The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review

Abstract
1. Introduction
2. Structural Characterization
2.1. Cyclic Peptides
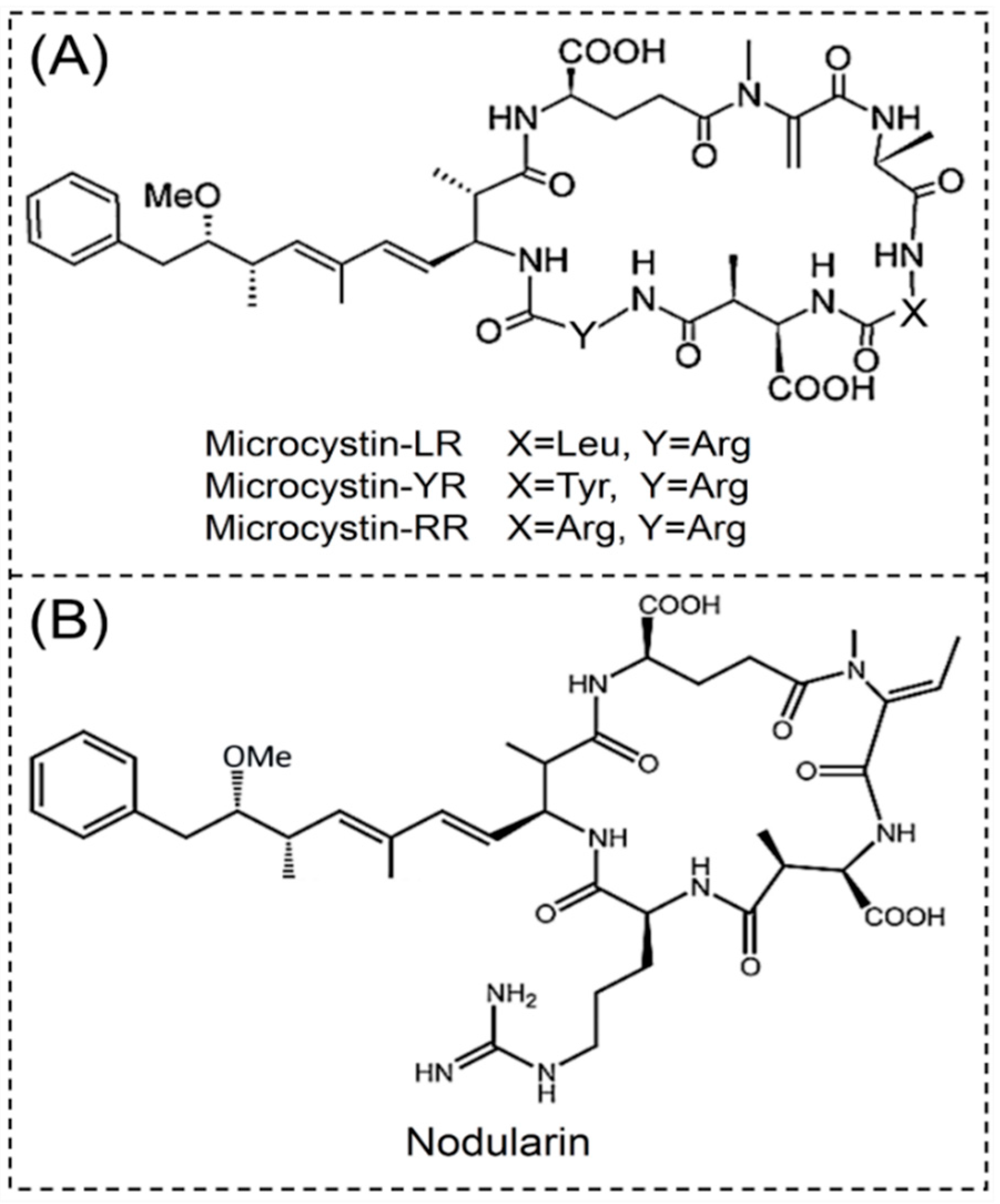
2.1.1. Microcystins
2.1.2. Nodularins
2.2. Alkaloids
2.2.1. Cylindrospermopsins
2.2.2. Saxitoxins
2.2.3. Anatoxin-a
2.2.4. Anatoxin-a(s)
2.2.5. Lyngbyatoxins
2.2.6. Aplysiatoxins
2.3. Lipopeptides
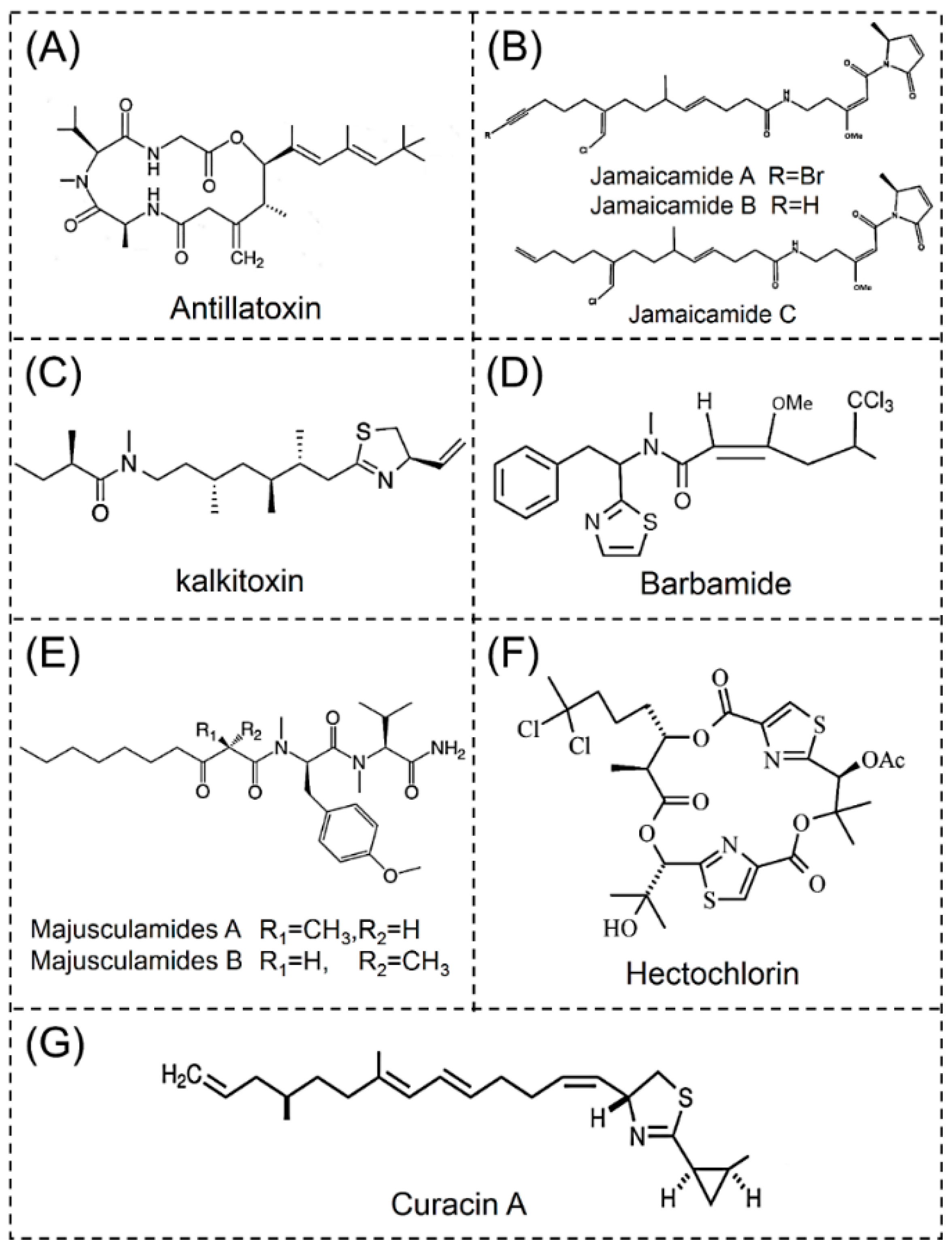
2.3.1. Antillatoxins
2.3.2. Jamaicamides
2.3.3. Kalkitoxins
2.3.4. Barbamides
2.3.5. Majusculamides
2.3.6. Hectochlorins
2.3.7. Curacins
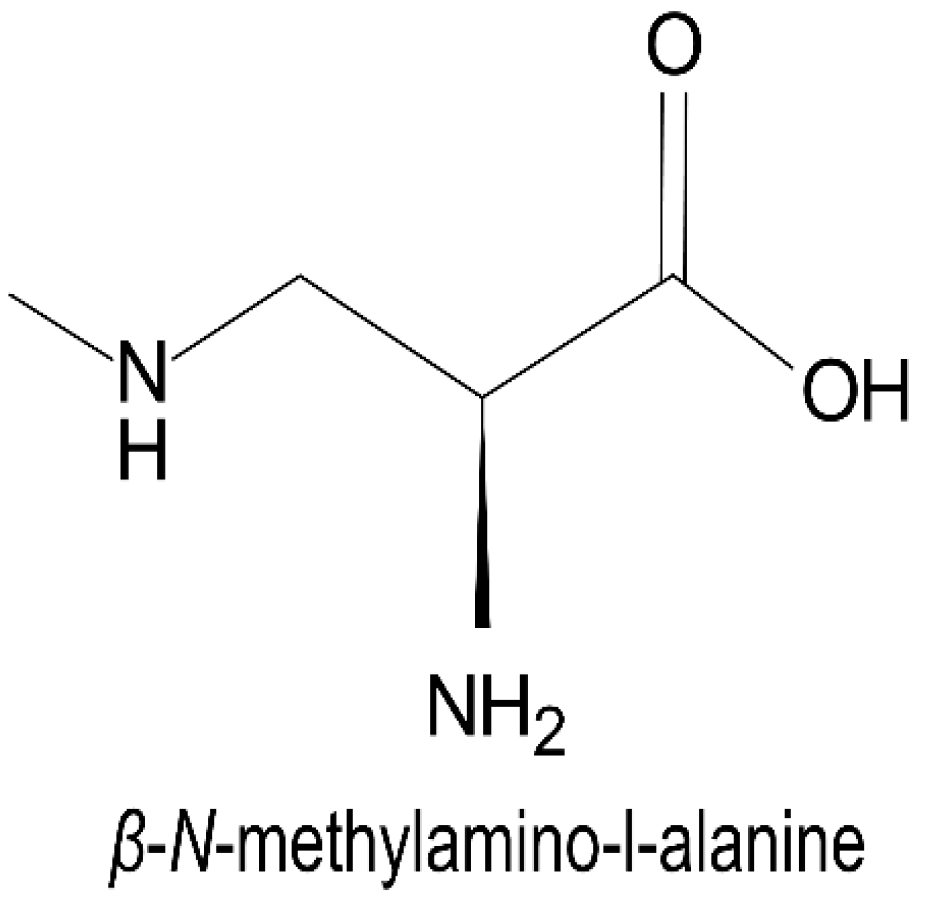
2.4. Nonprotein Amino Acids
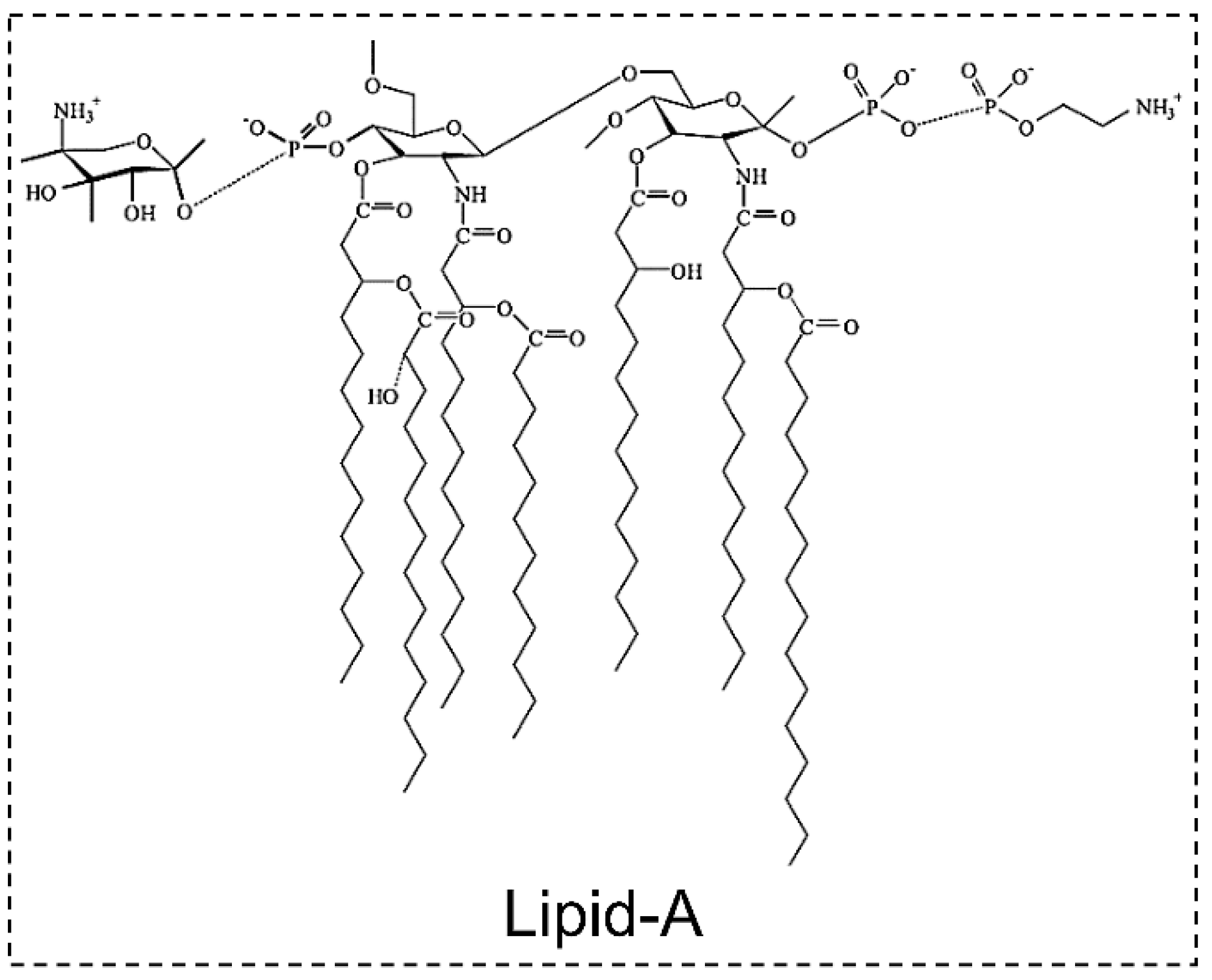
2.5. Lipoglycans
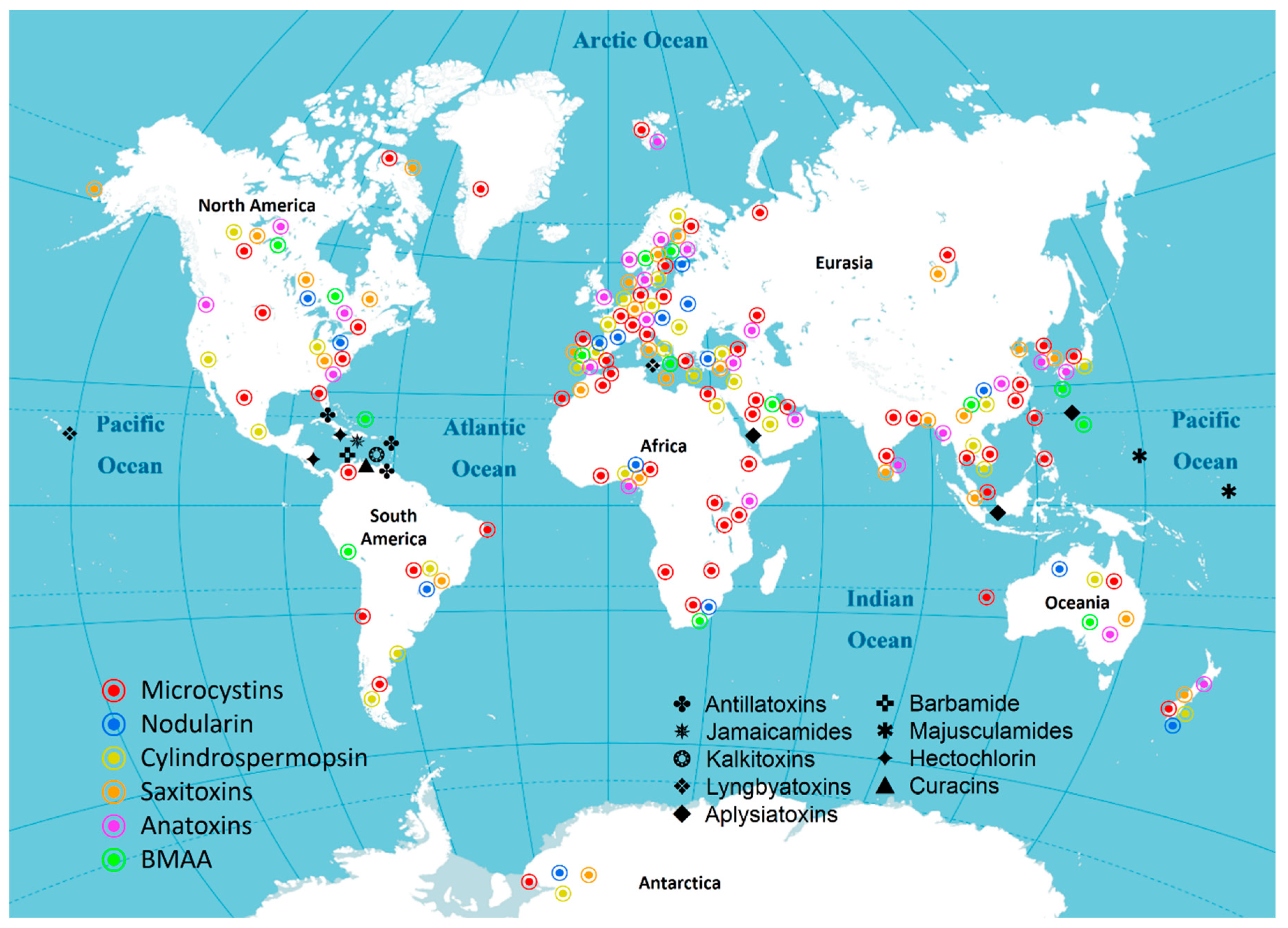
3. Distribution of Cyanotoxins
3.1. Asia
3.2. Africa
3.3. North America
3.4. South America
3.5. Antarctica
3.6. Europe
3.7. Oceania
3.8. The oceans
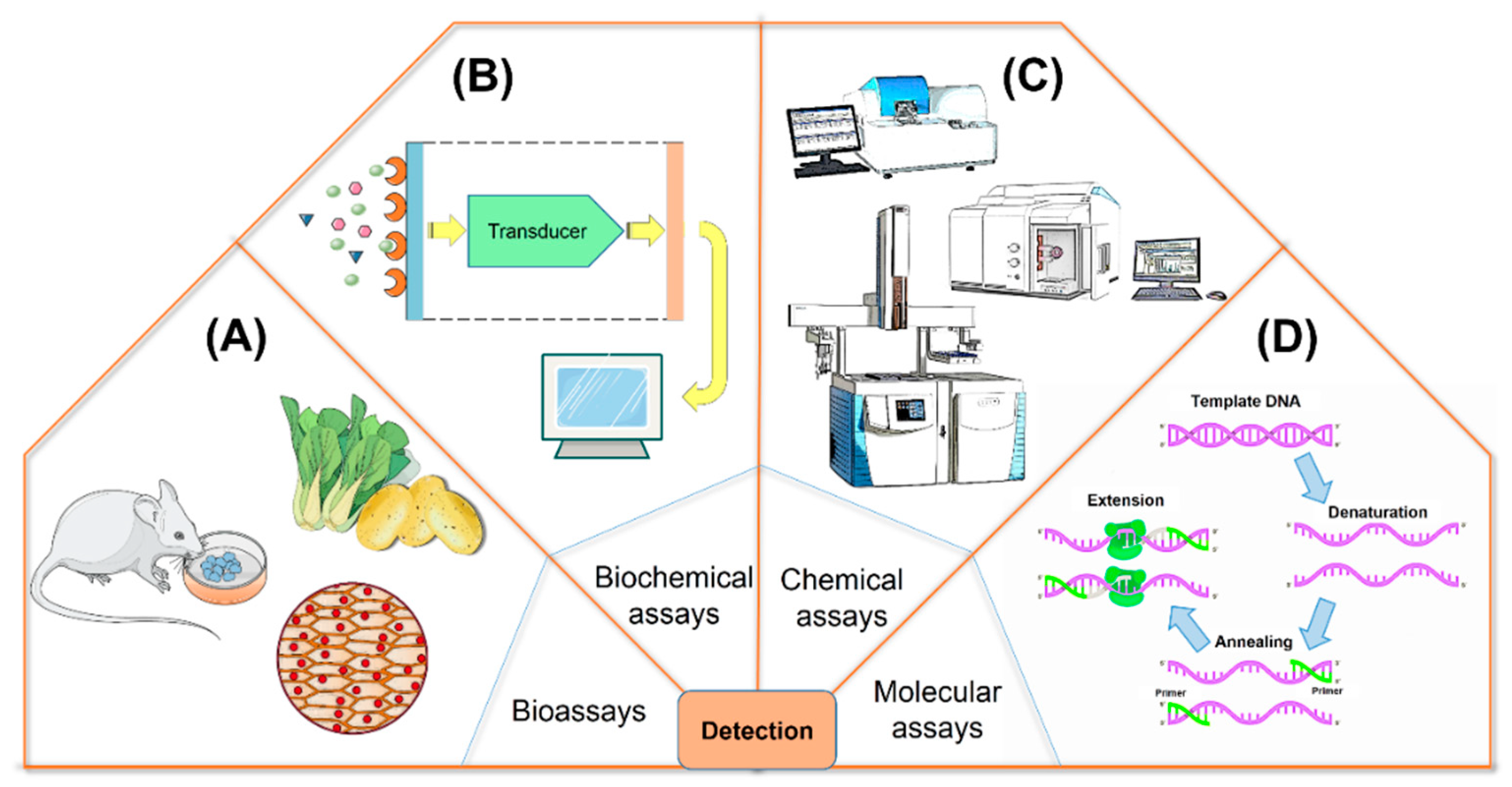
4. Identification of Cyanotoxins
4.1. Bioassays
4.2. Biochemical Assays
4.2.1. Immunoassays
4.2.2. Enzyme Inhibition Assays
4.2.3. Receptor Bioassays
4.3. Chemical Assays
4.4. Molecular Assays
5. Final Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Tokodi, N.; Drobac, D.; Lazić, G.; Petrović, T.; Marinović, Z.; Lujić, J.; Malešević, T.P.; Meriluoto, J.; Svirčev, Z. Screening of cyanobacterial cultures originating from different environments for cyanotoxicity and cyanotoxins. Toxicon 2018, 154, 1–6. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaicha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef]
- Giordano, D.; Costantini, M.; Coppola, D.; Lauritano, C.; Nunez Pons, L.; Ruocco, N.; di Prisco, G.; Ianora, A.; Verde, C. Biotechnological applications of bioactive peptides from marine sources. Adv. Microb. Physiol. 2018, 73, 171–220. [Google Scholar] [CrossRef] [PubMed]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [PubMed]
- Ruibal-Conti, A.L.; Ruiz, M.A.; Rodriguez, M.I.; Lerda, D.; Romero, M.D. Assessment of specific antibodies as biological indicators of human chronic exposure to microcystins. Ecotoxicol. Environ. Saf. 2019, 175, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Casero, M.C.; Quesada, A. Toxicity at the edge of life: A review on cyanobacterial toxins from extreme environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and nodularins. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 11, 107–126. [Google Scholar] [CrossRef]
- Harada, K.; Ogawa, K.; Matsuura, K.; Murata, H.; Suzuki, M.; Watanabe, M.F.; Itezono, Y.; Nakayama, N. Structural determination of geometrical isomers of microcystins LR and RR from cyanobacteria by two-dimensional NMR spectroscopic techniques. Chem. Res. Toxicol. 1990, 3, 473–481. [Google Scholar] [CrossRef]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Appendix 3: Tables of Microcystins and Nodularins. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 526–537. [Google Scholar] [CrossRef]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Humans, I. IARC monographs on the evaluation of carcinogenic risks to humans. J. Clin. Pathol. 2010, 94, v–vii, 1. [Google Scholar] [CrossRef]
- Salmaso, N.; Bernard, C.; Humbert, J.F.; Akçaalan, R.; Albay, M.; Ballot, A.; Catherine, A.; Fastner, J.; Häggqvist, K.; Horecká, M.; et al. Basic guide to detection and monitoring of potentially toxic cyanobacteria. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 6, 46–69. [Google Scholar] [CrossRef]
- Tamele, I.J.; Silva, M.; Vasconcelos, V. The incidence of marine toxins and the associated seafood poisoning episodes in the African countries of the Indian Ocean and the Red Sea. Toxins 2019, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Sangolkar, L.N.; Maske, S.S.; Chakrabarti, T. Methods for determining microcystins (peptide hepatotoxins) and microcystin-producing cyanobacteria. Water Res. 2006, 40, 3485–3496. [Google Scholar] [CrossRef]
- Shen, F.; Xu, Y.; Jiang, S.; Zhao, B.; Shi, J.; Chen, J. Determination of microcystins in algal bloom samples by ultra performance liquid chromatography-tandem mass spectrometry and elimination of their matrix effects. Chin. J. Chromatogr. (Se Pu) 2017, 35, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Waack, J.; Lewis, A.; Edwards, C.; Lawton, L. Development and single-laboratory validation of a UHPLC-MS/MS method for quantitation of microcystins and nodularin in natural water, cyanobacteria, shellfish and algal supplement tablet powders. J. Chromatogr. B 2018, 1074–1075, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shen, D.; Fang, D. Nodularins in poisoning. Clin. Chim. Acta 2013, 425, 18–29. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen nov., sp nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef]
- Shishido, T.K.; Jokela, J.; Humisto, A.; Suurnäkki, S.; Wahlsten, M.; Alvarenga, D.O.; Sivonen, K.; Fewer, D.P. The biosynthesis of rare homo-amino acid containing variants of microcystin by a benthic cyanobacterium. Mar. Drugs 2019, 17, 271. [Google Scholar] [CrossRef]
- Valério, E.; Chaves, S.; Tenreiro, R. Diversity and impact of prokaryotic toxins on aquatic environments: A review. Toxins 2010, 2, 2359–2410. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt-Oliveira, M.d.C.; Piccin, V.; Kujbida, P.; Moura, A. Cylindrospermopsin in water supply reservoirs in Brazil determined by immunochemical and molecular methods. J. Water Resour. Prot. 2011, 3, 349–355. [Google Scholar] [CrossRef]
- Kokocinski, M.; Maria Cameán, A.; Carmeli, S.; Guzmán-Guillén, R.; Jos, A.; Mankiewicz-Boczek, J.; Metcalf, J.; Moreno, I.; Isabel Prieto, A.; Sukenik, A. Cylindrospermopsin and congeners. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 12, 127–137. [Google Scholar] [CrossRef]
- Batsalova, T.; Basheva, D.; Bardarov, K.; Bardarov, V.; Dzhambazov, B.; Teneva, I. Assessment of the cytotoxicity, antioxidant activity and chemical composition of extracts from the cyanobacterium Fischerella major Gomont. Chemosphere 2019, 218, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Belykh, O.I.; Tikhonova, I.V.; Kuzmin, A.V.; Sorokovikova, E.G.; Fedorova, G.A.; Khanaev, I.V.; Sherbakova, T.A.; Timoshkin, O.A. First detection of benthic cyanobacteria in Lake Baikal producing paralytic shellfish toxins. Toxicon 2016, 121, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Murakami, T.; Watanabe, M.F.; Oda, T.; Yamada, J.; Tsujimura, S.; Nagai, H.; Oishi, S. Simultaneous production of homoanatoxin-a, anatoxin-a, and a new non-toxic 4-hydroxyhomoanatoxin-a by the cyanobacterium Raphidiopsis mediterranea Skuja. Toxicon 2003, 42, 533–538. [Google Scholar] [CrossRef]
- Wood, S.A.; Biessy, L.; Puddick, J. Anatoxins are consistently released into the water of streams with Microcoleus autumnalis-dominated (cyanobacteria) proliferations. Harmful Algae 2018, 80, 88–95. [Google Scholar] [CrossRef]
- Tan, L.T.; Goh, B.P.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.; Teo, S.L. Natural antifoulants from the marine cyanobacterium Lyngbya majuscula. Biofouling 2010, 26, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Cullen, A.; Pearson, L.A.; Mazmouz, R.; Liu, T.; Soeriyadi, A.H.; Ongley, S.E.; Neilan, B.A. Heterologous expression and biochemical characterisation of cyanotoxin biosynthesis pathways. Nat. Prod. Rep. 2018, 36, 1117–1136. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.; Shaala, L.A.; Mohamed, G.A.; Ibrahim, S.R.; Banjar, Z.M.; Badr, J.M.; McPhail, K.L.; Risinger, A.L.; Mooberry, S.L. 2,3-seco-2,3-dioxo-lyngbyatoxin A from a Red Sea strain of the marine cyanobacterium Moorea producens. Nat. Prod. Res. 2015, 29, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Aimi, N.; Odaka, H.; Sakai, S.I.; Fujiki, H.; Suganuma, M.; Moore, R.E.; Patterson, G.M.L. Lyngbyatoxins B and C, two new irritants from Lyngbya majuscula. J. Nat. Prod. 1990, 53, 1593–1596. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.Y.G.; Pang, L.M.; Liang, Z.X.; Goh, K.K.K.; Glukhov, E.; Gerwick, W.H.; Tan, L.T. MS/MS-based molecular networking approach for the detection of aplysiatoxin-related compounds in environmental marine cyanobacteria. Mar. Drugs 2018, 16, 505. [Google Scholar] [CrossRef] [PubMed]
- Nogle, L.M.; Okino, T.; Gerwick, W.H. Antillatoxin B, a neurotoxic lipopeptide from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2001, 64, 983–985. [Google Scholar] [CrossRef]
- Orjala, J.; Nagle, D.G.; Hsu, V.; Gerwick, W.H. Antillatoxin: An exceptionally ichthyotoxic cyclic lipopeptide from the tropical cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 1995, 117, 8281–8282. [Google Scholar] [CrossRef]
- Boudreau, P.D.; Monroe, E.A.; Mehrotra, S.; Desfor, S.; Korobeynikov, A.; Sherman, D.H.; Murray, T.F.; Gerwick, L.; Dorrestein, P.C.; Gerwick, W.H. Expanding the described metabolome of the marine cyanobacterium Moorea producens jhb through orthogonal natural products workflows. PLoS ONE 2015, 10, e0133297. [Google Scholar] [CrossRef]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and biosynthesis of the jamaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscula. Chem. Biol. 2004, 11, 817–833. [Google Scholar] [CrossRef]
- Umezawa, T.; Sueda, M.; Kamura, T.; Kawahara, T.; Han, X.; Okino, T.; Matsuda, F. Synthesis and biological activity of kalkitoxin and its analogues. J. Org. Chem. 2012, 77, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Okino, T.; Nogle, L.M.; Marquez, B.L.; Williamson, R.T.; Sitachitta, N.; Berman, F.W.; Murray, T.F.; McGough, K.; Jacobs, R.; et al. Structure, synthesis, and biological properties of kalkitoxin, a novel neurotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 2000, 122, 12041–12042. [Google Scholar] [CrossRef]
- Kothari, A.; Vaughn, M.; Garciapichel, F. Comparative genomic analyses of the cyanobacterium, Lyngbya aestuarii BL J, a powerful hydrogen producer. Front. Microbiol. 2013, 4, 363. [Google Scholar] [CrossRef] [PubMed]
- Caro-Diaz, E.J.E.; Valeriote, F.A.; Gerwick, W.H. Highly convergent total synthesis and assignment of absolute configuration of majusculamide D, a potent and selective cytotoxic metabolite from Moorea sp. Org. Lett. 2019, 21, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.C.; Moore, R.E.; Mynderse, J.S.; Niemczura, W.P.; Todd, J.S. Structure of majusculamide C, a cyclic depsipeptide from Lyngbya majuscula. J. Org. Chem. 1984, 49. [Google Scholar] [CrossRef]
- Suntornchashwej, S.; Chaichit, N.; Isobe, M.; Suwanborirux, K. Hectochlorin and Morpholine Derivatives from the Thai Sea Hare, Bursatella leachii. J. Nat. Prod. 2005, 68, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, A.V.; Sorrels, C.M.; Gerwick, W.H. Cloning and biochemical characterization of the hectochlorin biosynthetic gene cluster from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Burja, A.M.; Abou-Mansour, E.; Banaigs, B.; Payri, C.; Burgess, J.G.; Wright, P.C. Culture of the marine cyanobacterium, Lyngbya majuscula (Oscillatoriaceae), for bioprocess intensified production of cyclic and linear lipopeptides. J. Microbiol. Methods 2002, 48, 207–219. [Google Scholar] [CrossRef]
- Yoo, H.D.; Gerwick, W.H. Curacins B and C, new antimitotic natural products from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1995, 58, 1961–1965. [Google Scholar] [CrossRef]
- Zhang, C.; Naman, C.B.; Engene, N.; Gerwick, W.H. Laucysteinamide A, a hybrid PKS/NRPS metabolite from a Saipan Cyanobacterium, cf. Caldora penicillata. Mar. Drugs 2017, 15, 121. [Google Scholar] [CrossRef]
- Márquez, B.; Verdier-Pinard, P.; Hamel, E.; Gerwick, W.H. Curacin D, an antimitotic agent from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 1998, 49, 2387–2389. [Google Scholar] [CrossRef]
- Rodgers, K.J.; Main, B.J.; Samardzic, K. Cyanobacterial neurotoxins: Their occurrence and mechanisms of toxicity. Neurotox. Res. 2018, 33, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Murch, S.J. Methods for the chemical analysis of β-N-Methylamino-L-A lanine: What is known and what remains to be determined. Neurotox. Res. 2018, 33, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.; Costa, M.; Moreira, C.; Vasconcelos, V.M.; Baptista, M.S. Screening of BMAA-producing cyanobacteria in cultured isolates and in in situ blooms. J. Appl. Phycol. 2017, 29, 879–888. [Google Scholar] [CrossRef]
- Lage, S.; Burian, A.; Rasmussen, U.; Costa, P.R.; Annadotter, H.; Godhe, A.; Rydberg, S. BMAA extraction of cyanobacteria samples: Which method to choose? Environ. Sci. Pollut. Res. Int. 2016, 23, 338–350. [Google Scholar] [CrossRef]
- Manolidi, K.; Triantis, T.M.; Kaloudis, T.; Hiskia, A. Neurotoxin BMAA and its isomeric amino acids in cyanobacteria and cyanobacteria-based food supplements. J. Hazard. Mater. 2019, 365, 346–365. [Google Scholar] [CrossRef]
- Kehr, J.C.; Dittmann, E. Biosynthesis and function of extracellular glycans in cyanobacteria. Life (Basel) 2015, 5, 164–180. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Gutierrez-Praena, D.; Prieto, A.I.; Guzman-Guillen, R.; Jos, A.; Camean, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef]
- Ohtani, I.; Moore, R.E.; Runnegar, M. Cylindrospermopsin: A potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. ChemInform 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Cameán, A.M.; Jos, A. Neurotoxic assessment of Microcystin-LR, cylindrospermopsin and their combination on the human neuroblastoma SH-SY5Y cell line. Chemosphere 2019, 224, 751–764. [Google Scholar] [CrossRef]
- Bourke, A.T.C.; Hawes, R.B.; Neilson, A.; Stallman, N.D. An outbreak of hepato-enteritis (the Palm Island mystery disease) possibly caused by algal intoxication. Toxicon 1983, 21, 45–48. [Google Scholar] [CrossRef]
- Harada, K.I.; Ohtani, I.; Iwamoto, K.; Suzuki, M.; Watanabe, M.F.; Watanabe, M.; Terao, K. Isolation of cylindrospermopsin from a cyanobacterium Umezakia natans and its screening method. Toxicon 1994, 32, 73–84. [Google Scholar] [CrossRef]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2013, 114, 605–620. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Prieto, A.I.; González, A.G.; Soria-Díaz, M.E.; Cameán, A.M. Cylindrospermopsin determination in water by LC-MS/MS: Optimization and validation of the method and application to real samples. Environ. Toxicol. Chem. 2012, 31, 2233–2238. [Google Scholar] [CrossRef]
- Wong, J.L.; Oesterlin, R.; Rapoport, H. The structure of saxitoxin. J. Am. Chem. Soc. 1971, 93, 7344–7345. [Google Scholar] [CrossRef]
- Ember, L. Chemical weapons convention. Chem. Eng. News Arch. 1998, 76, 9. [Google Scholar] [CrossRef]
- Hallegraeff, G. Harmful algal blooms: A gobal overview. Camb. Law J. 1995, 9, 268–270. [Google Scholar] [CrossRef]
- Humpage, A.R.; Magalhaes, V.F.; Froscio, S.M. Comparison of analytical tools and biological assays for detection of paralytic shellfish poisoning toxins. Anal. Bioanal. Chem. 2010, 397, 1655–1671. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef]
- Haddad, S.P.; Bobbitt, J.M.; Taylor, R.B.; Lovin, L.M.; Conkle, J.L.; Chambliss, C.K.; Brooks, B.W. Determination of microcystins, nodularin, anatoxin-a, cylindrospermopsin, and saxitoxin in water and fish tissue using isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019. [Google Scholar] [CrossRef]
- Bragg, W.A.; Lemire, S.W.; Coleman, R.M.; Hamelin, E.I.; Johnson, R.C. Detection of human exposure to saxitoxin and neosaxitoxin in urine by online-solid phase extraction-liquid chromatography-tandem mass spectrometry. Toxicon 2015, 99, 118–124. [Google Scholar] [CrossRef]
- Devlin, J.P.; Edwards, O.E.; Gorham, P.R.; Hunter, N.R.; Pike, R.K.; Stavric, B. Anatoxin-a, a toxic alkaloid from Anabaena flos-aquae NRC-44h. Can. J. Chem. 1977, 55, 1367–1371. [Google Scholar] [CrossRef]
- Wonnacott, S.; Swanson, K.L.; Albuquerque, E.X.; Huby, N.J.S.; Thompson, P.; Gallagher, T. Homoanatoxin: A potent analogue of anatoxin-a. Biochem. Pharmacol. 1992, 43, 419–423. [Google Scholar] [CrossRef]
- Bruno, M.; Ploux, O.; Metcalf, J.; Méjean, A.; Pawlik-Skowrońska, B.; Furey, A. Anatoxin-a, homoanatoxin-a, and natural analogues. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 13, 138–147. [Google Scholar] [CrossRef]
- Osswald, J.; Rellán, S.; Gago, A.; Vasconcelos, V. Toxicology and detection methods of the alkaloid neurotoxin produced by cyanobacteria, anatoxin-a. Environ. Int. 2007, 33, 1070–1089. [Google Scholar] [CrossRef]
- Legrand, B.; Lesobre, J.; Colombet, J.; Latour, D.; Sabart, M. Molecular tools to detect anatoxin-a genes in aquatic ecosystems: Toward a new nested PCR-based method. Harmful Algae 2016, 58, 16–22. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Matsunaga, S.; Moore, R.E.; Niemczura, W.P.; Carmichael, W.W. Anatoxin-a(s), a potent anticholinesterase from Anabaena flos-aquae. J. Am. Chem. Soc. 1989, 111, 8021–8023. [Google Scholar] [CrossRef]
- Mahmood, N.A.; Carmichael, W.W. The pharmacology of anatoxin-a(s), a neurotoxin produced by the freshwater cyanobacterium Anabaena flos-aquae NRC 525-17. Toxicon 1986, 24, 425–434. [Google Scholar] [CrossRef]
- Marie, B.; Blaha, L.; Maria Cameán, A.; Fessard, V.; Gutiérrez-Praena, D.; Jos, A.; Metcalf, J.; Pichardo, S.; Puerto, M.; Törökné, A.; et al. Bioassay use in the field of toxic cyanobacteria. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2016, 27, 272–278. [Google Scholar] [CrossRef]
- Metcalf, J.; Bruno, M. Anatoxin-a(S). Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 15, 155–159. [Google Scholar] [CrossRef]
- Villatte, F.; Schulze, H.; Schmid, R.D.; Bachmann, T.T. A disposable acetylcholinesterase-based electrode biosensor to detect anatoxin-a(s) in water. Anal. Bioanal. Chem. 2002, 372, 322–326. [Google Scholar] [CrossRef]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Seaweed dermatitis: Structure of lyngbyatoxin A. Science 1979, 204, 193. [Google Scholar] [CrossRef]
- Fujiki, H.; Mori, M.; Nakayasu, M.; Terada, M.; Sugimura, T.; Moore, R.E. Indole alkaloids: Dihydroteleocidin B, teleocidin, and lyngbyatoxin A as members of a new class of tumor promoters. Proc. Natl. Acad. Sci. USA 1981, 78, 3872–3876. [Google Scholar] [CrossRef]
- Ito, E.; Satake, M.; Yasumoto, T. Pathological effects of lyngbyatoxin A upon mice. Toxicon 2002, 40, 551–556. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, W.; Uchida, H.; Kikumori, M.; Irie, K.; Watanabe, R.; Suzuki, T.; Sakamoto, B.; Kamio, M.; Nagai, H. A new lyngbyatoxin from the Hawaiian cyanobacterium Moorea producens. Mar. Drugs 2014, 12, 2748–2759. [Google Scholar] [CrossRef]
- Esposito, G.; Teta, R.; Marrone, R.; De Sterlich, C.; Casazza, M.; Anastasio, A.; Lega, M.; Costantino, V. A Fast Detection Strategy for Cyanobacterial blooms and associated cyanotoxins (FDSCC) reveals the occurrence of lyngbyatoxin A in campania (South Italy). Chemosphere 2019, 225, 342–351. [Google Scholar] [CrossRef]
- Kato, Y.; Scheuer, P.J. Aplysiatoxin and debromoaplysiatoxin, constituents of the marine mollusk Stylocheilus longicauda (Quoy and Gaimard, 1824). J. Am. Chem. Soc. 1974, 96, 2245–2246. [Google Scholar] [CrossRef]
- Fujiki, H.; Ikegami, K.; Hakii, H.; Suganuma, M.; Yamaizumi, Z.; Yamazato, K.; Moore, R.E.; Sugimura, T. A blue-green alga from Okinawa contains aplysiatoxins, the third class of tumor promoters. Jpn. J. Cancer Res. 1985, 76, 257–259. [Google Scholar]
- Richard, K.; Williams, D.E.; de Silva, E.D.; Brockman, M.A.; Brumme, Z.L.; Andersen, R.J.; Tietjen, I. Identification of novel HIV-1 latency-reversing agents from a library of marine natural products. Viruses 2018, 10, 348. [Google Scholar] [CrossRef]
- Li, W.I.; Marquez, B.L.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. Characterization of the preferred stereochemistry for the neuropharmacologic actions of antillatoxin. J. Nat. Prod. 2004, 67, 559–568. [Google Scholar] [CrossRef]
- Jabba, S.V.; Prakash, A.; Dravid, S.M.; Gerwick, W.H.; Murray, T.F. Antillatoxin, a novel lipopeptide, enhances neurite outgrowth in immature cerebrocortical neurons through activation of voltage-gated sodium channels. J. Pharmacol. Exp. Ther. 2010, 332, 698–709. [Google Scholar] [CrossRef]
- Okura, K.; Matsuoka, S.; Inoue, M. The bulky side chain of antillatoxin is important for potent toxicity: Rational design of photoresponsive cytotoxins based on SAR studies. Chem. Commun. 2013, 49, 8024–8026. [Google Scholar] [CrossRef]
- Graf, K.M.; Tabor, M.G.; Brown, M.L.; Paige, M. Synthesis of (S)-jamaicamide C carboxylic acid. Org. Lett. 2009, 11, 5382–5385. [Google Scholar] [CrossRef]
- Araoz, R.; Molgo, J.; de Marsac, N.T. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef]
- Morgan, J.B.; Liu, Y.; Coothankandaswamy, V.; Mahdi, F.; Jekabsons, M.B.; Gerwick, W.H.; Valeriote, F.A.; Zhou, Y.D.; Nagle, D.G. Kalkitoxin inhibits angiogenesis, disrupts cellular hypoxic signaling, and blocks mitochondrial electron transport in tumor cells. Mar. Drugs 2015, 13, 1552–1568. [Google Scholar] [CrossRef]
- Orjala, J.; Gerwick, W.H. Barbamide, a chlorinated metabolite with molluscicidal activity from the Caribbean Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1996, 59, 427–430. [Google Scholar] [CrossRef]
- Sitachitta, N.; Márquez, B.L.; Williamson, R.T.; Rossi, J.; Roberts, M.A.; Gerwick, W.H.; Nguyen, V.A.; Willis, C.L.J.T. Biosynthetic pathway and origin of the chlorinated methyl group in barbamide and dechlorobarbamide, metabolites from the marine cyanobacterium Lyngbya majuscula. Tetrahedron 2000, 56, 9103–9113. [Google Scholar] [CrossRef]
- Cetusic, J.R.P.; Green, F.R.; Graupner, P.R.; Oliver, M.P. Total synthesis of hectochlorin. Org. Lett. 2002, 4, 1307–1310. [Google Scholar] [CrossRef]
- Marquez, B.L.; Watts, K.S.; Yokochi, A.; Roberts, M.A.; Verdier-Pinard, P.; Jimenez, J.I.; Hamel, E.; Scheuer, P.J.; Gerwick, W.H. Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J. Nat. Prod. 2002, 65, 866–871. [Google Scholar] [CrossRef]
- Timmins, A.; Fowler, N.J.; Warwicker, J.; Straganz, G.D.; de Visser, S.P. Does substrate positioning affect the selectivity and reactivity in the hectochlorin biosynthesis halogenase? Front. Chem. 2018, 6, 513. [Google Scholar] [CrossRef] [PubMed]
- Ueoka, R.; Hitora, Y.; Ito, A.; Yoshida, M.; Okada, S.; Takada, K.; Matsunaga, S. Curacin E from the Brittle Star Ophiocoma scolopendrina. J. Nat. Prod. 2016, 79, 2754–2757. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Proteau, P.J.; Nagle, D.G.; Hamel, E.; Slate, D.L.J.C. Structure of curacin A, a novel antimitotic, antiproliferative and brine shrimp toxic natural product from the marine cyanobacterium Lyngbya majuscula. J. Organic Chem. 1994, 59. [Google Scholar] [CrossRef]
- Chang, Z.; Sitachitta, N.; Rossi, J.V.; Roberts, M.A.; Flatt, P.M.; Jia, J.; Sherman, D.H.; Gerwick, W.H. Biosynthetic pathway and gene cluster analysis of curacin A, an antitubulin natural product from the tropical marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2004, 67, 1356–1367. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.D.; Dodge, G.J.; Sherman, D.H.; Smith, J.L.; Aldrich, C.C. Vinylogous dehydration by a polyketide dehydratase domain in curacin biosynthesis. J. Am. Chem. Soc. 2016, 138, 16024–16036. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Bell, E.A. α-Amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Ploux, O.; Combes, A.; Eriksson, J.; Metcalf, J. β-N-methylamino-l-alanine and (S)-2,4-diaminobutyric acid. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 16, 160–164. [Google Scholar] [CrossRef]
- Main, B.J.; Bowling, L.C.; Padula, M.P.; Bishop, D.P.; Mitrovic, S.M.; Guillemin, G.J.; Rodgers, K.J. Detection of the suspected neurotoxin β-methylamino-l-alanine (BMAA) in cyanobacterial blooms from multiple water bodies in Eastern Australia. Harmful Algae 2018, 74, 10–18. [Google Scholar] [CrossRef]
- Cox, P.A.; Kostrzewa, R.M.; Guillemin, G.J. BMAA and neurodegenerative illness. Neurotox. Res. 2018, 33, 178–183. [Google Scholar] [CrossRef]
- Banack, S.A.; Cox, P.A. Biomagnification of cycad neurotoxins in flying foxes: Implications for ALS-PDC in Guam. Neurology 2003, 61, 387–389. [Google Scholar] [CrossRef]
- Porojan, C.; Mitrovic, S.M.; Yeo, D.C.; Furey, A. Overview of the potent cyanobacterial neurotoxin β-methylamino-l-alanine (BMAA) and its analytical determination. Food Addit. Contam. Part A 2016, 33, 1570–1586. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Bernard, C.; Fastner, J. Saxitoxin and analogues. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 14, 148–154. [Google Scholar] [CrossRef]
- Banack, S.A.; Johnson, H.E.; Cheng, R.; Cox, P.A. Production of the neurotoxin BMAA by a marine cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Kerrin, E.S.; White, R.L.; Quilliam, M.A. Quantitative determination of the neurotoxin beta-N-methylamino-l-alanine (BMAA) by capillary electrophoresis-tandem mass spectrometry. Anal. Bioanal. Chem. 2017, 409, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Durai, P.; Batool, M.; Choi, S. Structure and effects of cyanobacterial lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Lahti, K.; Rasanen, L.A.; Esala, A.L.; Niemela, S.I.; Sivonen, K. Endotoxins associated with cyanobacteria and their removal during drinking water treatment. Water Res. 2002, 36, 2627–2635. [Google Scholar] [CrossRef]
- Best, J.H.; Pflugmacher, S.; Wiegand, C.; Eddy, F.B.; Metcalf, J.S.; Codd, G.A. Effects of enteric bacterial and cyanobacterial lipopolysaccharides, and of microcystin-LR, on glutathione S-transferase activities in zebra fish (Danio rerio). Aquat. Toxicol. 2002, 60, 223–231. [Google Scholar] [CrossRef]
- Bláhová, L.; Blaha, L. Extraction, purification, and testing of LPS from cyanobacterial samples. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 56, 447–451. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, F.; Zhang, M.; Zhang, Z.; Bai, M.; Guo, G.; Zheng, W.; Wang, Q.; Shi, Y.; Wang, L. Endotoxin contamination, a potentially important inflammation factor in water and wastewater: A review. Sci. Total Environ. 2019, 681, 365–378. [Google Scholar] [CrossRef]
- Blahova, L.; Adamovsky, O.; Kubala, L.; Svihalkova Sindlerova, L.; Zounkova, R.; Blaha, L. The isolation and characterization of lipopolysaccharides from Microcystis aeruginosa, a prominent toxic water bloom forming cyanobacteria. Toxicon 2013, 76, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Reta, N.; Michelmore, A.; Saint, C.P.; Prieto-Simon, B.; Voelcker, N.H. Label-free bacterial toxin detection in water supplies using porous silicon nanochannel sensors. ACS Sens. 2019, 4, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wei, J.; Yang, K.; Massey, I.Y.; Guo, J.; Zhang, C.; Yang, F. Isolation, molecular identification, and characterization of a unique toxic cyanobacterium Microcystis sp. found in Hunan Province, China. J. Toxicol. Environ. Health Part A 2018, 81, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of microcystin-LR by a novel native effective bacterial community designated as YFMCD4 isolated from Lake Taihu. Toxins 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lou, I.; Ung, W.K.; Kong, Y.; Mok, K.M. Analysis of cylindrospermopsin- and microcystin-producing genotypes and cyanotoxin concentrations in the Macau storage reservoir. Hydrobiologia 2014, 741, 51–68. [Google Scholar] [CrossRef]
- Huang, W.J.; Lai, C.H.; Cheng, Y.L. Evaluation of extracellular products and mutagenicity in cyanobacteria cultures separated from a eutrophic reservoir. Sci. Total Environ. 2007, 377, 214–223. [Google Scholar] [CrossRef]
- Takahashi, T.; Umehara, A.; Tsutsumi, H. Diffusion of microcystins (cyanobacteria hepatotoxins) from the reservoir of Isahaya Bay, Japan, into the marine and surrounding ecosystems as a result of large-scale drainage. Mar. Pollut. Bull. 2014, 89, 250–258. [Google Scholar] [CrossRef]
- Joung, S.H.; Oh, H.M.; Ko, S.R.; Ahn, C.Y. Correlations between environmental factors and toxic and non-toxic Microcystis dynamics during bloom in Daechung Reservoir, Korea. Harmful Algae 2011, 10, 188–193. [Google Scholar] [CrossRef]
- Welker, M.; Khan, S.; Haque, M.M.; Islam, S.; Khan, N.H.; Chorus, I.; Fastner, J. Microcystins (cyanobacterial toxins) in surface waters of rural Bangladesh: Pilot study. J. Water Health 2005, 3, 325–337. [Google Scholar] [CrossRef]
- Shu Harn, T.; Gin, K. The dynamics of cyanobacteria and microcystin production in a tropical reservoir of Singapore. Harmful Algae 2010, 10, 1–44. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystins in groundwater wells and their accumulation in vegetable plants irrigated with contaminated waters in Saudi Arabia. J. Hazard. Mater. 2009, 172, 310–315. [Google Scholar] [CrossRef]
- Singh, S.; Asthana, R.K. Assessment of microcystin concentration in carp and catfish: A case study from Lakshmikund pond, Varanasi, India. Bull. Environ. Contam. Toxicol. 2014, 92, 687–692. [Google Scholar] [CrossRef]
- Cuvin-Aralar, M.L.; Fastner, J.; Focken, U.; Becker, K.; Aralar, E.V. Microcystins in natural blooms and laboratory cultured Microcystis aeruginosa from Laguna de Bay, Philippines. Syst. Appl. Microbiol. 2002, 25, 179–182. [Google Scholar] [CrossRef]
- Somdee, T.; Kaewsan, T.; Somdee, A. Monitoring toxic cyanobacteria and cyanotoxins (microcystins and cylindrospermopsins) in four recreational reservoirs (Khon Kaen, Thailand). Environ. Monit. Assess. 2013, 185, 9521–9529. [Google Scholar] [CrossRef]
- Dao, T.S.; Nimptsch, J.; Wiegand, C. Dynamics of cyanobacteria and cyanobacterial toxins and their correlation with environmental parameters in Tri An Reservoir, Vietnam. J. Water Health 2016, 14, 699–712. [Google Scholar] [CrossRef]
- Trung, B.; Dao, T.S.; Faassen, E.; Lurling, M. Cyanobacterial blooms and Microcystins in Southern Vietnam. Toxins 2018, 10, 471. [Google Scholar] [CrossRef]
- Lifshits, M.; Carmeli, S. Metabolites of Microcystis aeruginosa bloom material from Lake Kinneret, Israel. J. Nat. Prod. 2012, 75, 209–219. [Google Scholar] [CrossRef]
- Rodriguez, I.; Fraga, M.; Alfonso, A.; Guillebault, D.; Medlin, L.; Baudart, J.; Jacob, P.; Helmi, K.; Meyer, T.; Breitenbach, U.; et al. Monitoring of freshwater toxins in European environmental waters by using novel multi-detection methods. Environ. Toxicol. Chem. 2017, 36, 645–654. [Google Scholar] [CrossRef]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Liu, Y.; Watanabe, M.M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (Cyantobacteria). J. Phycol. 2001, 37, 1121–1126. [Google Scholar] [CrossRef]
- Chonudomkul, D.; Yongmanitchai, W.; Theeragool, G.; Kawachi, M.; Kasai, F.; Kaya, K.; Watanabe, M.M. Morphology, genetic diversity, temperature tolerance and toxicity of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) strains from Thailand and Japan. FEMS Microbiol. Ecol. 2004, 48, 345–355. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al-Shehri, A.M. Assessment of cylindrospermopsin toxin in an arid Saudi lake containing dense cyanobacterial bloom. Environ. Monit. Assess. 2013, 185, 2157–2166. [Google Scholar] [CrossRef]
- Dao, T.S.; Ortiz-Rodríguez, R.; Do-Hong, L.C.; Wiegand, C. Non-microcystin and non-cylindrospermopsin producing cyanobacteria affect the biochemical responses and behavior of Daphnia magna. Int. Rev. Hydrobiol. 2013, 98, 235–244. [Google Scholar] [CrossRef]
- Wimmer, K.M.; Strangman, W.K.; Wright, J.L.C. 7-Deoxy-desulfo-cylindrospermopsin and 7-deoxy-desulfo-12-acetylcylindrospermopsin: Two new cylindrospermopsin analogs isolated from a Thai strain of Cylindrospermopsis raciborskii. Harmful Algae 2014, 37, 203–206. [Google Scholar] [CrossRef]
- Ballot, A.; Ramm, J.; Rundberget, T.; Kaplan-Levy, R.N.; Hadas, O.; Sukenik, A.; Wiedner, C. Occurrence of non-cylindrospermopsin-producing Aphanizomenon ovalisporum and Anabaena bergii in Lake Kinneret (Israel). J. Plankton Res. 2011, 33, 1736–1746. [Google Scholar] [CrossRef]
- Akcaalan, R.; Koker, L.; Oguz, A.; Spoof, L.; Meriluoto, J.; Albay, M. First report of cylindrospermopsin production by two cyanobacteria (Dolichospermum mendotae and Chrysosporum ovalisporum) in Lake Iznik, Turkey. Toxins 2014, 6, 3173–3186. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, S.; Beier, R.C.; Beloglazova, N.V.; Lei, H.; Sun, X.; Ke, Y.; Zhang, S.; Wang, Z. Simple, high efficiency detection of microcystins and nodularin-R in water by fluorescence polarization immunoassay. Anal. Chim. Acta 2017, 992, 119–127. [Google Scholar] [CrossRef]
- Wu, H.Y.; Luan, Q.S.; Guo, M.M.; Gu, H.F.; Zhai, Y.X.; Tan, Z.J. Phycotoxins in scallops (Patinopecten yessoensis) in relation to source, composition and temporal variation of phytoplankton and cysts in North Yellow Sea, China. Mar. Pollut. Bull. 2018, 135, 1198–1204. [Google Scholar] [CrossRef]
- Shin, C.; Jo, H.; Kim, S.H.; Kang, G.J. Exposure assessment to paralytic shellfish toxins through the shellfish consumption in Korea. Food Res. Int. 2018, 108, 274–279. [Google Scholar] [CrossRef]
- Shunmugam, S.; Gayathri, M.; Prasannabalaji, N.; Thajuddin, N.; Muralitharan, G. Unraveling the presence of multi-class toxins from Trichodesmium bloom in the Gulf of Mannar region of the Bay of Bengal. Toxicon 2017, 135, 43–50. [Google Scholar] [CrossRef]
- James Holmes, M.; Bolch, C.; Green, D.; Cembella, A.; Lay Ming Teo, S. Singapore isolates of the dinoflagellate Gymnodinium catenatum (Dinophyceae) produce a unique profile of paralytic shellfish toxins. J. Phycol. 2002, 38, 96–106. [Google Scholar] [CrossRef]
- Zaman, L.; Arakawa, O.; Shimosu, A.; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 35, 423–431. [Google Scholar] [CrossRef]
- Yilmaz, M.; Foss, A.J.; Selwood, A.I.; Ozen, M.; Boundy, M. Paralytic shellfish toxin producing Aphanizomenon gracile strains isolated from Lake Iznik, Turkey. Toxicon 2018, 148, 132–142. [Google Scholar] [CrossRef]
- Srivastava, A.; Ahn, C.Y.; Asthana, R.K.; Lee, H.G.; Oh, H.M. Status, alert system, and prediction of cyanobacterial bloom in South Korea. Biomed. Res. Int. 2015, 2015, 584696. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Richer, R.; Cox, P.A.; Codd, G.A. Cyanotoxins in desert environments may present a risk to human health. Sci. Total Environ. 2012, 421–422, 118–123. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin β-methylamino-l-alanine within the aquatic food webs of Gonghu Bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468–469, 457–463. [Google Scholar] [CrossRef]
- Caparros-Lefebvre, D.; Steele, J.; Kotake, Y.; Ohta, S. Geographic isolates of atypical Parkinsonism and tauopathy in the tropics: Possible synergy of neurotoxins. Mov. Disord. 2006, 21, 1769–1771. [Google Scholar] [CrossRef]
- Craighead, D.; Metcalf, J.S.; Banack, S.A.; Amgalan, L.; Reynolds, H.V.; Batmunkh, M. Presence of the neurotoxic amino acids beta-N-methylamino-l-alanine (BMAA) and 2,4-diamino-butyric acid (DAB) in shallow springs from the Gobi Desert. Amyotroph. Lateral Scler. 2009, 10 (Suppl. 2), 96–100. [Google Scholar] [CrossRef]
- Major, Y.; Kifle, D.; Spoof, L.; Meriluoto, J. Cyanobacteria and microcystins in Koka reservoir (Ethiopia). Environ. Sci. Pollut. Res. Int. 2018, 25, 26861–26873. [Google Scholar] [CrossRef]
- Bouhaddada, R.; Nelieu, S.; Nasri, H.; Delarue, G.; Bouaicha, N. High diversity of microcystins in a Microcystis bloom from an Algerian lake. Environ. Pollut. 2016, 216, 836–844. [Google Scholar] [CrossRef]
- Kimambo, O.N.; Gumbo, J.R.; Chikoore, H. The occurrence of cyanobacteria blooms in freshwater ecosystems and their link with hydro-meteorological and environmental variations in Tanzania. Heliyon 2019, 5, e01312. [Google Scholar] [CrossRef]
- Simiyu, B.M.; Oduor, S.O.; Rohrlack, T.; Sitoki, L.; Kurmayer, R. Microcystin content in phytoplankton and in small fish from eutrophic Nyanza Gulf, Lake Victoria, Kenya. Toxins 2018, 10, 275. [Google Scholar] [CrossRef]
- Ballot, A.; Sandvik, M.; Rundberget, T.; Botha, C.; Miles, C. Diversity of cyanobacteria and cyanotoxins in Hartbeespoort Dam, South Africa. Mar. Freshw. Res. 2014, 65, 175–189. [Google Scholar] [CrossRef]
- Herry, S.; Fathalli, A.; Jenhani, A.; Bouaïcha, N. Seasonal occurrence and toxicity of Microcystis spp. and Oscillatoria tenuis in the Lebna Dam, Tunisia. Water Res. 2008, 42, 1263–1273. [Google Scholar] [CrossRef]
- Ndebele, M.R.; Magadza, C.H.D. The occurrence of microcystin-LR in Lake Chivero, Zimbabwe. Lakes Reserv. Res. Manag. 2006, 11, 57–62. [Google Scholar] [CrossRef]
- Douma, M.; Ouahid, Y.; Loudiki, M.; Del Campo, F.F.; Oudra, B. The first detection of potentially toxic Microcystis strains in two Middle Atlas Mountains natural lakes (Morocco). Environ. Monit. Assess. 2017, 189, 39. [Google Scholar] [CrossRef]
- Okello, W.; Kurmayer, R. Seasonal development of cyanobacteria and microcystin production in Ugandan freshwater lakes. Lakes Reserv. Res. Manag. 2011, 16, 123–135. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Deyab, M.A.; Abou-Dobara, M.I.; El-Sayed, A.K.; El-Raghi, W.M. Occurrence of cyanobacteria and microcystin toxins in raw and treated waters of the Nile River, Egypt: Implication for water treatment and human health. Environ. Sci. Pollut. Res. 2015, 22, 11716–11727. [Google Scholar] [CrossRef]
- Chia, A.M.; Abolude, D.S.; Ladan, Z.; Akanbi, O.; Kalaboms, A. The presence of microcystins in aquatic ecosystems in Northern Nigeria: Zaria as a case study. Res. J. Environ. Toxicol. 2009, 3, 170–178. [Google Scholar] [CrossRef]
- Addico, G.N.D. Toxic Cyanobacteria, Cyanotoxins and Drinking Water Production in Ghana: Implications to Human Health. Ph.D. Thesis, University of Hull, Hull, UK, 2008. [Google Scholar]
- Mohamed, Z.A.; Bakr, A. Concentrations of cylindrospermopsin toxin in water and tilapia fish of tropical fishponds in Egypt, and assessing their potential risk to human health. Environ. Sci. Pollut. Res. Int. 2018, 25, 36287–36297. [Google Scholar] [CrossRef]
- Odokuma, L.O.; Isirima, J.C. Distribution of cyanotoxins in aquatic environments in the Niger Delta. Afr. J. Biotechnol. 2007, 6, 2375–2385. [Google Scholar]
- Harding, W.R.; Rowe, N.; Wessels, J.C.; Beattie, K.A.; Codd, G.A. Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 256–259. [Google Scholar]
- Magonono, M.; Oberholster, P.J.; Addmore, S.; Stanley, M.; Gumbo, J.R. The presence of toxic and non-toxic Cyanobacteria in the sediments of the Limpopo River Basin: Implications for human health. Toxins 2018, 10, 269. [Google Scholar] [CrossRef]
- Naouli, J.; Abouabdellah, R.; Bennouna, A.; Laissaoui, A.; Swarzenski, P.W.; Ait Bouh, H.; Mesfioui, A.; Benbrahim, M.S.; Dechraoui Bottein, M.Y. Using the radioligand-receptor binding assay for paralytic shellfish toxins: A case study on shellfish from Morocco. J. Environ. Radioact. 2018, 192, 485–490. [Google Scholar] [CrossRef]
- Mussagy, A.; Annadotter, H.; Cronberg, G. An experimental study of toxin production in Arthrospira fusiformis (Cyanophyceae) isolated from African waters. Toxicon 2006, 48, 1027–1034. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Downing, T.G. Beta-N-methylamino-l-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Tatters, A.O.; Howard, M.D.; Nagoda, C.; Busse, L.; Gellene, A.G.; Caron, D.A. Multiple stressors at the land-sea interface: Cyanotoxins at the land-sea interface in the Southern California Bight. Toxins 2017, 9. [Google Scholar] [CrossRef]
- Wiltsie, D.; Schnetzer, A.; Green, J.; Vander Borgh, M.; Fensin, E. Algal blooms and Cyanotoxins in Jordan Lake, North Carolina. Toxins 2018, 10, 92. [Google Scholar] [CrossRef]
- Foss, A.J.; Butt, J.; Aubel, M.T. Benthic periphyton from Pennsylvania, USA is a source for both hepatotoxins (microcystins/nodularin) and neurotoxins (anatoxin-a/homoanatoxin-a). Toxicon 2018, 150, 13–16. [Google Scholar] [CrossRef]
- Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L.; Miller, T.R. Variable cyanobacterial toxin and metabolite profiles across six eutrophic lakes of differing physiochemical characteristics. Toxins 2017, 9, 62. [Google Scholar] [CrossRef]
- Banack, S.A.; Metcalf, J.S.; Bradley, W.G.; Cox, P.A. Detection of cyanobacterial neurotoxin beta-N-methylamino-l-alanine within shellfish in the diet of an ALS patient in Florida. Toxicon 2014, 90, 167–173. [Google Scholar] [CrossRef]
- Zastepa, A.; Taranu, Z.E.; Kimpe, L.E.; Blais, J.M.; Gregory-Eaves, I.; Zurawell, R.W.; Pick, F.R. Reconstructing a long-term record of microcystins from the analysis of lake sediments. Sci. Total Environ. 2017, 579, 893–901. [Google Scholar] [CrossRef]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S. Determination of BMAA and three alkaloid cyanotoxins in lake water using dansyl chloride derivatization and high-resolution mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 5487–5501. [Google Scholar] [CrossRef]
- Lajeunesse, A.; Segura, P.A.; Gelinas, M.; Hudon, C.; Thomas, K.; Quilliam, M.A.; Gagnon, C. Detection and confirmation of saxitoxin analogues in freshwater benthic Lyngbya wollei algae collected in the St. Lawrence River (Canada) by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2012, 1219, 93–103. [Google Scholar] [CrossRef]
- Zamora-Barrios, C.A.; Nandini, S.; Sarma, S.S.S. Bioaccumulation of microcystins in seston, zooplankton and fish: A case study in Lake Zumpango, Mexico. Environ. Pollut. 2019, 249, 267–276. [Google Scholar] [CrossRef]
- Berry, J.P.; Jaja-Chimedza, A.; Davalos-Lind, L.; Lind, O. Apparent bioaccumulation of cylindrospermopsin and paralytic shellfish toxins by finfish in Lake Catemaco (Veracruz, Mexico). Food Addit. Contam. Part A 2012, 29, 314–321. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Martins, A.; Vale, M.; Antunes, A.; Azevedo, J.; Welker, M.; Lopez, O.; Montejano, G. First report on the occurrence of microcystins in planktonic cyanobacteria from Central Mexico. Toxicon 2010, 56, 425–431. [Google Scholar] [CrossRef]
- Trout-Haney, J.V.; Wood, Z.T.; Cottingham, K.L. Presence of the cyanotoxin microcystin in Arctic Lakes of Southwestern Greenland. Toxins 2016, 8, 256. [Google Scholar] [CrossRef]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Kupper, F.C.; Dietrich, D. Potent toxins in Arctic environments—Presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem.-Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef]
- Jokela, J.; Heinila, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of high amounts of hepatotoxin nodularin and new protease inhibitors pseudospumigins by the Brazilian benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef]
- Walter, J.M.; Lopes, F.A.C.; Lopes-Ferreira, M.; Vidal, L.M.; Leomil, L.; Melo, F.; de Azevedo, G.S.; Oliveira, R.M.S.; Medeiros, A.J.; Melo, A.S.O.; et al. Occurrence of harmful cyanobacteria in drinking water from a severely drought-impacted semi-arid region. Front. Microbiol. 2018, 9, 176. [Google Scholar] [CrossRef]
- Casali, S.P.; Dos Santos, A.C.A.; de Falco, P.B.; Calijuri, M.D.C. Influence of environmental variables on saxitoxin yields by Cylindrospermopsis raciborskii in a mesotrophic subtropical reservoir. J. Water Health 2017, 15, 509–518. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef]
- Barón-Sola, Á.; Ouahid, Y.; del Campo, F.F. Detection of potentially producing cylindrospermopsin and microcystin strains in mixed populations of cyanobacteria by simultaneous amplification of cylindrospermopsin and microcystin gene regions. Ecotoxicol. Env. Saf. 2012, 75, 102–108. [Google Scholar] [CrossRef]
- Nimptsch, J.; Woelfl, S.; Osorio Ruiz, S.; Valenzuela, J.; Moreira, C.; Ramos, V.; Castelo Branco, R.; Leao, P.; Vasconcelos, V. First record of toxins associated with cyanobacterial blooms in oligotrophic North. Patagonian lakes of Chile—A genomic approach. Int. Rev. Hydrobiol. 2016, 101, 57–168. [Google Scholar] [CrossRef]
- Vidal, L.; Kruk, C. Cylindrospermopsis raciborskii (Cyanobacteria) extends its distribution to Latitude 34°53’ S: Taxonomical and ecological features in Uruguayan eutrophic lakes. Pan-Am. J. Aquat. Sci. 2008, 3, 142–151. [Google Scholar]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P.; Holland, P.T. Further characterization of glycine-containing microcystins from the McMurdo dry Valleys of Antarctica. Toxins 2015, 7, 493–515. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Lampert, C.S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D.R. Toxin production in cyanobacterial mats from ponds on the McMurdo ice shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef]
- Kleinteich, J.; Hildebrand, F.; AWood, S.; Cirés, S.; Agha, R.; Quesada, A.; Pearce, D.; Convey, P.; Küpper, F.; Dietrich, D. Diversity of toxin and non-toxin containing cyanobacterial mats of meltwater ponds on the Antarctic Peninsula: A pyrosequencing approach. Antarct. Sci. 2014, 26, 521–532. [Google Scholar] [CrossRef]
- Genuario, D.B.; Correa, D.M.; Komarek, J.; Fiore, M.F. Characterization of freshwater benthic biofilm-forming Hydrocoryne (Cyanobacteria) isolates from Antarctica. J. Phycol. 2013, 49, 1142–1153. [Google Scholar] [CrossRef]
- Ceglowska, M.; Torunska-Sitarz, A.; Kowalewska, G.; Mazur-Marzec, H. Specific chemical and genetic markers revealed a thousands-year presence of toxic nodularia spumigena in the Baltic Sea. Mar. Drugs 2018, 16, 116. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Sun, F.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Isolation and characterization of a variety of microcystins from seven strains of the cyanobacterial genus Anabaena. Appl. Environ. Microbiol. 1992, 58, 2495–2500. [Google Scholar] [CrossRef]
- Chernova, E.; Sidelev, S.; Russkikh, I.; Voyakina, E.; Zhakovskaya, Z. First observation of microcystin- and anatoxin-a-producing cyanobacteria in the easternmost part of the Gulf of Finland (the Baltic Sea). Toxicon 2019, 157, 18–24. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Fastner, J.; Borner, T. Abundance of active and inactive microcystin genotypes in populations of the toxic cyanobacterium Planktothrix spp. Environ. Microbiol. 2004, 6, 831–841. [Google Scholar] [CrossRef]
- Gkelis, S.; Lanaras, T.; Sivonen, K. Cyanobacterial toxic and bioactive peptides in freshwater bodies of Greece: Concentrations, occurrence patterns, and implications for human health. Mar. Drugs 2015, 13, 6319–6335. [Google Scholar] [CrossRef]
- Grabowska, M.; Kobos, J.; Toruńska-Sitarz, A.; Mazur-Marzec, H. Non-ribosomal peptides produced by Planktothrix agardhii from Siemianówka Dam Reservoir SDR (northeast Poland). Arch. Microbiol. 2014, 196, 697–707. [Google Scholar] [CrossRef]
- Touzet, N.; McCarthy, D.; Gill, A.; Fleming, G.T.A. Comparative summer dynamics of surface cyanobacterial communities in two connected lakes from the west of Ireland. Sci. Total Environ. 2016, 553, 416–428. [Google Scholar] [CrossRef]
- Lopes, V.R.; Ramos, V.; Martins, A.; Sousa, M.; Welker, M.; Antunes, A.; Vasconcelos, V.M. Phylogenetic, chemical and morphological diversity of cyanobacteria from Portuguese temperate estuaries. Mar. Environ. Res. 2012, 73, 7–16. [Google Scholar] [CrossRef]
- Farkas, O.; Gyemant, G.; Hajdu, G.; Gonda, S.; Parizsa, P.; Horgos, T.; Mosolygo, A.; Vasas, G. Variability of microcystins and its synthetase gene cluster in Microcystis and Planktothrix waterblooms in shallow lakes of Hungary. Acta Biol. Hung. 2014, 65, 227–239. [Google Scholar] [CrossRef]
- Handeland, K.; Ostensvik, O. Microcystin poisoning in roe deer (Capreolus capreolus). Toxicon 2010, 56, 1076–1078. [Google Scholar] [CrossRef]
- Faassen, E.J.; Lurling, M. Occurrence of the microcystins MC-LW and MC-LF in Dutch surface waters and their contribution to total microcystin toxicity. Mar. Drugs 2013, 11, 2643–2654. [Google Scholar] [CrossRef]
- Jancula, D.; Strakova, L.; Sadilek, J.; Marsalek, B.; Babica, P. Survey of cyanobacterial toxins in Czech water reservoirs—the first observation of neurotoxic saxitoxins. Env. Sci. Pollut. Res. Int. 2014, 21, 8006–8015. [Google Scholar] [CrossRef]
- Rücker, J.; Wiedner, C.; Zippel, P. Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. Hydrobiologia 1997, 342, 107–115. [Google Scholar] [CrossRef]
- Pekar, H.; Westerberg, E.; Bruno, O.; Laane, A.; Persson, K.M.; Sundstrom, L.F.; Thim, A.M. Fast, rugged and sensitive ultra high pressure liquid chromatography tandem mass spectrometry method for analysis of cyanotoxins in raw water and drinking water—First findings of anatoxins, cylindrospermopsins and microcystin variants in Swedish source waters and infiltration ponds. J. Chromatogr. A 2016, 1429, 265–276. [Google Scholar] [CrossRef]
- Aboal, M. Are we underestimating benthic cyanotoxins? Extensive sampling results from Spain. Toxins 2017, 9, 385. [Google Scholar] [CrossRef]
- Krzton, W.; Pudas, K.; Pociecha, A.; Strzesak, M.; Kosiba, J.; Walusiak, E.; Szarek-Gwiazda, E.; Wilk-Wozniak, E. Microcystins affect zooplankton biodiversity in oxbow lakes. Environ. Toxicol. Chem. 2017, 36, 165–174. [Google Scholar] [CrossRef]
- Hunter, P.D.; Tyler, A.N.; Carvalho, L.; Codd, G.A.; Maberly, S.C. Hyperspectral remote sensing of cyanobacterial pigments as indicators for cell populations and toxins in eutrophic lakes. Remote Sens. Environ. 2010, 114, 2705–2718. [Google Scholar] [CrossRef]
- Teneva, I.; Mladenov, R.; Belkinova, D.; Dimitrova-Dyulgerova, I.; Dzhambazov, B. Phytoplankton community of the drinking water supply reservoir Borovitsa (South Bulgaria) with an emphasis on cyanotoxins and water quality. Cent. Eur. J. Biol. 2010, 5, 231–239. [Google Scholar] [CrossRef]
- Osswald, J.; Rellan, S.; Gago-Martinez, A.; Vasconcelos, V. Production of anatoxin-a by cyanobacterial strains isolated from Portuguese fresh water systems. Ecotoxicology 2009, 18, 1110–1115. [Google Scholar] [CrossRef]
- Sabart, M.; Misson, B.; Descroix, A.; Duffaud, E.; Combourieu, B.; Salencon, M.J.; Latour, D. The importance of small colonies in sustaining Microcystis population exposed to mixing conditions: An exploration through colony size, genotypic composition and toxic potential. Environ. Microbiol. Rep. 2013, 5, 747–756. [Google Scholar] [CrossRef]
- Pitois, F.; Fastner, J.; Pagotto, C.; Dechesne, M. Multi-toxin occurrences in ten french water resource reservoirs. Toxins 2018, 10, 283. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Lujic, J.; Marinovic, Z.; Subakov-Simic, G.; Dulic, T.; Vazic, T.; Nybom, S.; Meriluoto, J.; Codd, G.A.; et al. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef]
- Belykh, O.I.; Gladkikh, A.S.; Sorokovikova, E.G.; Tikhonova, I.V.; Butina, T.V. Identification of toxic Cyanobacteria in Lake Baikal. Dokl. Biochem. Biophys. 2015, 463, 220–224. [Google Scholar] [CrossRef]
- Sulcius, S.; Pilkaityte, R.; Mazur-Marzec, H.; Kasperoviciene, J.; Ezhova, E.; Blaszczyk, A.; Paskauskas, R. Increased risk of exposure to microcystins in the scum of the filamentous cyanobacterium Aphanizomenon flos-aquae accumulated on the western shoreline of the Curonian Lagoon. Mar. Pollut. Bull. 2015, 99, 264–270. [Google Scholar] [CrossRef]
- Sidelev, S.I.; Golokolenova, T.B.; Chernova, E.N.; Russkikh, I.V. Analysis of phytoplankton in Tsimlyansk Reservoir (RUSSIA) for the presence of cyanobacterial hepato- and neurotoxins. Mikrobiologiia 2015, 84, 732–742. [Google Scholar] [CrossRef]
- Barda, I.; Kankaanaaa, H.; Purina, I.; Balode, M.; Sjovall, O.; Meriluoto, J. Bioaccumulation of hepatotoxins—A considerable risk in the Latvian environment. Environ. Pollut. 2015, 196, 313–320. [Google Scholar] [CrossRef]
- Greer, B.; McNamee, S.E.; Boots, B.; Cimarelli, L.; Guillebault, D.; Helmi, K.; Marcheggiani, S.; Panaiotov, S.; Breitenbach, U.; Akcaalan, R.; et al. A validated UPLC-MS/MS method for the surveillance of ten aquatic biotoxins in European brackish and freshwater systems. Harmful Algae 2016, 55, 31–40. [Google Scholar] [CrossRef]
- Preussel, K.; Stuken, A.; Wiedner, C.; Chorus, I.; Fastner, J. First report on cylindrospermopsin producing Aphanizomenon flos-aquae (Cyanobacteria) isolated from two German lakes. Toxicon 2006, 47, 156–162. [Google Scholar] [CrossRef]
- Brient, L.; Lengronne, M.; Bormans, M.; Fastner, J. First occurrence of cylindrospermopsin in freshwater in France. Environ. Toxicol. 2009, 24, 415–420. [Google Scholar] [CrossRef]
- Messineo, V.; Melchiorre, S.; Di Corcia, A.; Gallo, P.; Bruno, M. Seasonal succession of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum blooms with cylindrospermopsin occurrence in the volcanic Lake Albano, Central Italy. Environ. Toxicol. 2010, 25, 18–27. [Google Scholar] [CrossRef]
- Bláhová, L.; Babica, P.; Adamovský, O.; Kohoutek, J.; Maršálek, B.; Bláha, L. Analyses of cyanobacterial toxins (microcystins, cylindrospermopsin) in the reservoirs of the Czech Republic and evaluation of health risks. Environ. Chem. Lett. 2008, 6, 223–227. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef]
- Cires, S.; Ballot, A. A review of the phylogeny, ecology and toxin production of bloom-forming Aphanizomenon spp. and related species within the Nostocales (cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef]
- Gkelis, S.; Zaoutsos, N. Cyanotoxin occurrence and potentially toxin producing cyanobacteria in freshwaters of Greece: A multi-disciplinary approach. Toxicon 2014, 78, 1–9. [Google Scholar] [CrossRef]
- Spoof, L.; Berg, K.A.; Rapala, J.; Lahti, K.; Lepistö, L.; Metcalf, J.S.; Codd, G.A.; Meriluoto, J. First observation of cylindrospermopsin in Anabaena lapponica isolated from the boreal environment (Finland). Environ. Toxicol. 2006, 21, 552–560. [Google Scholar] [CrossRef]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. 2013, 20, 5243–5264. [Google Scholar] [CrossRef]
- Đorđević, N.; Simić, S.; Ciric, A. First identification of the cylindrospermopsin (CYN) producting by cyanobacterium Cylindrospermopsis raciborskii (Woloszynska) Seenayya & Subba Raju in Serbia. Fresenius Environ. Bull. 2015, 24, 3736–3742. [Google Scholar]
- Moreira, C.; Mendes, R.; Azevedo, J.; Vasconcelos, V.; Antunes, A. First occurrence of cylindrospermopsin in Portugal: A contribution to its continuous global dispersal. Toxicon 2017, 130, 87–90. [Google Scholar] [CrossRef]
- Antal, O.; Karisztl-Gácsi, M.; Farkas, A.; Kovács, A.; Ács, A.; Törő, N.; Kiss, G.; Saker, M.L.; Győri, J.; Bánfalvi, G.; et al. Screening the toxic potential of Cylindrospermopsis raciborskii strains isolated from Lake Balaton, Hungary. Toxicon 2011, 57, 831–840. [Google Scholar] [CrossRef]
- James, K.J.; Sherlock, I.R.; Stack, M.A. Anatoxin-a in Irish freshwater and cyanobacteria, determined using a new fluorimetric liquid chromatographic method. Toxicon 1997, 35, 963–971. [Google Scholar] [CrossRef]
- Shams, S.; Capelli, C.; Cerasino, L.; Ballot, A.; Dietrich, D.R.; Sivonen, K.; Salmaso, N. Anatoxin-a producing Tychonema (Cyanobacteria) in European waterbodies. Water Res. 2015, 69, 68–79. [Google Scholar] [CrossRef]
- Echenique-Subiabre, I.; Tenon, M.; Humbert, J.F.; Quiblier, C. Spatial and Temporal Variability in the Development and Potential Toxicity of Phormidium Biofilms in the Tarn River, France. Toxins 2018, 10, 418. [Google Scholar] [CrossRef]
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lurling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef]
- Onodera, H.; Oshima, Y.; Henriksen, P.; Yasumoto, T. Confirmation of anatoxin-a(s), in the cyanobacterium Anabaena lemmermannii, as the cause of bird kills in Danish Lakes. Toxicon 1997, 35, 1645–1648. [Google Scholar] [CrossRef]
- Sieroslawska, A.; Rymuszka, A.; Kalinowska, R.; Skowronski, T.; Bownik, A.; Pawlik-Skowronska, B. Toxicity of cyanobacterial bloom in the eutrophic dam reservoir (Southeast Poland). Environ. Toxicol. Chem. 2010, 29, 556–560. [Google Scholar] [CrossRef]
- Rivetti, C.; Gomez-Canela, C.; Lacorte, S.; Diez, S.; Lazaro, W.L.; Barata, C. Identification of compounds bound to suspended solids causing sub-lethal toxic effects in Daphnia magna. A field study on re-suspended particles during river floods in Ebro River. Aquat. Toxicol. 2015, 161, 41–50. [Google Scholar] [CrossRef]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-A in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef]
- Ballot, A.; Fastner, J.; Lentz, M.; Wiedner, C. First report of anatoxin-a-producing cyanobacterium Aphanizomenon issatschenkoi in northeastern Germany. Toxicon 2010, 56, 964–971. [Google Scholar] [CrossRef]
- Ballot, A.; Scherer, P.I.; Wood, S.A. Variability in the anatoxin gene clusters of Cuspidothrix issatschenkoi from Germany, New Zealand, China and Japan. PLoS ONE 2018, 13, e0200774. [Google Scholar] [CrossRef]
- Kaas, H.; Henriksen, P. Saxitoxins (PSP toxins) in Danish Lakes. Water Res. 2000, 34, 2089–2097. [Google Scholar] [CrossRef]
- Ledreux, A.; Thomazeau, S.; Catherine, A.; Duval, C.; Yéprémian, C.; Marie, A.; Bernard, C. Evidence for saxitoxins production by the cyanobacterium Aphanizomenon gracile in a French recreational water body. Harmful Algae 2010, 10, 88–97. [Google Scholar] [CrossRef]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef]
- Pomati, F.; Sacchi, S.; Rossetti, C.; Giovannardi, S.; Onodera, H.; Oshima, Y.; Neilan, B.A. The freshwater cyanobacterium Planktothrix sp. fp1: Molecular identification and detection of paralytic shellfish poisoning toxins. J. Phycol. 2000, 36, 553–562. [Google Scholar] [CrossRef]
- Rapala, J.; Robertson, A.; Negri, A.P.; Berg, K.A.; Tuomi, P.; Lyra, C.; Erkomaa, K.; Lahti, K.; Hoppu, K.; Lepisto, L. First report of saxitoxin in Finnish lakes and possible associated effects on human health. Environ. Toxicol. 2005, 20, 331–340. [Google Scholar] [CrossRef]
- Usleber, E.; Donald, M.; Straka, M.; Märtlbauer, E. Comparison of enzyme immunoassay and mouse bioassay for determining paralytic shellfish poisoning toxins in shellfish. Food Addit. Contam. 1997, 14, 193–198. [Google Scholar] [CrossRef]
- Savela, H.; Spoof, L.; Perälä, N.; Vehniäinen, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Kokocinski, M.; Meriluoto, J. First report of cyanobacterial paralytic shellfish toxin biosynthesis genes and paralytic shellfish toxin production in Polish freshwater lakes. Adv. Oceanogr. Limnol. 2017, 8. [Google Scholar] [CrossRef]
- Ballot, A.; Cerasino, L.; Hostyeva, V.; Cires, S. Variability in the sxt gene clusters of PSP toxin producing Aphanizomenon gracile strains from Norway, Spain, Germany and North America. PLoS ONE 2016, 11, e0167552. [Google Scholar] [CrossRef]
- Beltran, E.; Ibanez, M.; Sancho, J.V.; Hernandez, F. Determination of six microcystins and nodularin in surface and drinking waters by on-line solid phase extraction-ultra high pressure liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2012, 1266, 61–68. [Google Scholar] [CrossRef]
- Beattie, K.A.; Kaya, K.; Codd, G.A. The cyanobacterium Nodularia PCC 7804, of freshwater origin, produces [L-Har2]nodularin. Phytochemistry 2000, 54, 57–61. [Google Scholar] [CrossRef]
- Eiser, A.R. Why does Finland have the highest dementia mortality rate? Environmental factors may be generalizable. Brain Res. 2017, 1671, 14–17. [Google Scholar] [CrossRef]
- Rosen, J.; Westerberg, E.; Hellenas, K.E.; Salomonsson, M.L. A new method for analysis of underivatized free β-methylamino-alanine: Validation and method comparison. Toxicon 2016, 121, 105–108. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef]
- Luglie, A.; Giacobbe, M.G.; Riccardi, E.; Bruno, M.; Pigozzi, S.; Mariani, M.A.; Satta, C.T.; Stacca, D.; Bazzoni, A.M.; Caddeo, T.; et al. Paralytic shellfish toxins and cyanotoxins in the Mediterranean: New data from Sardinia and Sicily (Italy). Microorganisms 2017, 5, 72. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Mankiewicz-Boczek, J.; Faassen, E.; Jurczak, T.; Gągała, I.; Ballot, A.; Lurling, M.; Kokocinski, M. Polyphasic toxicological screening of Cylindrospermopsis raciborskii and Aphanizomenon gracile isolated in Poland. Algal Res. 2017, 24, 72–80. [Google Scholar] [CrossRef]
- Metcalf, J.; Anne Banack, S.; Lindsay, J.; FMorrison, L.; Alan Cox, P.; ACodd, G. Co-occurrence of β-N-methylamino-l-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA exposure in an amyotrophic lateral sclerosis cluster from southern France. PLoS ONE 2013, 8, e83406. [Google Scholar] [CrossRef]
- Violi, J.P.; Mitrovic, S.M.; Colville, A.; Main, B.J.; Rodgers, K.J. Prevalence of β-methylamino-l-alanine (BMAA) and its isomers in freshwater cyanobacteria isolated from eastern Australia. Ecotoxicol. Environ. Saf. 2019, 172, 72–81. [Google Scholar] [CrossRef]
- Gaget, V.; Humpage, A.R.; Huang, Q.; Monis, P.; Brookes, J.D. Benthic cyanobacteria: A source of cylindrospermopsin and microcystin in Australian drinking water reservoirs. Water Res. 2017, 124, 454–464. [Google Scholar] [CrossRef]
- Wood, S.A.; Maier, M.Y.; Puddick, J.; Pochon, X.; Zaiko, A.; Dietrich, D.R.; Hamilton, D.P. Trophic state and geographic gradients influence planktonic cyanobacterial diversity and distribution in New Zealand lakes. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Wood, S.A.; Stirling, D.J. First identification of the cylindrospermopsin-producing cyanobacterium Cylindrospermopsis raciborskii in New Zealand. N. Z. J. Mar. Freshw. Res. 2003, 37, 821–828. [Google Scholar] [CrossRef]
- Smith, F.M.; Wood, S.A.; van Ginkel, R.; Broady, P.A.; Gaw, S. First report of saxitoxin production by a species of the freshwater benthic cyanobacterium, Scytonema Agardh. Toxicon 2011, 57, 566–573. [Google Scholar] [CrossRef]
- Vogiazi, V.; de la Cruz, A.; Mishra, S.; Shanov, V.; Heineman, W.R.; Dionysiou, D.D. A comprehensive review: Development of electrochemical biosensors for detection of cyanotoxins in freshwater. ACS Sens. 2019, 4, 1151–1173. [Google Scholar] [CrossRef]
- Straser, A.; Filipic, M.; Zegura, B. Cylindrospermopsin induced transcriptional responses in human hepatoma HepG2 cells. Toxicol. In Vitro 2013, 27, 1809–1819. [Google Scholar] [CrossRef]
- Vasas, G.; Gaspar, A.; Suranyi, G.; Batta, G.; Gyemant, G.; M, M.H.; Mathe, C.; Grigorszky, I.; Molnar, E.; Borbely, G. Capillary electrophoretic assay and purification of cylindrospermopsin, a cyanobacterial toxin from Aphanizomenon ovalisporum, by plant test (blue-green Sinapis test). Anal. Biochem. 2002, 302, 95–103. [Google Scholar] [CrossRef]
- McElhiney, J.; Lawton, L.A.; Leifert, C. Investigations into the inhibitory effects of microcystins on plant growth, and the toxicity of plant tissues following exposure. Toxicon 2001, 39, 1411–1420. [Google Scholar] [CrossRef]
- Wu, Z.; He, D.; Cui, B.; Jin, Z. Ultrasensitive detection of microcystin-LR with gold immunochromatographic assay assisted by a molecular imprinting technique. Food Chem. 2019, 283, 517–521. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, J.; Beloglazova, N.; Yang, S.; De Saeger, S.; Mari, G.M.; Zhang, S.; Shen, J.; Wang, Z.; Yu, X. Portable multiplex immunochromatographic assay for quantitation of two typical algae toxins based on dual-color fluorescence microspheres. J. Agric. Food Chem. 2019, 67, 6041–6047. [Google Scholar] [CrossRef]
- Cunha, I.; Biltes, R.; Sales, M.; Vasconcelos, V. Aptamer-based biosensors to detect aquatic phycotoxins and cyanotoxins. Sensors 2018, 18, 2367. [Google Scholar] [CrossRef]
- Wharton, R.E.; Feyereisen, M.C.; Gonzalez, A.L.; Abbott, N.L.; Hamelin, E.I.; Johnson, R.C. Quantification of saxitoxin in human blood by ELISA. Toxicon 2017, 133, 110–115. [Google Scholar] [CrossRef]
- Hicks, J.M. Fluorescence immunoassay. Hum. Pathol. 1984, 15, 112–116. [Google Scholar] [CrossRef]
- Rossi, A.M.; Taylor, C.W. Analysis of protein-ligand interactions by fluorescence polarization. Nat. Protoc. 2011, 6, 365–387. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.S.; Eremin, S.A. Fluorescence polarization immunoassays and related methods for simple, high-throughput screening of small molecules. Anal. Bioanal. Chem. 2008, 391, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Bahadır, E.B.; Sezgintürk, M.K. Lateral flow assays: Principles, designs and labels. TrAC Trends Anal. Chem. 2016, 82, 286–306. [Google Scholar] [CrossRef]
- Xing, C.; Liu, L.; Song, S.; Feng, M.; Kuang, H.; Xu, C. Ultrasensitive immunochromatographic assay for the simultaneous detection of five chemicals in drinking water. Biosens. Bioelectron. 2015, 66, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Srivastava, A.; Oh, H.M.; Ahn, C.Y.; Choi, G.G.; Asthana, R.K. Recent trends in development of biosensors for detection of microcystin. Toxicon 2012, 60, 878–894. [Google Scholar] [CrossRef]
- Bickman, S.R.; Campbell, K.; Elliott, C.; Murphy, C.; O’Kennedy, R.; Papst, P.; Lochhead, M.J. An innovative portable biosensor system for the rapid detection of freshwater cyanobacterial algal bloom toxins. Environ. Sci. Technol. 2018, 52, 11691–11698. [Google Scholar] [CrossRef]
- Metcalf, J.; Hiskia, A.; Kaloudis, T. Protein Phosphatase Inhibition Assays. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 26, 267–271. [Google Scholar] [CrossRef]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef]
- Heresztyn, T.; Nicholson, B.C. Determination of cyanobacterial hepatotoxins directly in water using a protein phosphatase inhibition assay. Water Res. 2001, 35, 3049–3056. [Google Scholar] [CrossRef]
- Ruberu, S.R.; Langlois, G.W.; Masuda, M.; Kittredge, C.; Perera, S.K.; Kudela, R.M. Receptor binding assay for the detection of paralytic shellfish poisoning toxins: Comparison to the mouse bioassay and applicability under regulatory use. Food Addit. Contam. Part A 2018, 35, 144–158. [Google Scholar] [CrossRef]
- Araoz, R.; Vilarino, N.; Botana, L.M.; Molgo, J. Ligand-binding assays for cyanobacterial neurotoxins targeting cholinergic receptors. Anal. Bioanal. Chem. 2010, 397, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Ramos, V.; Azevedo, J.; Vasconcelos, V. Methods to detect cyanobacteria and their toxins in the environment. Appl. Microbiol. Biotechnol. 2014, 98, 8073–8082. [Google Scholar] [CrossRef]
- Sun, L.W.; Jiang, W.J.; Zhang, J.Y.; Wang, W.Q.; Du, Y.; Sato, H.; Kawachi, M.; Yu, R. Identification and detection sensitivity of Microcystis aeruginosa from mixed and field samples using MALDI-TOF MS. Environ. Monit. Assess. 2018, 190, 712. [Google Scholar] [CrossRef] [PubMed]
- Nicolau, R.; Leloup, M.; Lachassagne, D.; Pinault, E.; Feuillade-Cathalifaud, G. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) coupled to XAD fractionation: Method to algal organic matter characterization. Talanta 2015, 136, 102–107. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Wu, Z.; Cui, B.; Jin, Z. A novel SERS-based aptasensor for ultrasensitive sensing of microcystin-LR. Food Chem. 2019, 278, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.B.; Guedes, I.A.; Azevedo, S.M. Is qPCR a reliable indicator of cyanotoxin risk in freshwater? Toxins 2016, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Humbert, J.F.; Quiblier, C.; Gugger, M. Molecular approaches for monitoring potentially toxic marine and freshwater phytoplankton species. Anal. Bioanal. Chem. 2010, 397, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Neilan, B.A. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk. Curr. Opin. Biotechnol. 2008, 19, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Singh, S.; Ahn, C.Y.; Oh, H.M.; Asthana, R.K. Monitoring approaches for a toxic cyanobacterial bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef]
- Humbert, J.F. Molecular tools for the detection of toxigenic cyanobacteria in natural ecosystems. Handb. Cyanobacterial Monit. Cyanotoxin Anal. 2017, 28, 280–283. [Google Scholar] [CrossRef]
- McNamee, S.E.; Elliott, C.T.; Greer, B.; Lochhead, M.; Campbell, K. Development of a planar waveguide microarray for the monitoring and early detection of five harmful algal toxins in water and cultures. Environ. Sci. Technol. 2014, 48, 13340–13349. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.; Vilariño, N.; Louzao, M.C.; Rodríguez, L.P.; Alfonso, A.; Campbell, K.; Elliott, C.T.; Taylor, P.; Ramos, V.; Vasconcelos, V.; et al. Multi-detection method for five common microalgal toxins based on the use of microspheres coupled to a flow-cytometry system. Anal. Chim. Acta 2014, 850, 57–64. [Google Scholar] [CrossRef]
- Zeller, P.; Mejean, A.; Biegala, I.; Contremoulins, V.; Ploux, O. Fluorescence in situ hybridization of Microcystis strains producing microcystin using specific mRNA probes. Lett. Appl. Microbiol. 2016, 63, 376–383. [Google Scholar] [CrossRef]
- Johnson, B.N.; Mutharasan, R. A cantilever biosensor-based assay for toxin-producing cyanobacteria Microcystis aeruginosa using 16S rRNA. Environ. Sci. Technol. 2013, 47, 12333–12341. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.; Huang, Q.; Zheng, L.; Song, L. Quantitative assessment of toxic and nontoxic Microcystis colonies in natural environments using fluorescence in situ hybridization and flow cytometry. Sci. China Life Sci. 2010, 53, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, A.; Pernthaler, J.; Amann, R. Fluorescence in situ hybridization and catalyzed reporter deposition for the identification of marine bacteria. Appl. Environ. Microbiol. 2002, 68, 3094–3101. [Google Scholar] [CrossRef]
- Schonhuber, W.; Fuchs, B.; Juretschko, S.; Amann, R. Improved sensitivity of whole-cell hybridization by the combination of horseradish peroxidase-labeled oligonucleotides and tyramide signal amplification. Appl. Env. Microbiol. 1997, 63, 3268–3273. [Google Scholar] [CrossRef]
- Ramm, J.; Lupu, A.; Hadas, O.; Ballot, A.; Rucker, J.; Wiedner, C.; Sukenik, A. A CARD-FISH protocol for the identification and enumeration of cyanobacterial akinetes in lake sediments. FEMS Microbiol. Ecol. 2012, 82, 23–36. [Google Scholar] [CrossRef]
- Riou, V.; Périot, M.; Biegala, I.C. Specificity re-evaluation of oligonucleotide probes for the detection of marine picoplankton by tyramide signal amplification-fluorescent in situ hybridization. Front. Microbiol. 2017, 8, 854. [Google Scholar] [CrossRef]
- Meriluoto, J.; Metcalf, J.; Codd, G.A. Selection of analytical methodology for cyanotoxin analysis. Biochemistry 2017, 32, 309–312. [Google Scholar] [CrossRef]
- Khait, I.; Obolski, U.; Yovel, Y.; Hadany, L. Sound perception in plants. Semin. Cell Dev. Biol. 2019, 92, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zeng, H.; Lin, H.; Wang, J.; Feng, X.; Qiu, Z.; Chen, J.A.; Luo, J.; Luo, Y.; Huang, Y.; et al. Serum microcystin levels positively linked with risk of hepatocellular carcinoma: A case-control study in southwest China. Hepatology 2017, 66, 1519–1528. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Common Name | Primary Toxicity | Analogues | Producing Genera | Reference |
|---|---|---|---|---|---|
| Cyclic peptides | Microcystins | Hepatotoxicity | 246 | Anabaena, Aphanizomenon, Dolichospermum, Fischerella, Geitlerinema, Hapalosiphon, Leptolyngbya, Limnothrix, Merismopedia, Microcystis, Nostoc, Oscillatoria, Phormidium, Planktothrix, Pseudanabaena, Radiocystis, Pseudoanabaena, Spirulina, Synechococcus, Trichodesmium, Woronichinia | [6,11,13,14,20,21,22] |
| Nodularins | Hepatotoxicity | 10 | Iningainema, Nodularia | [10,11,19] | |
| Alkaloids | Cylindrospermopsins | Hepatotoxicity | 5 | Anabaena, Aphanizomenon, Chrysosporum, Cylindrospermopsis, Dolichospermum, Lyngbya, Oscillatoria, Raphidiopsis, Sphaerospermopsis | [13,23,24,25] |
| Saxitoxins | Neurotoxicity | 57 | AnabaenaAphanizomenon, Cuspidothrix, Cylindrospermopsis,Dolichospermum, Fischerella, Geitlerinema, Lyngbya, Phormidium, Planktothrix, Raphidiopsis, Scytonema, Tolypothrix | [11,13,21,26,27,28] | |
| Anatoxin-a | Neurotoxicity | 4 | Anabaena, Aphanizomenon, Arthrospira, Cuspidothrix, Cylindrospermum, Dolichospermum, Microcoleus, Microcystis, Oscillatoria, Phormidium, Planktothrix, Pseudanabaena, Raphidiopsis, Tychonema | [13,29,30] | |
| Anatoxin-a(s) | Neurotoxicity | - | Anabaena | [22,23] | |
| Lyngbyatoxins | Dermatoxicity | 7 | Oscillatoria, Schizothrix, Lyngbya majuscula | [31,32,33,34,35] | |
| Aplysiatoxins | Dermatoxicity | 5 | Lyngbya majuscula | [34,36] | |
| Lipopeptides | Antillatoxins | Neurotoxicity | 2 | Lyngbya majuscula | [37,38] |
| Jamaicamides | Neurotoxicity | 6 | [39,40] | ||
| Kalkitoxins | Neurotoxicity | 7 | [41,42] | ||
| Barbamides | Molluscicidal | 2 | [43] | ||
| Majusculamides | Cytotoxicity | 4 | [31,34,44,45] | ||
| Hectochlorins | Cytotoxicity | 5 | [46,47] | ||
| Curacins | Cytotoxicity | 5 | [48,49,50,51] | ||
| Nonprotein amino acid | β-N-methylamino-l-alanine | Neurotoxicity | 4 | Anabaena, Calothrix, Cyanodictyon, Leptolyngbya, Limnothrix, Microcoleus, Microscystis, Myxosarcina, Nodularia, Nostoc, Oscillatoria, Phormidium, Planktothrix Pseudanabaena, Scytonema, Synechococcus, Trichodesmium | [52,53,54,55,56] |
| Lipoglycans | Lipopolysaccharides | Endotoxicity | - | All genera of cyanobacteria | [57] |
| Regions | Countries | MCs | CYNs, | NODs | STXs | ATXs | BMAA |
|---|---|---|---|---|---|---|---|
| Asia (13 countries) | China | √ | √ | √ | √ | √ | √ |
| Turkey | √ | √ | √ | √ | √ | ||
| Japan | √ | √ | √ | √ | |||
| India and Korea | √ | √ | √ | ||||
| Qatar | √ | √ | √ | ||||
| Bangladesh and Singapore | √ | √ | |||||
| Israel, Saudi Arabia, | √ | √ | |||||
| Thailand and Vietnam | |||||||
| Philippines | √ | ||||||
| Africa (13 countries) | Nigeria | √ | √ | √ | √ | √ | |
| South Africa | √ | √ | √ | ||||
| Egypt | √ | √ | |||||
| Kenya | √ | √ | |||||
| Morocco | √ | √ | |||||
| Algeria, Ethiopia, Ghana, Namibia, Tanzania, Tunisia, Uganda and Zimbabwe | √ | ||||||
| North America (3 countries) | Canada and the US | √ | √ | √ | √ | √ | √ |
| Mexico | √ | √ | √ | ||||
| South America (5 countries) | Brazil | √ | √ | √ | √ | √ | |
| Argentina and Uruguay | √ | √ | |||||
| Chile | √ | ||||||
| Peru | √ | ||||||
| Antarctica | - | √ | √ | √ | √ | ||
| Europe (24 countries) | France | √ | √ | √ | √ | √ | √ |
| Finland, Italy, Poland and Portugal | √ | √ | √ | √ | √ | ||
| Germany and Spain | √ | √ | √ | √ | √ | ||
| Sweden | √ | √ | √ | √ | |||
| the UK | √ | √ | √ | √ | |||
| Bulgaria | √ | √ | √ | ||||
| Czech, Greece and Serbia | √ | √ | √ | ||||
| Ireland | √ | √ | √ | ||||
| Denmark | √ | √ | |||||
| Hungary | √ | √ | |||||
| Latvia | √ | √ | |||||
| Netherlands | √ | √ | |||||
| Norway and Russia | √ | √ | |||||
| Austria, Romania and Switzerland | √ | ||||||
| Lebanon | √ | ||||||
| Oceania (2 countries) | Australia and New Zealand | √ | √ | √ | √ | √ | √ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins 2019, 11, 530. https://doi.org/10.3390/toxins11090530
Du X, Liu H, Yuan L, Wang Y, Ma Y, Wang R, Chen X, Losiewicz MD, Guo H, Zhang H. The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins. 2019; 11(9):530. https://doi.org/10.3390/toxins11090530
Chicago/Turabian StyleDu, Xingde, Haohao Liu, Le Yuan, Yueqin Wang, Ya Ma, Rui Wang, Xinghai Chen, Michael D. Losiewicz, Hongxiang Guo, and Huizhen Zhang. 2019. "The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review" Toxins 11, no. 9: 530. https://doi.org/10.3390/toxins11090530
APA StyleDu, X., Liu, H., Yuan, L., Wang, Y., Ma, Y., Wang, R., Chen, X., Losiewicz, M. D., Guo, H., & Zhang, H. (2019). The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins, 11(9), 530. https://doi.org/10.3390/toxins11090530