11 nucleotide sequences in the cases of β-tubulin/ITS1-5.8S-ITS2/calmodulin, the 3′-end of the PKS, and the complete cluster datasets, respectively.
Supplementary Materials
The following are available online at https://www.mdpi.com/2072-6651/12/12/754/s1: Table S1: Genomes used in this study, together with their accession links and their years of publication.
Author Contributions
All authors conceived the experimental design. J.G.-S. performed the laboratory analysis. J.G.-S. and B.P. wrote the original draft. B.P. and C.V. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministry of Science and Innovation, grant number RTI2018-097593-B-C21.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Microbiol. 2010, 27, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Wine contamination with ochratoxins: A review. Beverages 2018, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Leitao, A.L. Occurrence of ochratoxin A in coffee: Threads and solutions—A mini-review. Beverages 2019, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Contaminants in the Food Chain. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, 6113. [Google Scholar]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section Circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Gil-Serna, J.; Vázquez, C.; Sardiñas, N.; González-Jaén, M.T.; Patiño, B. Discrimination of the main ochratoxin A-producing species in Aspergillus section Circumdati by specific PCR assays. Int. J. Food Microbiol. 2009, 136, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; Sardiñas, N.; González-Jaén, M.T.; Patiño, B. Revision of ochratoxin A production capacity by the main species of Aspergillus section Circumdati. Aspergillus steynii revealed as the main risk of OTA contamination. Food Control 2011, 22, 343–345. [Google Scholar] [CrossRef]
- Visagie, C.M.; Varga, J.; Houbraken, J.; Meijer, M.; Kocsubé, S.; Yilmaz, N.; Fotedar, R.; Seifert, K.A.; Frisvad, J.C.; Samson, R.A. Ochratoxin production and taxonomy of the yellow aspergilli (Aspergillus section Circumdati). Stud. Mycol. 2014, 78, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Perrone, G.; Gallo, A. Aspergillus species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Springer: New York, NY, USA, 2017; pp. 33–49. [Google Scholar]
- Cabañes, F.J.; Bragulat, M.R. Black aspergilli and ochratoxin A-producing species in foods. Curr. Opin. Food Sci. 2018, 23, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Geisen, R.; Schmidt-Heydt, M.; Stoll, D.; Touhami, N. Aspects of the occurrence, genetics, and regulation of biosynthesis of the three food relevant Penicillium mycotoxins: Ochratoxin A, citrinin and patulin. In The Mycota XV. Physiology and Genetics, 2nd ed.; Anke, T., Schüffler, A., Eds.; Springer: New York, NY, USA, 2018; pp. 413–433. [Google Scholar]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.-B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenár, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Ferrara, M.; Perrone, G. Recent advances on the molecular aspects of ochratoxin A biosynthesis. Curr. Opin. Food Sci. 2017, 17, 49–56. [Google Scholar] [CrossRef]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [Green Version]
- Gil-Serna, J.; García-Díaz, M.; González-Jaén, M.T.; Vázquez, C.; Patiño, B. Description of an orthologous cluster of ochratoxin A biosynthetic genes in Aspergillus and Penicillium species. A comparative analysis. Int. J. Food Microbiol. 2018, 268, 35–43. [Google Scholar] [CrossRef]
- Susca, A.; Proctor, R.H.; Morelli, M.; Haidukowski, M.; Gallo, A.; Logrieco, A.F.; Moretti, A. Variation in fumonisin and ochratoxin production with differences in biosynthetic gene content in Aspergillus niger and A. welwitschiae isolates from multiple crop and geographic origins. Front. Microbiol. 2016, 7, 1412. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Gil-Serna, J.; Vázquez, C.; Patiño, B. Genetic regulation of aflatoxin, ochratoxin A, trichothecene, and fumonisin biosynthesis: A review. Int. Microbiol. 2020, 23, 89–96. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef]
- Kusumoto, K.I.; Nogata, Y.; Ohta, H. Directed deletions in the aflatoxin biosynthesis gene homolog cluster of Aspergillus oryzae. Curr. Genet. 2000, 37, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.N.; Bandyopadhyay, R.; Cotty, P.J. Degeneration of aflatoxin gene clusters in Aspergillus flavus from Africa and North America. AMB Express 2016, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Gil-Serna, J.; García-Díaz, M.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Significance of Aspergillus niger aggregate species as contaminants of food products in Spain regarding their occurrence and their ability to produce mycotoxins. Food Microbiol. 2019, 82, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.A.; Staats, M.; van Kan, J.A.L.; Rokas, A.; Slot, J.C. Repeated loss of an anciently horizontally transferred gene cluster in Botrytis. Mycologia 2013, 105, 1126–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernane, F.; Cano-Sancho, G.; Sanchis, V.; Marín, S.; Ramos, A.J. Aflatoxins and ochratoxin A in pistachios sampled in Spain: Occurrence and presence of mycotoxigenic fungi. Food Addit. Contam. Part B Surveill. 2010, 3, 185–192. [Google Scholar] [CrossRef]
- Jeswal, P.; Kumar, D. Mycofloral association and co-occurrence of ochratoxin A and citrinin in aromatic herbs and spices from Bihar state (India) detected by ELISA and LC-MS/MS. Am. J. Biol. Chem. Pharm. Sci. 2013, 1, 96–105. [Google Scholar]
- Priyanka, S.R.; Venkataramana, M.; Kumar, G.P.; Rao, V.K.; Murali, H.C.S.; Batra, H.V. Occurrence and molecular detection of toxigenic Aspergillus species in food grain samples from India. J. Sci. Food Agric. 2014, 94, 537–543. [Google Scholar] [CrossRef]
- Jeswal, P.; Kumar, D. Mycobiota and natural incidence of aflatoxins, ochratoxin A, and citrinin in Indian spices confirmed by LC-MS/MS. Int. J. Microbiol. 2015, 242486. [Google Scholar] [CrossRef] [Green Version]
- Nyongesa, B.W.; Okoth, S.; Ayugi, V. Identification key for Aspergillus species isolated from maize and soil of Nandi County, Kenia. Adv. Microbiol. 2015, 5, 55282. [Google Scholar] [CrossRef] [Green Version]
- Pandit, P.; Panta, O.; Karki, T. Isolation of Aspergillus ochraceus and production of ochratoxin in coffee samples. Nepal J. Sci. Technol. 2015, 15, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Orole, O.O. Ochratoxin A production by cocoa infested Aspergillus species in Ondo State, Nigeria. Biotechnol. J. Int. 2016, 12, 1–7. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.M.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A consensus ochratoxin A biosynthetic pathway: Insights from the genome sequence of Aspergillus ochraceus and a comparative genome analysis. App. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Shi, J.; Jiang, C.; Liu, Y. Inhibition of the growth and ochratoxin A production by Aspergillus carbonarius and Aspergillus ochraceus in vitro and in vivo through antagonistic yeasts. Food Control 2015, 50, 125–132. [Google Scholar] [CrossRef]
- Storari, M.; Bigler, L.; Gessler, C.; Broggini, G.A.L. Assessment of the ochratoxin A production ability of Aspergillus tubingensis. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk. Assess. 2012, 29, 1450–1454. [Google Scholar] [CrossRef]
- Stajich, J.E. Fungal genomes and insights into the evolution of the kingdom. Microbiol. Spectr. 2018, 5, 1–25. [Google Scholar]
- Gillot, G.; Jany, J.L.; Domínguez-Santos, R.; Poirier, E.; Debaets, S.; Hidalgo, P.I.; Ullán, R.V.; Coton, E.; Coton, M. Genetic basis for mycophenolic acid production and strain-dependent production variability in Penicillium roqueforti. Food Microbiol. 2017, 62, 239–250. [Google Scholar] [CrossRef]
- Martín, J.F.; Liras, P. Evolutionary formation of gene clusters by reorganization: The meleagrin/roquefortine paradigm in different fungi. Appl. Microbiol. Biotechnol. 2016, 100, 1579–1587. [Google Scholar] [CrossRef]
- Frías-De-León, M.G.; Espinosa-Hernández, V.M.; Bonifaz, A.; Martínez-Herrera, E. Onychomycosis due to Aspergillus spp.: A current review. Curr. Fungal Infect. Rep. 2018, 12, 112–119. [Google Scholar] [CrossRef]
- Oetari, A.; Susetyo-Salim, T.; Sjamsuridzal, W.; Suherman, E.A.; Monica, M.; Wongso, R.; Fitri, R.; Nurlaili, D.G.; Ayu, D.C.; Teja, T.P. Occurrence of fungi on deteriorated old dluwang manuscripts from Indonesia. Int. Biodeterior. Biodegrad. 2016, 114, 94–103. [Google Scholar] [CrossRef]
- Davolos, D.; Persiani, A.M.; Pietrangeli, B.; Ricelli, A.; Maggi, O. Aspergillus affinis sp. nov., a novel ochratoxin A-producing Aspergillus species (section Circumdati) isolated from decomposing leaves. Int. J. Syst. Evol. Microbiol. 2012, 62, 1007–1015. [Google Scholar] [CrossRef]
- Nováková, A.; Hubka, V.; Valinová, S.; Kolarík, M.; Hillebrand-Voiculescu, A.M. Cultivable microscopic fungi from an underground chemosynthesis-based ecosystem: A preliminary study. Folia Microbiol. 2018, 63, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Castellá, G.; Bragulat, M.R.; Puig, L.; Sanseverino, W.; Cabañes, F.J. Genomic diversity in ochratoxigenic and non ochratoxigenic strains of Aspergillus carbonarius. Sci. Rep. 2018, 8, 5439. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Lan, F.; Yang, W.; Zhang, F.; Yang, K.; Li, Z.; Gao, P.; Wang, S. sRNA profiling in Aspergillus flavus reveals differentially expressed miRNA-like RNAs response to water activity and temperature. Fungal Genet. Biol. 2015, 81, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Chitara, W.; Siciliano, I.; Gaiotti, F.; Ciuffo, M.; Forgia, M.; Varese, G.C.; Turina, M. Mycoviruses mediate mycotoxin regulation in Aspergillus ochraceus. Environ. Microbiol. 2018, 21, 1957–1968. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Baranyi, N.; Chandrasekaran, M.; Vágvölgyi, C.; Kocsubé, S. Mycotoxin producers in the Aspergillus genus: An update. Acta Biol. Szeged. 2015, 59, 151–167. [Google Scholar]
- Abdel-Azeem, A.M.; Abdel-Azeem, M.A.; Abdul-Hadi, S.Y.; Darwish, A.G. Aspergillus: Biodiversity, ecological significances and industrial applications. In Recent Advancement in White Biotechnology through Fungi; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: New York, NY, USA, 2019; pp. 121–179. [Google Scholar]
- Magan, N.; Aldred, D. Why do fungi produce mycotoxins? In Food Mycology. A Multifaceted Approach to Fungi and Food; Dijksterhuis, J., Samson, R.A., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 121–133. [Google Scholar]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knya, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
Figure 1.
Comparison of the OTA biosynthetic cluster in A. westerdijkiae (above) with the corresponding regions found in A. melleus, A. ochraceus, A. ostianus, A. sesamicola, and A. westlandensis. The schemes are drawn at scale, except in the cases of black regions marked by an asterisk (*), which are slightly longer than represented. The length of the complete region in fungal genomes is indicated below the corresponding species name. Gray regions correspond to sequences that share high identity with the OTA biosynthetic cluster and are located under the corresponding homologous region in A. westerdijkiae. Black regions share no identity with any sequence available in databases. The percentages below the boxes indicate the identity of the region with A. westerdijkiae. HAL = halogenase-, BZIP = bZIP transcription factor-, P450 = cytochrome P450 monooxygenase-, NRPS = non-ribosomal peptide synthetase-, and PKS = polyketide synthase-encoding genes.
Figure 1.
Comparison of the OTA biosynthetic cluster in A. westerdijkiae (above) with the corresponding regions found in A. melleus, A. ochraceus, A. ostianus, A. sesamicola, and A. westlandensis. The schemes are drawn at scale, except in the cases of black regions marked by an asterisk (*), which are slightly longer than represented. The length of the complete region in fungal genomes is indicated below the corresponding species name. Gray regions correspond to sequences that share high identity with the OTA biosynthetic cluster and are located under the corresponding homologous region in A. westerdijkiae. Black regions share no identity with any sequence available in databases. The percentages below the boxes indicate the identity of the region with A. westerdijkiae. HAL = halogenase-, BZIP = bZIP transcription factor-, P450 = cytochrome P450 monooxygenase-, NRPS = non-ribosomal peptide synthetase-, and PKS = polyketide synthase-encoding genes.
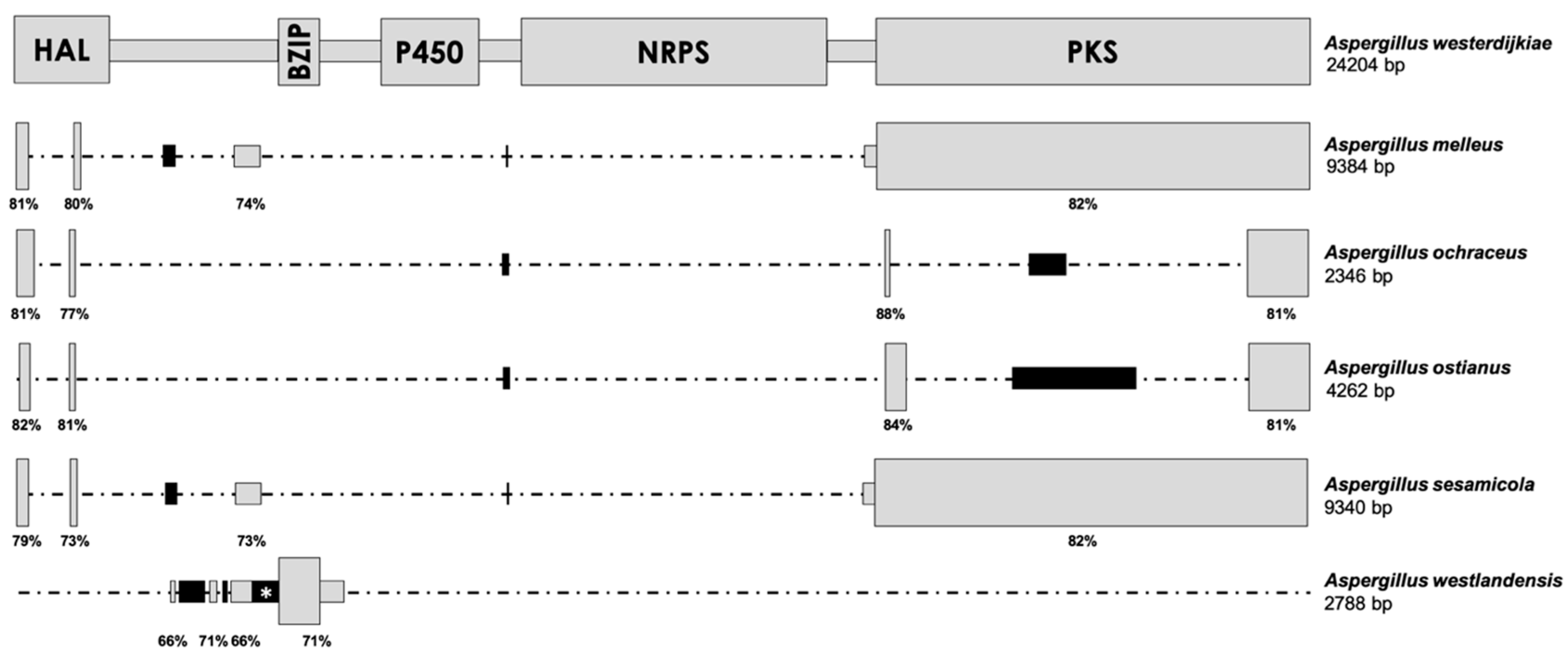
Figure 2.
Comparison of the OTA biosynthetic cluster in A. steynii (above) with the corresponding regions found in A. persii and A. sclerotiorum. The schemes are drawn at scale, except in the cases of black regions marked by an asterisk (*), which are slightly longer than represented. The length of the complete region in fungal genomes is indicated below the corresponding species name. Gray regions correspond to sequences that share high identity with the OTA biosynthetic cluster and are located under the corresponding homologous region in A. steynii. Black regions show no identity with any sequence available in databases. The striped square represents an inverted region. The percentages below the boxes indicate the region’s identity with A. steynii. HAL = halogenase-, BZIP = bZIP transcription factor-, P450 = cytochrome P450 monooxygenase-, NRPS = non-ribosomal peptide synthetase-, and PKS = polyketide synthase-encoding genes.
Figure 2.
Comparison of the OTA biosynthetic cluster in A. steynii (above) with the corresponding regions found in A. persii and A. sclerotiorum. The schemes are drawn at scale, except in the cases of black regions marked by an asterisk (*), which are slightly longer than represented. The length of the complete region in fungal genomes is indicated below the corresponding species name. Gray regions correspond to sequences that share high identity with the OTA biosynthetic cluster and are located under the corresponding homologous region in A. steynii. Black regions show no identity with any sequence available in databases. The striped square represents an inverted region. The percentages below the boxes indicate the region’s identity with A. steynii. HAL = halogenase-, BZIP = bZIP transcription factor-, P450 = cytochrome P450 monooxygenase-, NRPS = non-ribosomal peptide synthetase-, and PKS = polyketide synthase-encoding genes.
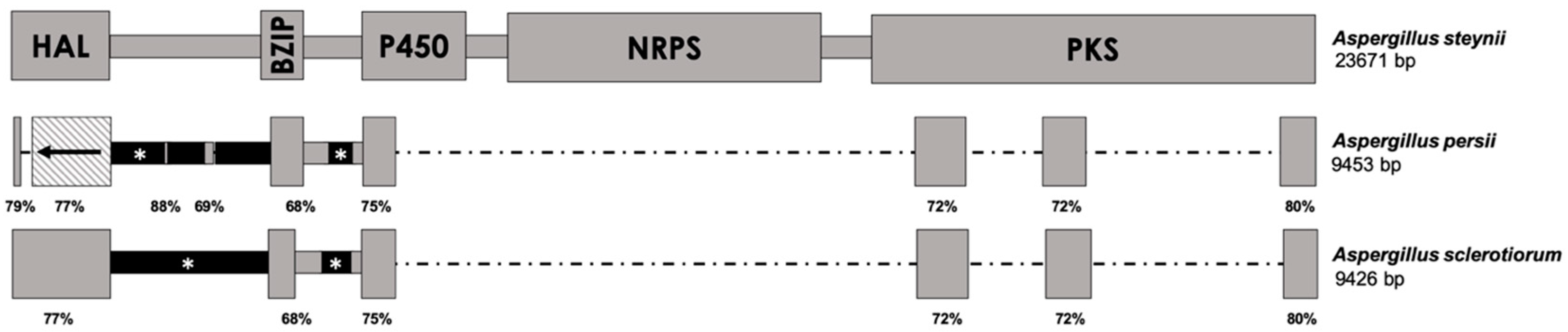
Figure 3.
Maximum likelihood tree derived from a combination of β-tubulin, ITS1-5.8S-ITS2, and calmodulin data. The tree with the highest log-likelihood (−5103.86) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches. A scheme (drawn at scale) of the putative region that includes the biosynthetic cluster in the genome of each taxon is located close to the corresponding branch.
Figure 3.
Maximum likelihood tree derived from a combination of β-tubulin, ITS1-5.8S-ITS2, and calmodulin data. The tree with the highest log-likelihood (−5103.86) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches. A scheme (drawn at scale) of the putative region that includes the biosynthetic cluster in the genome of each taxon is located close to the corresponding branch.
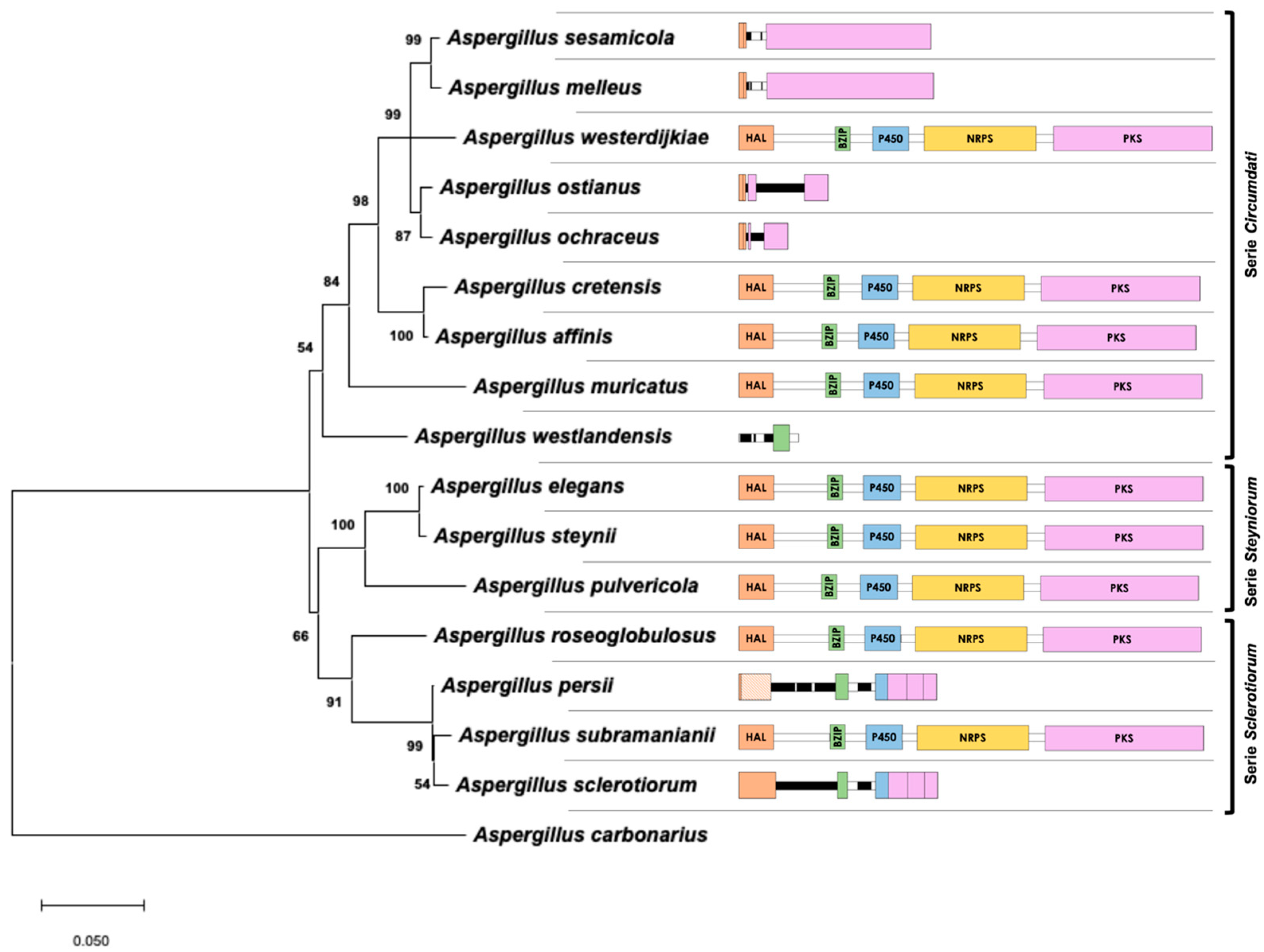
Figure 4.
Maximum likelihood tree derived from the 3′-end of the PKS-encoding gene sequences. The tree with the highest log likelihood (−3842.76) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches.
Figure 4.
Maximum likelihood tree derived from the 3′-end of the PKS-encoding gene sequences. The tree with the highest log likelihood (−3842.76) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches.
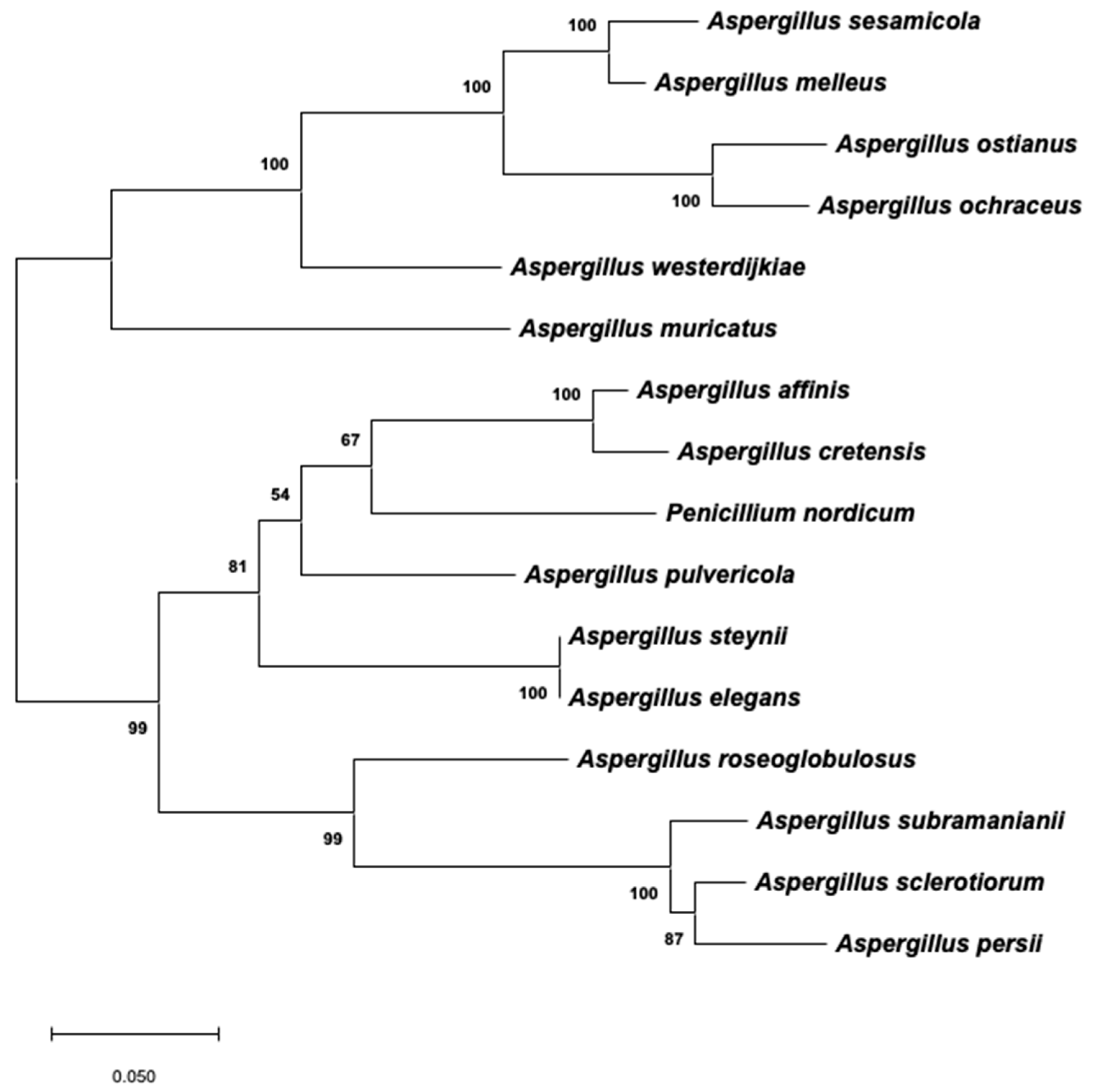
Figure 5.
Maximum likelihood tree derived from the sequences of the complete OTA biosynthetic cluster. The tree with the highest log-likelihood (−5103.86) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches.
Figure 5.
Maximum likelihood tree derived from the sequences of the complete OTA biosynthetic cluster. The tree with the highest log-likelihood (−5103.86) is shown. Branch lengths are measured in the number of base substitutions per site. Bootstrap values are shown next to the branches.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the ochratoxin A (OTA) biosynthetic clusters described in this work. The length of the complete cluster, from the beginning of the halogenase (HAL)-encoding gene to the end of the polyketide synthase (PKS)-encoding gene, is indicated, together with the accession number of the sequence in the NCBI database. The identity of the complete region compared to the reference clusters in A. steynii and A. westerdijkiae is also indicated.
Table 1.
Characteristics of the ochratoxin A (OTA) biosynthetic clusters described in this work. The length of the complete cluster, from the beginning of the halogenase (HAL)-encoding gene to the end of the polyketide synthase (PKS)-encoding gene, is indicated, together with the accession number of the sequence in the NCBI database. The identity of the complete region compared to the reference clusters in A. steynii and A. westerdijkiae is also indicated.
| Species | Cluster Length | Accession Number | Identity with OTA Cluster | |
|---|---|---|---|---|
| A. steynii | A. westerdijkiae | |||
| A. affinis | 23,387 | MT955635 | 75% | 67% |
| A. cretensis | 23,586 | MT955636 | 73% | 66% |
| A. elegans | 23,680 | MT904985 | 99% | 67% |
| A. muricatus | 23,696 | MT955638 | 70% | 70% |
| A. pulvericola | 23,409 | MT904986 | 74% | 66% |
| A. roseoglobulosus | 23,599 | MT955637 | 70% | 67% |
| A. subramanianii | 23,730 | MT955639 | 68% | 65% |
Table 2.
Accession numbers in the NCBI database of the sequences used to generate the combined dataset β-tubulin/ITS1-5.8S-ITS2/calmodulin for maximum likelihood phylogenetic analysis.
Table 2.
Accession numbers in the NCBI database of the sequences used to generate the combined dataset β-tubulin/ITS1-5.8S-ITS2/calmodulin for maximum likelihood phylogenetic analysis.
| Species | β-Tubulin | ITS1-5.8S-ITS2 | Calmodulin |
|---|---|---|---|
| Aspergillus affininis CBS 129190 | GU721092 | JN882309 | GU721091 |
| Aspergillus cretensis CBS 112802 | AY819977 | FJ491572 | FJ491534 |
| Aspergillus elegans CBS 102.14 | EF661349 | EF661414 | EF661390 |
| Aspergillus melleus CBS 546.65 | EF661326 | EF661425 | EF661391 |
| Aspergillus muricatus CBS 112808 | EF661356 | EF661434 | EF661377 |
| Aspergillus ochraceus CECT 2093 | EF661322 | EF661419 | EF661381 |
| Aspergillus ostianus CBS 103.07 | EF661324 | EF661421 | EF661385 |
| Aspergillus persii CBS 112795 | AY819988 | FJ491580 | FJ491559 |
| Aspergillus pulvericola CBS 137327 | KJ775055 | KJ775440 | KJ775236 |
| Aspergillus roseoglobulosus CBS 112800 | AY819984 | FJ491583 | FJ491555 |
| Aspergillus sclerotiorum CBS 549.65 | EF661337 | EF661400 | EF661384 |
| Aspergillus sesamicola CBS 137324 | KJ775063 | KJ775437 | KJ775233 |
| Aspergillus steynii CBS 112.812 | EF661347 | EF661416 | EF661378 |
| Aspergillus subramanianii CBS 138230 | EF661339 | EF661403 | EF661397 |
| Aspergillus westerdijkiae CECT 2948 | EF661329 | EF661427 | EF661360 |
| Aspergillus westlandensis CBS 123905 | KJ775065 | KJ775433 | KJ775229 |
| Aspergillus carbonarius NRRL 4849 | EF661100 | EF661205 | EF661168 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gil-Serna, J.; Vázquez, C.; Patiño, B. The Genomic Regions That Contain Ochratoxin A Biosynthetic Genes Widely Differ in Aspergillus Section Circumdati Species. Toxins 2020, 12, 754. https://doi.org/10.3390/toxins12120754
AMA Style
Gil-Serna J, Vázquez C, Patiño B. The Genomic Regions That Contain Ochratoxin A Biosynthetic Genes Widely Differ in Aspergillus Section Circumdati Species. Toxins. 2020; 12(12):754. https://doi.org/10.3390/toxins12120754
Chicago/Turabian StyleGil-Serna, Jéssica, Covadonga Vázquez, and Belén Patiño. 2020. "The Genomic Regions That Contain Ochratoxin A Biosynthetic Genes Widely Differ in Aspergillus Section Circumdati Species" Toxins 12, no. 12: 754. https://doi.org/10.3390/toxins12120754
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.