Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals


Abstract
1. Underestimated Potent Environmental Disruptor: Mycotoxins
2. Spermatogenesis: A Complex and Delicate Process
2.1. Effect of Mycotoxins on Fertility
2.2. Effect of Mycotoxins on Sertoli Cells
2.3. Effect of Mycotoxins on Leydig Cells
2.4. Effect of Mycotoxins on Spermatogenesis
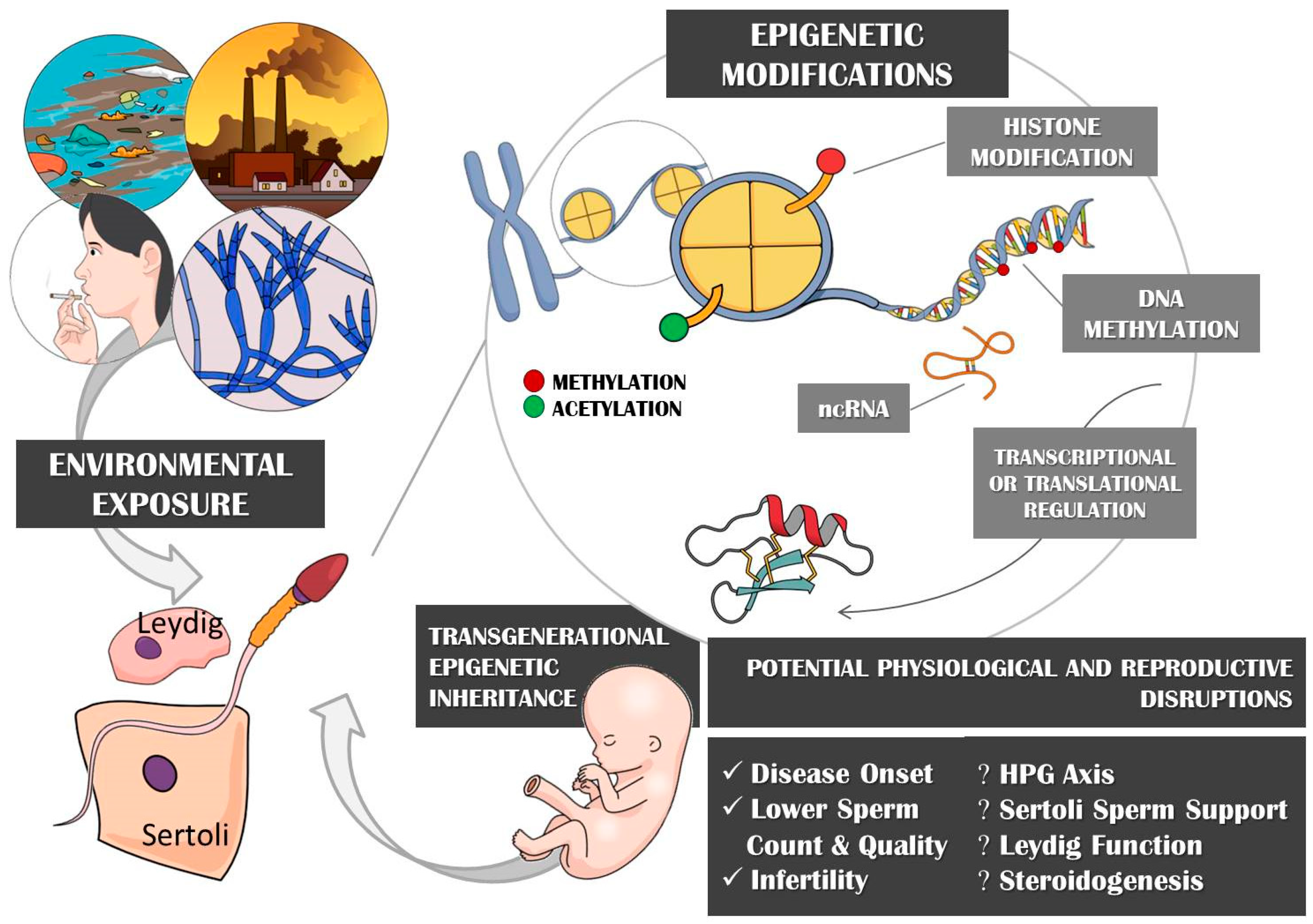
3. Epigenetic Implications
3.1. Epigenetic Mechanisms and Environmental Exposure
3.2. Contribution of Epigenetics in Environmentally-Induced Disease Predisposition
3.3. Epigenetics Involvement in Germline Modulation and Infertility
3.4. Epigenetic Effects of Mycotoxins in Disease and Infertility
3.5. Transgenerational Epigenetic Inheritance through Imprinted Genes
4. Challenges to the Study of the Effect of Mycotoxins on Male Spermatogenesis
5. Conclusion and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Hope, J. A review of the mechanism of injury and treatment approaches for illness resulting from exposure to water-damaged buildings, mold, and mycotoxins. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced neurotoxicity through oxidative stress-associated pathways. Int. J. Mol. Sci. 2011, 12, 5213–5237. [Google Scholar] [CrossRef] [PubMed]
- Brewer, J.; Thrasher, J.; Hooper, D. Chronic illness associated with mold and mycotoxins: Is naso-sinus fungal biofilm the culprit? Toxins 2014, 6, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A. Occurrence of 26 mycotoxins in the grain of cereals cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Assaf, H.; Betbeder, A.-M.; Creppy, E.E.; Pallardy, M.; Azouri, H. Ochratoxin A levels in human plasma and foods in Lebanon. Hum. Exp. Toxicol. 2004, 23, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.; Bertuzzi, T.; Nielsen, K. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Drusch, S.; Ragab, W. Mycotoxins in fruits, fruit juices, and dried fruits. J. Food Prot. 2003, 66, 1514–1527. [Google Scholar] [CrossRef]
- El Khoury, A.; Rizk, T.; Lteif, R.; Azouri, H.; Delia, M.-L.; Lebrihi, A. Occurrence of Ochratoxin A- and Aflatoxin B1-Producing Fungi in Lebanese Grapes and Ochratoxin A Content in Musts and Finished Wines during 2004. J. Agric. Food Chem. 2006, 54, 8977–8982. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Rizk, T.; Lteif, R.; Azouri, H.; Delia, M.-L.; Lebrihi, A. Fungal contamination and Aflatoxin B1 and Ochratoxin A in Lebanese wine–grapes and musts. Food Chem. Toxicol. 2008, 46, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Darwish, W.S.; Ikenaka, Y.; Nakayama, S.M.; Ishizuka, M. An overview on mycotoxin contamination of foods in Africa. J. Vet. Med. Sci. 2014, 76, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Nleya, N.; Adetunji, M.; Mwanza, M. Current status of mycotoxin contamination of food commodities in Zimbabwe. Toxins 2018, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Font, G.; Ruiz, M.; Ferrer, E. Co-occurrence and risk assessment of mycotoxins in food and diet from Mediterranean area. Food Chem. 2012, 135, 423–429. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Joubrane, K.; Khoury, A.E.; Lteif, R.; Rizk, T.; Kallassy, M.; Hilan, C.; Maroun, R. Occurrence of aflatoxin B1 and ochratoxin A in Lebanese cultivated wheat. Mycotoxin Res. 2011, 27, 249–257. [Google Scholar] [CrossRef]
- Raad, F.; Nasreddine, L.; Hilan, C.; Bartosik, M.; Parent-Massin, D. Dietary exposure to aflatoxins, ochratoxin A and deoxynivalenol from a total diet study in an adult urban Lebanese population. Food Chem. Toxicol. 2014, 73, 35–43. [Google Scholar] [CrossRef]
- Eze, U.A.; Okonofua, F.E. High Prevalence of Male Infertility in Africa: Are Mycotoxins to Blame? Afr. J. Reprod. Health 2015, 19, 9–17. [Google Scholar]
- Carlsen, E.; Giwercman, A.; Keiding, N.; Skakkebaek, N.E. Evidence for decreasing quality of semen during past 50 years. Bmj 1992, 305, 609–613. [Google Scholar] [CrossRef]
- Stuppia, L.; Franzago, M.; Ballerini, P.; Gatta, V.; Antonucci, I. Epigenetics and male reproduction: The consequences of paternal lifestyle on fertility, embryo development, and children lifetime health. Clin. Epigenet. 2015, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Swan, S.H.; Elkin, E.P.; Fenster, L. Have sperm densities declined? A reanalysis of global trend data. Environ. Health Perspect. 1997, 105, 1228–1232. [Google Scholar] [CrossRef] [PubMed]
- Swan, S.H.; Elkin, E.P.; Fenster, L. The question of declining sperm density revisited: An analysis of 101 studies published 1934–1996. Environ. Health Perspect. 2000, 108, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Ibeh, I.N.; Uraih, N.; Ogonar, J.I. Dietary exposure to aflatoxin in human male infertility in Benin City, Nigeria. Int. J. Fertil. Menopausal Stud. 1994, 39, 208–214. [Google Scholar] [PubMed]
- Martenies, S.E.; Perry, M.J. Environmental and occupational pesticide exposure and human sperm parameters: A systematic review. Toxicology 2013, 307, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Bahadur, G.; Ling, K.L.E.; Katz, M. Andrology: Statistical modelling reveals demography and time are the main contributing factors in global sperm count changes between 1938 and 1996. Hum. Reprod. 1996, 11, 2635–2639. [Google Scholar] [CrossRef] [PubMed]
- Lecoutre, S.; Petrus, P.; Ryden, M.; Breton, C. Transgenerational Epigenetic Mechanisms in Adipose Tissue Development. Trends Endocrinol. Metab. Tem 2018, 29, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, K.; Hariya, N.; Honma, K.; Goda, T. Relationship between epigenetic regulation, dietary habits, and the developmental origins of health and disease theory. Congenit. Anom. 2017, 57, 184–190. [Google Scholar] [CrossRef]
- Jobson, M.A.; Jordan, J.M.; Sandrof, M.A.; Hibshman, J.D.; Lennox, A.L.; Baugh, L.R. Transgenerational Effects of Early Life Starvation on Growth, Reproduction, and Stress Resistance in Caenorhabditis elegans. Genetics 2015, 201, 201–212. [Google Scholar] [CrossRef]
- Youssef, N.A.; Lockwood, L.; Su, S.; Hao, G.; Rutten, B.P.F. The Effects of Trauma, with or without PTSD, on the Transgenerational DNA Methylation Alterations in Human Offsprings. Brain Sci. 2018, 8, 83. [Google Scholar] [CrossRef]
- Finegersh, A.; Homanics, G.E. Paternal alcohol exposure reduces alcohol drinking and increases behavioral sensitivity to alcohol selectively in male offspring. PLoS ONE 2014, 9, e99078. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.C.; Glastonbury, C.A.; Eliot, M.N.; Bollepalli, S.; Yet, I.; Castillo-Fernandez, J.E.; Carnero-Montoro, E.; Hardiman, T.; Martin, T.C.; Vickers, A.; et al. Smoking induces coordinated DNA methylation and gene expression changes in adipose tissue with consequences for metabolic health. Clin. Epigenet. 2018, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Ben Maamar, M.; Sadler-Riggleman, I.; Beck, D.; Nilsson, E.; McBirney, M.; Klukovich, R.; Xie, Y.; Tang, C.; Yan, W. Alterations in sperm DNA methylation, non-coding RNA and histone retention associate with DDT-induced epigenetic transgenerational inheritance of disease. Epigenet. Chromatin 2018, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Rezapour, S.; Shiravand, M.; Mardani, M. Epigenetic changes due to physical activity. Biotechnol. Appl. Biochem. 2018, 65, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Donkin, I.; Barres, R. Sperm epigenetics and influence of environmental factors. Mol. Metab. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lea, A.J.; Tung, J.; Archie, E.A.; Alberts, S.C. Developmental plasticity: Bridging research in evolution and human health. Evol. Med. Public Health 2017, 2017, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Burton, A.; Torres-Padilla, M.E. Epigenetic reprogramming and development: A unique heterochromatin organization in the preimplantation mouse embryo. Brief. Funct. Genom. 2010, 9, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Sethi, G. Role of epigenetics in inflammation-associated diseases. Sub-Cell. Biochem. 2013, 61, 627–657. [Google Scholar] [CrossRef]
- Qin, Y.; Wade, P.A. Crosstalk between the microbiome and epigenome: Messages from bugs. J. Biochem. 2018, 163, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.K.; Itchon-Ramos, N.; Visco, Z.; Huang, Z.; Grenier, C.; Schrott, R.; Acharya, K.; Boudreau, M.H.; Price, T.M.; Raburn, D.J.; et al. Cannabinoid exposure and altered DNA methylation in rat and human sperm. Epigenetics 2018, 13, 1208–1221. [Google Scholar] [CrossRef] [PubMed]
- Alfano, R.; Herceg, Z.; Nawrot, T.S.; Chadeau-Hyam, M.; Ghantous, A.; Plusquin, M. The Impact of Air Pollution on Our Epigenome: How Far Is the Evidence? (A Systematic Review). Curr. Environ. Health Rep. 2018, 5, 544–578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kutateladze, T.G. Diet and the epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Huang, K.; Zhang, B.; Zhu, L.; Xu, W. Aflatoxin B1-induced epigenetic alterations: An overview. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 109, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Vargas, H.; Castelino, J.; Silver, M.J.; Dominguez-Salas, P.; Cros, M.P.; Durand, G.; Le Calvez-Kelm, F.; Prentice, A.M.; Wild, C.P.; Moore, S.E.; et al. Exposure to aflatoxin B1 in utero is associated with DNA methylation in white blood cells of infants in The Gambia. Int. J. Epidemiol. 2015, 44, 1238–1248. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Q.C.; Han, J.; Xiong, B.; Sun, S.C. Aflatoxin B1 is toxic to porcine oocyte maturation. Mutagenesis 2015, 30, 527–535. [Google Scholar] [CrossRef]
- Rieswijk, L.; Claessen, S.M.; Bekers, O.; van Herwijnen, M.; Theunissen, D.H.; Jennen, D.G.; de Kok, T.M.; Kleinjans, J.C.; van Breda, S.G. Aflatoxin B1 induces persistent epigenomic effects in primary human hepatocytes associated with hepatocellular carcinoma. Toxicology 2016, 350–352, 31–39. [Google Scholar] [CrossRef]
- Tryndyak, V.; Kindrat, I.; Dreval, K.; Churchwell, M.I.; Beland, F.A.; Pogribny, I.P. Effect of aflatoxin B1, benzo[a]pyrene, and methapyrilene on transcriptomic and epigenetic alterations in human liver HepaRG cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 121, 214–223. [Google Scholar] [CrossRef]
- Wu, H.C.; Wang, Q.; Yang, H.I.; Tsai, W.Y.; Chen, C.J.; Santella, R.M. Global DNA methylation in a population with aflatoxin B1 exposure. Epigenetics 2013, 8, 962–969. [Google Scholar] [CrossRef]
- Mezzelani, A.; Raggi, M.E.; Marabotti, A.; Milanesi, L. Ochratoxin A as possible factor trigging autism and its male prevalence via epigenetic mechanism. Nutr. Neurosci. 2016, 19, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Rasic, D.; Mladinic, M.; Zeljezic, D.; Pizent, A.; Stefanovic, S.; Milicevic, D.; Konjevoda, P.; Peraica, M. Effects of combined treatment with ochratoxin A and citrinin on oxidative damage in kidneys and liver of rats. Toxicon 2018, 146, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gan, F.; Hou, L.; Zhou, X.; Adam Ibrahim, Y.A.; Huang, K. Modulations of DNMT1 and HDAC1 are involved in the OTA-induced cytotoxicity and apoptosis in vitro. Chem.-Biol. Interact. 2017, 278, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Ansari, K.I.; Hussain, I.; Das, H.K.; Mandal, S.S. Overexpression of human histone methylase MLL1 upon exposure to a food contaminant mycotoxin, deoxynivalenol. FEBS J. 2009, 276, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef]
- Moosavi, A.; Motevalizadeh Ardekani, A. Role of Epigenetics in Biology and Human Diseases. Iran. Biomed. J. 2016, 20, 246–258. [Google Scholar] [PubMed]
- Toh, T.B.; Lim, J.J.; Chow, E.K. Epigenetics of hepatocellular carcinoma. Clin. Transl. Med. 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Pasculli, B.; Barbano, R.; Parrella, P. Epigenetics of breast cancer: Biology and clinical implication in the era of precision medicine. Semin. Cancer Biol. 2018, 51, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Wouters, B.J.; Delwel, R. Epigenetics and approaches to targeted epigenetic therapy in acute myeloid leukemia. Blood 2016, 127, 42–52. [Google Scholar] [CrossRef]
- Alelú-Paz, R.; Carmona, F.J.; Sanchez-Mut, J.V.; Cariaga-Martínez, A.; González-Corpas, A.; Ashour, N.; Orea, M.J.; Escanilla, A.; Monje, A.; Guerrero Márquez, C.; et al. Epigenetics in Schizophrenia: A Pilot Study of Global DNA Methylation in Different Brain Regions Associated with Higher Cognitive Functions. Front. Psychol. 2016, 7, 1496. [Google Scholar] [CrossRef]
- Eshraghi, A.A.; Liu, G.; Kay, S.-I.S.; Eshraghi, R.S.; Mittal, J.; Moshiree, B.; Mittal, R. Epigenetics and autism spectrum disorder: Is there a correlation? Front. Cell. Neurosci. 2018, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Pan, F.; Yang, J.; Fu, Z.; Lu, Y.; Wu, X.; Han, X.; Chen, M.; Lu, C.; Xia, Y. Idiopathic male infertility is strongly associated with aberrant DNA methylation of imprinted loci in sperm: A case-control study. Clin. Epigenet. 2018, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Dada, R.; Kumar, M.; Jesudasan, R.; Fernández, J.L.; Gosálvez, J.; Agarwal, A. Epigenetics and its role in male infertility. J. Assist. Reprod. Genet. 2012, 29, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.G.; Aston, K.I.; James, E.R.; Carrell, D.T. Sperm epigenetics in the study of male fertility, offspring health, and potential clinical applications. Syst. Biol. Reprod. Med. 2017, 63, 69–76. [Google Scholar] [CrossRef]
- Friemel, C.; Ammerpohl, O.; Gutwein, J.; Schmutzler, A.G.; Caliebe, A.; Kautza, M.; von Otte, S.; Siebert, R.; Bens, S. Array-based DNA methylation profiling in male infertility reveals allele-specific DNA methylation in PIWIL1 and PIWIL2. Fertil. Steril. 2014, 101, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Maamar, M.B.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Epigenetic transgenerational inheritance of altered sperm histone retention sites. Sci. Rep. 2018, 8, 5308. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kokkinaki, M.; Pant, D.; Gallicano, G.I.; Dym, M. Small RNA molecules in the regulation of spermatogenesis. Reproduction (Camb. Engl.) 2009, 137, 901–911. [Google Scholar] [CrossRef]
- Clermont, Y. Kinetics of spermatogenesis in mammals: Seminiferous epithelium cycle and spermatogonial renewal. Physiol. Rev. 1972, 52, 198–236. [Google Scholar] [CrossRef]
- Staub, C.; Johnson, L. Review: Spermatogenesis in the bull. Anim. Int. J. Anim. Biosci. 2018, 12, s27–s35. [Google Scholar] [CrossRef]
- Hamano, K.-I.; Sugimoto, R.; Takahashi, H.; Tsujii, H. Spermatogenesis in immature mammals. Reprod. Med. Biol. 2007, 6, 139–149. [Google Scholar] [CrossRef]
- Clermont, Y.; Huckins, C. Microscopic anatomy of the sex cords and seminiferous tubules in growing and adult male albino rats. Am. J. Anat. 1961, 108, 79–97. [Google Scholar] [CrossRef]
- Holstein, A.F.E.; Knobil, J.D. Neill (eds): The Physiology of Reproduction. Andrologia 1994, 26, 357. [Google Scholar] [CrossRef]
- Gwatkin, R.B.L. The Sertoli cell, edited by Lonnie, D. Russell and Michael, D. Griswold, Cache River Press, Clearwater, FL, 1993, 826 pp, $137.50. Mol. Reprod. Dev. 1993, 36, 517. [Google Scholar] [CrossRef]
- O’Donnell, L.; Stanton, P.; de Kretser, D.M. Endocrinology of the Male Reproductive System and Spermatogenesis. Available online: www.endotext.org (accessed on 11 January 2017).
- Neaves, W.B. A report prepared for the ford foundation review of research and support in reproductive biology and contraceptive development. Contraception 1975, 11, 571–606. [Google Scholar] [CrossRef]
- Walker, W.H. Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis 2011, 1, 116–120. [Google Scholar] [CrossRef]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef]
- Hilscher, B.; Engemann, A. Histological and morphometric studies on the kinetics of germ cells and immature Sertoli cells during human prespermatogenesis. Andrologia 1992, 24, 7–10. [Google Scholar] [CrossRef]
- Esteves, S. Male infertility due to spermatogenic failure: Current management and future perspectives. Anim. Reprod. 2015, 12, 62–80. [Google Scholar]
- Plant, T.M. The hypothalamo-pituitary-gonadal axis. J. Endocrinol. 2015, 226, T41–T45. [Google Scholar] [CrossRef]
- Wilson, R.C.; Kesner, J.S.; Kaufman, J.-M.; Uemura, T.; Akema, T.; Knobil, E. Central electrophysiologic correlates of pulsatile luteinizing hormone secretion in the rhesus monkey. Neuroendocrinology 1984, 39, 256–260. [Google Scholar] [CrossRef]
- Majumdar, S.; Mikuma, N.; Ishwad, P.; Winters, S.; Attardi, B.; Perera, A.; Plant, T. Replacement with recombinant human inhibin immediately after orchidectomy in the hypophysiotropically clamped male rhesus monkey (Macaca mulatta) maintains follicle-stimulating hormone (FSH) secretion and FSH beta messenger ribonucleic acid levels at precastration values. Endocrinology 1995, 136, 1969–1977. [Google Scholar] [PubMed]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]
- Horstman, A.M.; Dillon, E.L.; Urban, R.J.; Sheffield-Moore, M. The role of androgens and estrogens on healthy aging and longevity. J. Gerontol. Ser. A Biomed. Sci. Med Sci. 2012, 67, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Halkerston, I.; Eichhorn, J.; Hechter, O. A requirement for reduced triphosphopyridine nucleotide for cholesterol side-chain cleavage by mitochondrial fractions of bovine adrenal cortex. J. Biol. Chem. 1961, 236, 374–380. [Google Scholar] [PubMed]
- Tuckey, R.C.; Cameron, K.J. Side-chain specificities of human and bovine cytochromes P-450scc. Eur. J. Biochem. 1993, 217, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.L.; Myers, R.P.; Strickler, R.C. Human placental 3 beta-hydroxy-5-ene-steroid dehydrogenase and steroid 5 → 4-ene-isomerase: Purification from mitochondria and kinetic profiles, biophysical characterization of the purified mitochondrial and microsomal enzymes. J. Steroid Biochem. 1989, 33, 209–217. [Google Scholar] [CrossRef]
- Lachance, Y.; Labrie, C.; Simard, J.; Dumont, M.; De Launoit, Y.; Guérin, S.; Leblanc, G.; Labrie, F. Characterization of human 3 beta-hydroxysteroid dehydrogenase/delta 5-delta 4-isomerase gene and its expression in mammalian cells. J. Biol. Chem. 1990, 265, 20469–20475. [Google Scholar] [PubMed]
- Lorence, M.C.; MURRY, B.A.; TRANT, J.M.; MASON, J.I. Human 3β-hydroxysteroid dehydrogenase/Δ5 → 4isomerase from placenta: Expression in nonsteroidogenic cells of a protein that catalyzes the dehydrogenation/isomerization of C21 and C19 steroids. Endocrinology 1990, 126, 2493–2498. [Google Scholar] [CrossRef]
- Lee, E.; Starcevic, S.; Catlin, D. Effects of dietary supplements, 19-norandrostenedione, androstenediol and androstenedione on the profile of urine steroids. In Journal of Investigative Medicine; Slack Inc.: Thorofare, NJ, USA, 1999; p. 62A. [Google Scholar]
- Simpson, E.; Mahendroo, M.; Nichols, J.; Bulun, S. Aromatase gene expression in adipose tissue: Relationship to breast cancer. Int. J. Fertil. Menopausal Stud. 1994, 39, 75–83. [Google Scholar]
- Moore, R.J.; Wilson, J.D. Steroid 5alpha-reductase in cultured human fibroblasts. Biochemical and genetic evidence for two distinct enzyme activities. J. Biol. Chem. 1976, 251, 5895–5900. [Google Scholar]
- Gharbi, A.; Trillon, O.; Betbeder, A.M.; Counord, J.; Gauret, M.F.; Pfohl-Leszkowicz, A.; Dirheimer, G.; Creppy, E.E. Some effects of ochratoxin A, a mycotoxin contaminating feeds and food, on rat testis. Toxicology 1993, 83, 9–18. [Google Scholar] [CrossRef]
- Park, H.; Park, H.S.; Lim, W.; Song, G. Ochratoxin A suppresses proliferation of Sertoli and Leydig cells in mice. Med. Mycol. 2019. [Google Scholar] [CrossRef]
- Zatecka, E.; Ded, L.; Elzeinova, F.; Kubatova, A.; Dorosh, A.; Margaryan, H.; Dostalova, P.; Korenkova, V.; Hoskova, K.; Peknicova, J. Effect of zearalenone on reproductive parameters and expression of selected testicular genes in mice. Reprod. Toxicol. (Elmsford N.Y.) 2014, 45, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Netro, H.M.; Chorfi, Y.; Price, C.A. Effects of the mycotoxin deoxynivalenol on steroidogenesis and apoptosis in granulosa cells. Reproduction (Camb. Engl.) 2015, 149, 555–561. [Google Scholar] [CrossRef]
- Gupta, M.; Bandyopadhyay, S.; Paul, B.; Mazumdar, S.K. Ovarian steroidogenesis and development of fetuses following ochratoxin A treatment in pregnant rats. Endokrinologie 1981, 77, 152–160. [Google Scholar] [PubMed]
- Long, G.G.; Diekman, M.A. Effect of purified zearalenone on early gestation in gilts. J. Anim. Sci. 1984, 59, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Parandin, R.; Behnam-Rassouli, M.; Mahdavi-Shahri, N. Effects of Neonatal Exposure to Zearalenone on Puberty Timing, Hypothalamic Nuclei of AVPV and ARC, and Reproductive Functions in Female Mice. Reprod. Sci. (Thousand Oaks Calif.) 2017, 24, 1293–1303. [Google Scholar] [CrossRef]
- Yegani, M.; Chowdhury, S.R.; Oinas, N.; MacDonald, E.J.; Smith, T.K. Effects of feeding grains naturally contaminated with Fusarium mycotoxins on brain regional neurochemistry of laying hens, turkey poults, and broiler breeder hens. Poult. Sci. 2006, 85, 2117–2123. [Google Scholar] [CrossRef] [PubMed]
- Albonico, M.; Schutz, L.F.; Caloni, F.; Cortinovis, C.; Spicer, L.J. In vitro effects of the Fusarium mycotoxins fumonisin b1 and beauvericin on bovine granulosa cell proliferation and steroid production. Toxicon 2017, 128, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium Molds and Mycotoxins: Potential Species-Specific Effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Caloni, F.; Schreiber, N.B.; Spicer, L.J. Effects of fumonisin B1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology 2014, 81, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Ranzenigo, G.; Caloni, F.; Cremonesi, F.; Aad, P.Y.; Spicer, L.J. Effects of Fusarium mycotoxins on steroid production by porcine granulosa cells. Anim. Reprod. Sci. 2008, 107, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Spensley, P. Aflatoxin, the active principle in turkey‘X’disease. Endeavour 1963, 22, 75–79. [Google Scholar] [CrossRef]
- Sprando, R.L.; Collins, T.F.; Black, T.N.; Olejnik, N.; Rorie, J.I.; Eppley, R.M.; Ruggles, D.I. Characterization of the effect of deoxynivalenol on selected male reproductive endpoints. Food Chem. Toxicol. 2005, 43, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Sprando, R.; Pestka, J.; Collins, T.; Rorie, J.; O’donnell, M.; Hinton, D.; Chirtel, S. The Effect of Vomitoxin (Deoxnivalenol) on Testicular Morphology, Testicular Spermatid Counts and Epididymal Sperm Counts in IL-6KO [B6129-IL6 < tmlKopf > (IL-6 gene deficient)] and WT [B6129F2 (wild type to B6129-IL6 with an intact IL-6 gene)] mice. Food Chem. Toxicol. 1999, 37, 1073–1079. [Google Scholar] [CrossRef]
- Han, J.; Wang, Q.-C.; Zhu, C.-C.; Liu, J.; Zhang, Y.; Cui, X.-S.; Kim, N.-H.; Sun, S.-C. Deoxynivalenol exposure induces autophagy/apoptosis and epigenetic modification changes during porcine oocyte maturation. Toxicol. Appl. Pharmacol. 2016, 300, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, U.; Tomek, W.; Schneider, F.; Vanselow, J. Effects of the mycotoxins α-and β-zearalenol on regulation of progesterone synthesis in cultured granulosa cells from porcine ovaries. Reprod. Toxicol. 2003, 17, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Frizzell, C.; Shannon, M.; Ruiz, M.-J.; Connolly, L. An in vitro investigation on the cytotoxic and nuclear receptor transcriptional activity of the mycotoxins fumonisin B1 and beauvericin. Toxicol. Lett. 2016, 257, 1–10. [Google Scholar] [CrossRef]
- Behm, C.; Föllmann, W.; Degen, G.H. Cytotoxic potency of mycotoxins in cultures of V79 lung fibroblast cells. J. Toxicol. Environ. Health Part A 2012, 75, 1226–1231. [Google Scholar] [CrossRef]
- Mallebrera, B.; Brandolini, V.; Font, G.; Ruiz, M. Cytoprotective effect of resveratrol diastereomers in CHO-K1 cells exposed to beauvericin. Food Chem. Toxicol. 2015, 80, 319–327. [Google Scholar] [CrossRef]
- Schoevers, E.J.; Santos, R.R.; Fink-Gremmels, J.; Roelen, B.A. Toxicity of beauvericin on porcine oocyte maturation and preimplantation embryo development. Reprod. Toxicol. 2016, 65, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Aydin, Y.; Orta Yilmaz, B.; Yildizbayrak, N.; Korkut, A.; Arabul Kursun, M.; Irez, T.; Erkan, M. Evaluation of citrinin-induced toxic effects on mouse Sertoli cells. Drug Chem. Toxicol. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ewuola, E.O. Organ traits and histopathology of rabbits fed varied levels of dietary fumonisin B(1). J. Anim. Physiol. Anim. Nutr. 2009, 93, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Karacaoglu, E.; Selmanoglu, G. T-2 toxin induces cytotoxicity and disrupts tight junction barrier in SerW3 cells. Environ. Toxicol. Pharmacol. 2017, 56, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Koraïchi, F.; Inoubli, L.; Lakhdari, N.; Meunier, L.; Vega, A.; Mauduit, C.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Neonatal exposure to zearalenone induces long term modulation of ABC transporter expression in testis. Toxicology 2013, 310, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Pan, S.; Wang, G.; Wang, Y.J.; Liu, Q.; Gu, J.; Yuan, Y.; Liu, X.Z.; Liu, Z.P.; Bian, J.C. Zearalenone impairs the male reproductive system functions via inducing structural and functional alterations of sertoli cells. Environ. Toxicol. Pharmacol. 2016, 42, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.L.; Hu, J.; Guo, B.P.; Niu, Y.R.; Xiao, C.; Xu, Y.X. Exploration of intrinsic and extrinsic apoptotic pathways in zearalenone-treated rat sertoli cells. Environ. Toxicol. 2016, 31, 1731–1739. [Google Scholar] [CrossRef]
- Cai, G.; Si, M.; Li, X.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis of rat Sertoli cells through Fas-Fas ligand and mitochondrial pathway. Environ. Toxicol. 2019, 34, 424–433. [Google Scholar] [CrossRef]
- Milano, G.D.; Becu-Villalobos, D.; Tapia, M.O. Effects of long-term zearalenone administration on spermatogenesis and serum luteinizing hormone, follicle-stimulating hormone, and prolactin values in male rats. Am. J. Vet. Res. 1995, 56, 954–958. [Google Scholar]
- Filipiak, E.; Walczak-Jedrzejowska, R.; Oszukowska, E.; Guminska, A.; Marchlewska, K.; Kula, K.; Slowikowska-Hilczer, J. Xenoestrogens diethylstilbestrol and zearalenone negatively influence pubertal rat’s testis. Folia Histochem. Cytobiol. 2009, 47, S113–S120. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, R.Y.; Zhao, Y.; Xu, C.S.; Zhang, W.D.; Ge, W.; Liu, J.; Sun, Z.Y.; Zou, S.H.; Shen, W. Ochratoxin A exposure decreased sperm motility via the AMPK and PTEN signaling pathways. Toxicol. Appl. Pharm. 2018, 340, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yang, S.H.; Shi, W.; Li, P.; Guo, Y.; Guo, J.; He, J.B.; Zhang, Y. Protective effect of proanthocyanidin on mice Sertoli cell apoptosis induced by zearalenone via the Nrf2/ARE signalling pathway. Environ. Sci. Pollut. Res. Int. 2017, 24, 26724–26733. [Google Scholar] [CrossRef] [PubMed]
- Adedara, I.A.; Nanjappa, M.K.; Farombi, E.O.; Akingbemi, B.T. Aflatoxin B1 disrupts the androgen biosynthetic pathway in rat Leydig cells. Food Chem. Toxicol. 2014, 65, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.; Cockrum, R.; Jons, A.; Alexander, B.; Cammack, K. Renin mRNA is upregulated in testes and testicular cells in response to treatment with aflatoxin B1. Theriogenology 2012, 77, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, D.; Zhang, J.; Zhang, D.; Gong, M.; Wang, C.; Wei, N.; Liu, W.; Wang, Y.; Zhao, C. Citrinin reduces testosterone secretion by inducing apoptosis in rat Leydig cells. Toxicol. In Vitro 2012, 26, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y.; Jing, A.; Ma, K.; Gong, Q.; Qin, C. Effects of T-2 toxin on testosterone biosynthesis in mouse Leydig cells. Toxicol. Ind. Health 2014, 30, 873–877. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Wang, Y.; Cui, S. Toxic effects of zearalenone and α-zearalenol on the regulation of steroidogenesis and testosterone production in mouse Leydig cells. Toxicol. In Vitro 2007, 21, 558–565. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Gu, J.; Yuan, Y.; Liu, X.; Zheng, W.; Huang, Q.; Liu, Z.; Bian, J. Zearalenone inhibits testosterone biosynthesis in mouse Leydig cells via the crosstalk of estrogen receptor signaling and orphan nuclear receptor Nur77 expression. Toxicol. In Vitro 2014, 28, 647–656. [Google Scholar] [CrossRef]
- Eze, U.; Routledge, M.; Okonofua, F.; Huntriss, J.; Gong, Y. Mycotoxin exposure and adverse reproductive health outcomes in Africa: A review. World Mycotoxin J. 2018, 11, 321–339. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Huang, K.; He, X.; Luo, Y.; Liang, R.; Luo, H.; Shen, X.L.; Xu, W. Mitochondrial proteomic analysis reveals the molecular mechanisms underlying reproductive toxicity of zearalenone in MLTC-1 cells. Toxicology 2014, 324, 55–67. [Google Scholar] [CrossRef]
- Lin, P.; Chen, F.; Sun, J.; Zhou, J.; Wang, X.; Wang, N.; Li, X.; Zhang, Z.; Wang, A.; Jin, Y. Mycotoxin zearalenone induces apoptosis in mouse Leydig cells via an endoplasmic reticulum stress-dependent signalling pathway. Reprod. Toxicol. (Elmsford N.Y.) 2015, 52, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary Leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Tegelenbosch, R.A.; de Rooij, D.G. A quantitative study of spermatogonial multiplication and stem cell renewal in the C3H/101 F1 hybrid mouse. Mutat. Res. 1993, 290, 193–200. [Google Scholar] [CrossRef]
- Kuiper-Goodman, T.; Scott, P.M.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharmacol. 1987, 7, 253–306. [Google Scholar] [CrossRef]
- Qingqing, H.; Linbo, Y.; Yunqian, G.; Shuqiang, L. Toxic effects of citrinin on the male reproductive system in mice. Exp. Toxicol. Pathol. 2012, 64, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Supriya, C.; Girish, B.P.; Reddy, P.S. Aflatoxin B1-Induced Reproductive Toxicity in Male Rats: Possible Mechanism of Action. Int. J. Toxicol. 2014, 33, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Hasanzadeh, S.; Rezazadeh, L. Effects of aflatoxin B1 on the growth processes of spermatogenic cell series in adult male rats. Comp. Clin. Pathol. 2012, 22, 555–562. [Google Scholar] [CrossRef]
- Gbore, F.A. Reproductive organ weights and semen quality of pubertal boars fed dietary fumonisin B1. Anim. Int. J. Anim. Biosci. 2009, 3, 1133–1137. [Google Scholar] [CrossRef]
- Gbore, F.A.; Egbunike, G.N. Testicular and epididymal sperm reserves and sperm production of pubertal boars fed dietary fumonisin B(1). Anim. Reprod. Sci. 2008, 105, 392–397. [Google Scholar] [CrossRef]
- Ewuola, E.O.; Egbunike, G.N. Effects of dietary fumonisin B1 on the onset of puberty, semen quality, fertility rates and testicular morphology in male rabbits. Reproduction (Camb. Engl.) 2010, 139, 439–445. [Google Scholar] [CrossRef]
- Selmanoglu, G. Evaluation of the reproductive toxicity of patulin in growing male rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2006, 44, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Zhang, Y.F.; Liang, A.M.; Kong, X.F.; Li, Y.X.; Ma, K.W.; Jing, A.H.; Feng, S.Y.; Qiao, X.L. Toxic effects of T-2 toxin on reproductive system in male mice. Toxicol. Ind. Health 2010, 26, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.H.; Son, H.Y.; Cho, S.W.; Ha, C.S.; Kang, B.H. Zearalenone induces male germ cell apoptosis in rats. Toxicol. Lett. 2003, 138, 185–192. [Google Scholar] [CrossRef]
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, mechanisms and clinical perspective. In Seminars in Reproductive Medicine; Thieme Medical Publishers: Stuttgart, Germany, 2009; pp. 351–357. [Google Scholar]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Nery, J.R.; Lee, L.; Ye, Z.; Ngo, Q.-M. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315. [Google Scholar] [CrossRef] [PubMed]
- Alhamwe, B.A.; Khalaila, R.; Wolf, J.; von Bülow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H. Histone modifications and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, R.; Lindquist, S. Epigenetics in the extreme: Prions and the inheritance of environmentally acquired traits. Science 2010, 330, 629–632. [Google Scholar] [CrossRef]
- Becker, P.B.; Workman, J.L. Nucleosome remodeling and epigenetics. Cold Spring Harb. Perspect. Biol. 2013, 5, a017905. [Google Scholar] [CrossRef] [PubMed]
- Suelves, M.; Carrió, E.; Núñez-Álvarez, Y.; Peinado, M.A. DNA methylation dynamics in cellular commitment and differentiation. Brief. Funct. Genom. 2016, 15, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Chen, T. DNA methylation reprogramming during mammalian development. Genes 2019, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Shin, W.; Lee, J.; Do, J. CpG and non-CpG methylation in epigenetic gene regulation and brain function. Genes 2017, 8, 148. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, D.; Avvakumov, N.; Côté, J. Histone phosphorylation: A chromatin modification involved in diverse nuclear events. Epigenetics 2012, 7, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.B.; Morgan, C.P.; Bronson, S.L.; Revello, S.; Bale, T.L. Paternal stress exposure alters sperm microRNA content and reprograms offspring HPA stress axis regulation. J. Neurosci. 2013, 33, 9003–9012. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Saldanha, S.N.; Tollefsbol, T.O. Impact of epigenetic dietary compounds on transgenerational prevention of human diseases. AAPS J. 2014, 16, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, B.; Karlic, H.; Varga, F.; Fabianowska-Majewska, K.; Haslberger, A. Epigenetic mechanisms in anti-cancer actions of bioactive food components—The implications in cancer prevention. Br. J. Pharmacol. 2012, 167, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Ahmed, A.; Tollefsbol, T.O. Epigenetic targets of bioactive dietary components for cancer prevention and therapy. Clin. Epigenet. 2010, 1, 101. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T. Epigenetic alterations induced by environmental stress associated with metabolic and neurodevelopmental disorders. Environ. Epigenet. 2016, 2, dvw017. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.G.; Morgan, M.D.; Heger, A.H.; Sharpe, R.M.; Drake, A.J. High-fat diet disrupts metabolism in two generations of rats in a parent-of-origin specific manner. Sci. Rep. 2016, 6, 31857. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.; King, S.E.; McBirney, M.; Kubsad, D.; Pappalardo, M.; Beck, D.; Sadler-Riggleman, I.; Skinner, M.K. Vinclozolin induced epigenetic transgenerational inheritance of pathologies and sperm epimutation biomarkers for specific diseases. PLoS ONE 2018, 13, e0202662. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Anway, M.D. Epigenetic transgenerational actions of vinclozolin on the development of disease and cancer. Crit. Rev. Oncog. 2007, 13, 75–82. [Google Scholar] [CrossRef]
- Anway, M.D.; Leathers, C.; Skinner, M.K. Endocrine disruptor vinclozolin induced epigenetic transgenerational adult-onset disease. Endocrinology 2006, 147, 5515–5523. [Google Scholar] [CrossRef] [PubMed]
- Lacal, I.; Ventura, R. Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Ly, L.; Chan, D.; Trasler, J.M. Developmental windows of susceptibility for epigenetic inheritance through the male germline. Semin. Cell Dev. Biol. 2015, 43, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Endocrinol. Metab. Tem 2010, 21, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, E.; Lamb, M.J. The inheritance of acquired epigenetic variations. Int. J. Epidemiol. 2015, 44, 1103–1105. [Google Scholar] [CrossRef] [PubMed]
- Sloane, M.A.; Nunez, A.C.; Packham, D.; Kwok, C.T.; Suthers, G.; Hesson, L.B.; Ward, R.L. Mosaic Epigenetic Inheritance as a Cause of Early-Onset Colorectal Cancer. JAMA Oncol. 2015, 1, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.I.; Ward, R.; Suter, C.M. Germline epimutation: A basis for epigenetic disease in humans. Ann. N. Y. Acad. Sci. 2005, 1054, 68–77. [Google Scholar] [CrossRef]
- Ge, S.-Q.; Lin, S.-L.; Zhao, Z.-H.; Sun, Q.-Y. Epigenetic dynamics and interplay during spermatogenesis and embryogenesis: Implications for male fertility and offspring health. Oncotarget 2017, 8, 53804–53818. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Kovalchuk, I. Epigenetic memory in mammals. Front. Genet. 2011, 2, 28. [Google Scholar] [CrossRef]
- Miyoshi, N.; Stel, J.M.; Shioda, K.; Qu, N.; Odajima, J.; Mitsunaga, S.; Zhang, X.; Nagano, M.; Hochedlinger, K.; Isselbacher, K.J.; et al. Erasure of DNA methylation, genomic imprints, and epimutations in a primordial germ-cell model derived from mouse pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2016, 113, 9545–9550. [Google Scholar] [CrossRef]
- Seisenberger, S.; Peat, J.R.; Hore, T.A.; Santos, F.; Dean, W.; Reik, W. Reprogramming DNA methylation in the mammalian life cycle: Building and breaking epigenetic barriers. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20110330. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Dietmann, S.; Irie, N.; Leitch, H.G.; Floros, V.I.; Bradshaw, C.R.; Hackett, J.A.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Berletch, J.B.; Yang, F.; Xu, J.; Carrel, L.; Disteche, C.M. Genes that escape from X inactivation. Hum. Genet. 2011, 130, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef] [PubMed]
- Nassan, F.L.; Arvizu, M.; Minguez-Alarcon, L.; Williams, P.L.; Attaman, J.; Petrozza, J.; Hauser, R.; Chavarro, J. Marijuana smoking and markers of testicular function among men from a fertility centre. Hum. Reprod. (Oxf. Engl.) 2019, 34, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Sauria, M.E.G.; Lyu, X.; Cheema, M.S.; Ausio, J.; Taylor, J.; Corces, V.G. Chromatin States in Mouse Sperm Correlate with Embryonic and Adult Regulatory Landscapes. Cell Rep. 2017, 18, 1366–1382. [Google Scholar] [CrossRef] [PubMed]
- Champroux, A.; Cocquet, J.; Henry-Berger, J.; Drevet, J.R.; Kocer, A. A Decade of Exploring the Mammalian Sperm Epigenome: Paternal Epigenetic and Transgenerational Inheritance. Front. Cell Dev. Biol. 2018, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Bohacek, J.; Mansuy, I.M. Molecular insights into transgenerational non-genetic inheritance of acquired behaviours. Nat. Rev. Genet. 2015, 16, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Singh, A.K. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef]
- Kosan, C.; Heidel, F.H.; Godmann, M.; Bierhoff, H. Epigenetic Erosion in Adult Stem Cells: Drivers and Passengers of Aging. Cells 2018, 7, 237. [Google Scholar] [CrossRef]
- Shnorhavorian, M.M.; Schwartz, S.; Stansfeld, B.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Differential DNA Methylation Regions in Adult Human Sperm following Adolescent Chemotherapy: Potential for Epigenetic Inheritance. PLoS ONE 2017, 12, e0170085. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shinohara, T. Epigenetic modifications and self-renewal regulation of mouse germline stem cells. Cell Res. 2011, 21, 1164–1171. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sun, Y.C.; Wang, Y.Y.; Ge, W.; Cheng, S.F.; Dyce, P.W.; Shen, W. Epigenetic regulation during the differentiation of stem cells to germ cells. Oncotarget 2017, 8, 57836–57844. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, E. DNA Methylation Regulates Genomic Imprinting, X Inactivation, and Gene Expression during Mammalian Development. In Gene Expression and Regulation; Ma, J., Ed.; Springer New York: New York, NY, USA, 2006; pp. 377–391. [Google Scholar]
- Barlow, D.P.; Bartolomei, M.S. Genomic imprinting in mammals. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Ferguson-Smith, A.C.; Bourc’his, D. The discovery and importance of genomic imprinting. Elife 2018, 7, e42368. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.J.; Costa, P.; Vaz, B.; Carvalho, F.; Fernandes, S.; Barros, A.; Sousa, M. Abnormal methylation of imprinted genes in human sperm is associated with oligozoospermia. Mol. Hum. Reprod. 2008, 14, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Lucifero, D.; Chaillet, J.R.; Trasler, J.M. Potential significance of genomic imprinting defects for reproduction and assisted reproductive technology. Hum. Reprod. Update 2004, 10, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Nowak, K.; Stein, G.; Powell, E.; He, L.M.; Naik, S.; Morris, J.; Marlow, S.; Davis, T.L. Establishment of paternal allele-specific DNA methylation at the imprinted mouse Gtl2 locus. Epigenetics 2011, 6, 1012–1020. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Court, F.; Tayama, C.; Romanelli, V.; Martin-Trujillo, A.; Iglesias-Platas, I.; Okamura, K.; Sugahara, N.; Simon, C.; Moore, H.; Harness, J.V.; et al. Genome-wide parent-of-origin DNA methylation analysis reveals the intricacies of human imprinting and suggests a germline methylation-independent mechanism of establishment. Genome Res. 2014, 24, 554–569. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bretz, C.; Lee, S. Epigenetic instability of imprinted genes in human cancers. Nucleic Acids Res. 2015, 43, 10689–10699. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C. The epigenetics of embryo development. Anim. Front. 2015, 5, 42–49. [Google Scholar] [CrossRef]
- Boissonnas, C.C.; Jouannet, P.; Jammes, H. Epigenetic disorders and male subfertility. Fertil. Steril. 2013, 99, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhao, P.; Liu, J.; Zhang, J.; Zhang, J.; Wang, Y.; Wu, L.; Song, M.; Wang, W. Novel Epigenomic Biomarkers of Male Infertility Identified by Methylation Patterns of CpG Sites Within Imprinting Control Regions of H19 and SNRPN Genes. Omics A J. Integr. Biol. 2018, 22, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Selvatici, R.; Di Domenico, M.; Marci, R.; Vesce, F.; Tognon, M.; Martini, F. Methylation loss at H19 imprinted gene correlates with methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples from infertile males. Epigenetics 2013, 8, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, J.B.; Xiao, X.F.; Ma, Y.F.; Wang, J.; Liang, X.X.; Zhao, H.X.; Jiang, F.; Yao, Y.Q.; Wang, X.H. Altered DNA methylation patterns of the H19 differentially methylated region and the DAZL gene promoter are associated with defective human sperm. PLoS ONE 2013, 8, e71215. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Wang, Y.; Zou, Z.; Chen, L.; Shen, C.; Xu, S.; Zhang, J.; Zhao, F.; Ge, S.; Gao, Q.; et al. Abnormal Methylation of Imprinted Genes and Cigarette Smoking: Assessment of Their Association With the Risk of Male Infertility. Reprod. Sci. (Thousand Oaks Calif.) 2017, 24, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhou, X.; Li, D.K.; Yang, F.; Pan, H.; Li, T.; Miao, M.; Li, R.; Yuan, W. Genome-wide alteration in DNA hydroxymethylation in the sperm from bisphenol A-exposed men. PLoS ONE 2017, 12, e0178535. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Riley, R.T.; Sharma, R.P. Pharmacological antagonism of fumonisin B1 cytotoxicity in porcine renal epithelial cells (LLC-PK1): A model for reducing fumonisin-induced nephrotoxicity in vivo. Pharmacol. Toxicol. 2002, 90, 268–277. [Google Scholar] [CrossRef]
- Monge Mdel, P.; Magnoli, A.P.; Bergesio, M.V.; Tancredi, N.; Magnoli, C.E.; Chiacchiera, S.M. Activated carbons as potentially useful non-nutritive additives to prevent the effect of fumonisin B1 on sodium bentonite activity against chronic aflatoxicosis. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 1043–1052. [Google Scholar] [CrossRef]
- Filannino, A.; Stout, T.A.; Gadella, B.M.; Sostaric, E.; Pizzi, F.; Colenbrander, B.; Dell’Aquila, M.E.; Minervini, F. Dose-response effects of estrogenic mycotoxins (zearalenone, alpha- and beta-zearalenol) on motility, hyperactivation and the acrosome reaction of stallion sperm. Reprod. Biol. Endocrinol. 2011, 9, 134. [Google Scholar] [CrossRef]
- Rajkovic, A.; Uyttendaele, M.; Debevere, J. Computer aided boar semen motility analysis for cereulide detection in different food matrices. Int. J. Food Microbiol. 2007, 114, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, A.G. Spermatogonial stem cells as a therapeutic alternative for fertility preservation of prepubertal boys. Einstein 2015, 13, 637–639. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Altman, E.; Yango, P.; Moustafa, R.; Smith, J.F.; Klatsky, P.C.; Tran, N.D. Characterization of human spermatogonial stem cell markers in fetal, pediatric, and adult testicular tissues. Reproduction (Camb. Engl.) 2014, 148, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cairns, B.R. Isolation and Enrichment of Spermatogonial Stem Cells From Human Testis Tissues. Curr. Protoc. Stem Cell Biol. 2019, 49, e77. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kokkinaki, M.; Jiang, J.; Zeng, W.; Dobrinski, I.; Dym, M. Isolation of human male germ-line stem cells using enzymatic digestion and magnetic-activated cell sorting. Methods Mol. Biol. 2012, 825, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Kossack, N.; Meneses, J.; Shefi, S.; Nguyen, H.N.; Chavez, S.; Nicholas, C.; Gromoll, J.; Turek, P.J.; Reijo-Pera, R.A. Isolation and characterization of pluripotent human spermatogonial stem cell-derived cells. Stem Cells 2009, 27, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.J.; Sung, S.Y.; Kim, H.J.; Song, S.H.; Hong, J.Y.; Yoon, T.K.; Kim, J.K.; Kim, K.S.; Lee, D.R. Long-term proliferation and characterization of human spermatogonial stem cells obtained from obstructive and non-obstructive azoospermia under exogenous feeder-free culture conditions. Cell Prolif. 2010, 43, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tang, Z.; Xiong, T.; Tang, W. Isolation and characterization of human spermatogonial stem cells. Reprod. Biol. Endocrinol. 2011, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.; Stockl, J.B.; Flenkenthaler, F.; Dietrich, K.G.; Schwarzer, J.U.; Kohn, F.M.; Drummer, C.; Frohlich, T.; Arnold, G.J.; Behr, R.; et al. Characterization of a non-human primate model for the study of testicular peritubular cells-comparison with human testicular peritubular cells. Mol. Hum. Reprod. 2018, 24, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Y.; Qu, R.; He, Y.; Tian, X.; Zeng, W. Spermatogonial stem cells from domestic animals: Progress and prospects. Reproduction (Camb. Engl.) 2014, 147, R65–R74. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, L.; Sun, M.; Hai, Y.; Li, Z.; He, Z. Expansion and long-term culture of human spermatogonial stem cells via the activation of SMAD3 and AKT pathways. Exp. Biol. Med. 2015, 240, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Hai, Y.; Gong, Y.; Li, Z.; He, Z. Characterization, isolation, and culture of mouse and human spermatogonial stem cells. J. Cell. Physiol. 2014, 229, 407–413. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kokkinaki, M.; Jiang, J.; Dobrinski, I.; Dym, M. Isolation, characterization, and culture of human spermatogonia. Biol. Reprod. 2010, 82, 363–372. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Mycotoxin | Species | Dose | Exposure | Main Findings with Respect to Sertoli Cells | Ref. |
|---|---|---|---|---|---|
| CTN | Mouse | 0–200 μM | 6–72 h | Decreased cell viability and proliferation Increased apoptosis, and necrosis in a dose-dependent manner | [114] |
| DON | Mice | 10 ppm | 90 days | No effect on relative testis weight and testicular spermatid counts No effect on the number of Sertoli cells in the seminiferous tubules | [107] |
| FB1 | Rabbit | 0.13–10 mg/kg diet * | 196 days | Degeneration of Sertoli cell | [115] |
| OTA | Mice TM4 | 0–5 μM | 24 h | Decreased proliferation Dose-dependent phosphorylation of PI3K (Akt, P70S6K, and S6) and MAPK (ERK1/2 and JNK) pathways | [94] |
| T-2 | SerW3 cells | 0.012–1.2 μg/mL (0.025–25.72 μM) | 24–48 h | Increased cytotoxicity in a dose-dependent manner Targets blood-testis barrier in vitro | [116] |
| ZEA | Rat | 0–10 nM | 48 h | Negatively influenced spermatogenesis and male fertility ZEA effect inhibited by in vitro addition of anti-estrogen (ICI 182.780) → ZEA estrogenic activity | [117] |
| 0–20 g/mL | 24 h | Damages the cytoskeletal structure Disrupts specific secretory functions | [118] | ||
| 0–200 μM | 6–36 h | Induces apoptosis and necrosis via extrinsic and intrinsic apoptotic pathways | [119] | ||
| 0–20 μmol/L (0–62.3 μM) | Induces apoptosis Activates the Fas-Fas ligand signaling pathway Regulates mitochondrial apoptosis pathway | [120] | |||
| 20 mg/kg BW * | 5 weeks | Increased serum prolactin No effect on testis weights, serum luteinizing hormone, and follicle-stimulating hormone | [121] | ||
| 4 or 40 μg | 16 days | Weak estrogen effect on Sertoli cell development in pre-pubertal rats | [122] | ||
| Mice TM4 | 0–100 μM | 24 h | TM4 cell cycle G2/M arrest Apoptosis through ROS- and ER-stress and the ATP/AMPK pathway | [123] |
| Mycotoxin | Species | Dose | Exposure | Main Findings with Respect to Leydig Cells | Ref. |
|---|---|---|---|---|---|
| AFB1 | Mouse | 50 μg/kg BW * | 45 days | Upregulation of genes involved in cell differentiation, extracellular space, and immunity | [126] |
| Rat | 0–10 μM | 35 days | Extra-hepatic toxicity by inhibition of proteins involved in androgen biosynthesis such as StAR, HSDB3, and HSD17B3 | [125] | |
| CTN | 50 and 100 μM | 36 h | Reduced testosterone secretion Induced apoptosis | [127] | |
| T-2 | Mouse | 1–102 μM | 24 h | Dose-dependent decrease in testosterone levels | [128] |
| ZEA | 0–20 μg/mL (0–62.3 μM) | 1–24 h | Dose- and time-dependent inhibition of testosterone stimulated by both hCG and cAMP | [130] | |
| 0.01–100 μM | 24 h | Suppressed hCG-induced testosterone secretion | [129] | ||
| 5 μM | 24 h | Modified mitochondrial lipid metabolism Increased energy production Inhibited steroidogenesis and esterification | [132] | ||
| 0–200 μg/mL (0–623 μM) | 24 h | ER stress pathway activated in ZEA-induced apoptosis | [133] | ||
| Rat | 2.5–20 μg/mL (7.8–62.3 μM) | 12 h | Investigation of anti-ZEA compounds | [134] |
| Toxin | Species | Exposure | Daily Dose * | Effect on Spermatogenesis | Ref. |
|---|---|---|---|---|---|
| AFB1 | Rats | 60 days | 10–50 µg | Reduction of reproductive organ weights and sperm quantity and quality Decreased steroidogenesis | [138] |
| 48 days | 0.8–3.2 ppm | Dose-dependent decrease of developing spermatozoa in seminiferous tubules | [139] | ||
| CTN | Mice | 7 days | 0.0625–6.25 mg | Increased abnormal spermatozoa Decreased live spermatozoa number and count, and serum testosterone | [137] |
| DON | Rats | 28 days | 0.5–5 mg | Decreased testicular spermatid numbers Increased germ cell degeneration, sperm retention, and abnormal nuclear morphology | [106] |
| FB1 | Pigs | 6 months | 0.2–15 mg | Reduced testicular and epididymal sperm reserves Reduced daily sperm production No influence on the relative weights and volume of the testes or epididymis | [140,141] |
| Rabbits | 175 days | 0.13–10 mg | Delayed puberty, impaired semen quality and spermatogenesis, and induced embryo mortality | [142] | |
| OTA | Rats | 8 weeks | 289 µg | Decrease in stages I and VII germ cells Increase in stages XII and XIII germ cells | [93] |
| Patulin | Rats | 60–90 days | 0.1 mg | Increased sperm counts Decreased sperm counts | [143] |
| T-2 | Mice | 7 days | 0–15 mg | Increased abnormal spermatozoa Decreased testicular and cauda epididymal sperm counts, efficiency of sperm production, and serum testosterone concentrations | [144] |
| ZEA | Rats | 48 h | 5 mg | Germ cell degeneration, especially spermatogonia and spermatocytes | [145] |
| Mice | 7 days | 0–75 mg | Dose-dependent reduction of testicular and cauda epididymal sperm counts and serum testosterone | [129] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El. Khoury, D.; Fayjaloun, S.; Nassar, M.; Sahakian, J.; Aad, P.Y. Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals. Toxins 2019, 11, 515. https://doi.org/10.3390/toxins11090515
El. Khoury D, Fayjaloun S, Nassar M, Sahakian J, Aad PY. Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals. Toxins. 2019; 11(9):515. https://doi.org/10.3390/toxins11090515
Chicago/Turabian StyleEl. Khoury, Diala, Salma Fayjaloun, Marc Nassar, Joseph Sahakian, and Pauline Y. Aad. 2019. "Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals" Toxins 11, no. 9: 515. https://doi.org/10.3390/toxins11090515
APA StyleEl. Khoury, D., Fayjaloun, S., Nassar, M., Sahakian, J., & Aad, P. Y. (2019). Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals. Toxins, 11(9), 515. https://doi.org/10.3390/toxins11090515