Perennial Ryegrass Alkaloids Increase Respiration Rate and Decrease Plasma Prolactin in Merino Sheep under Both Thermoneutral and Mild Heat Conditions


 and
and 
Abstract
1. Introduction
2. Results
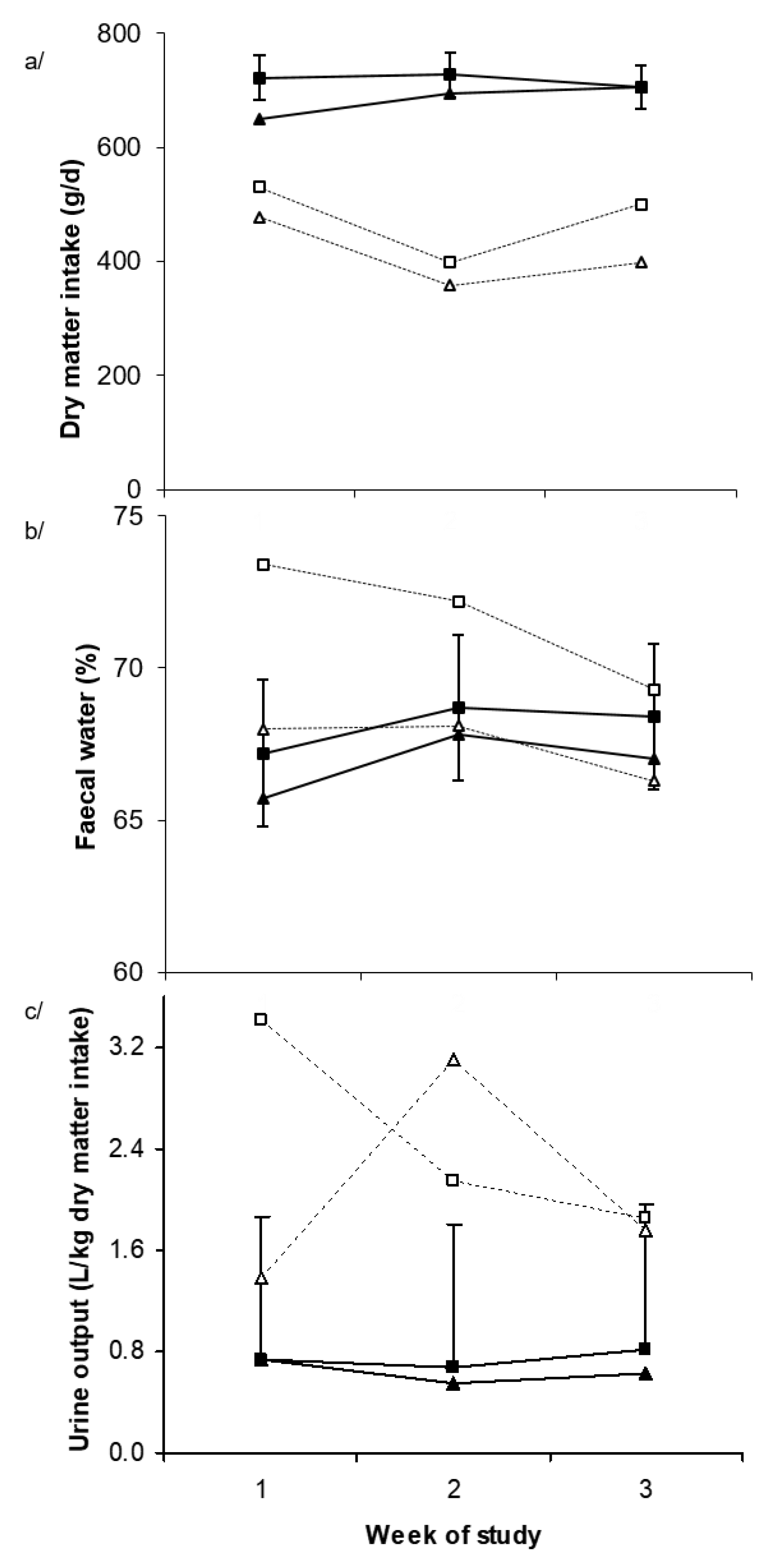
2.1. Environmental Conditions and Production Responses
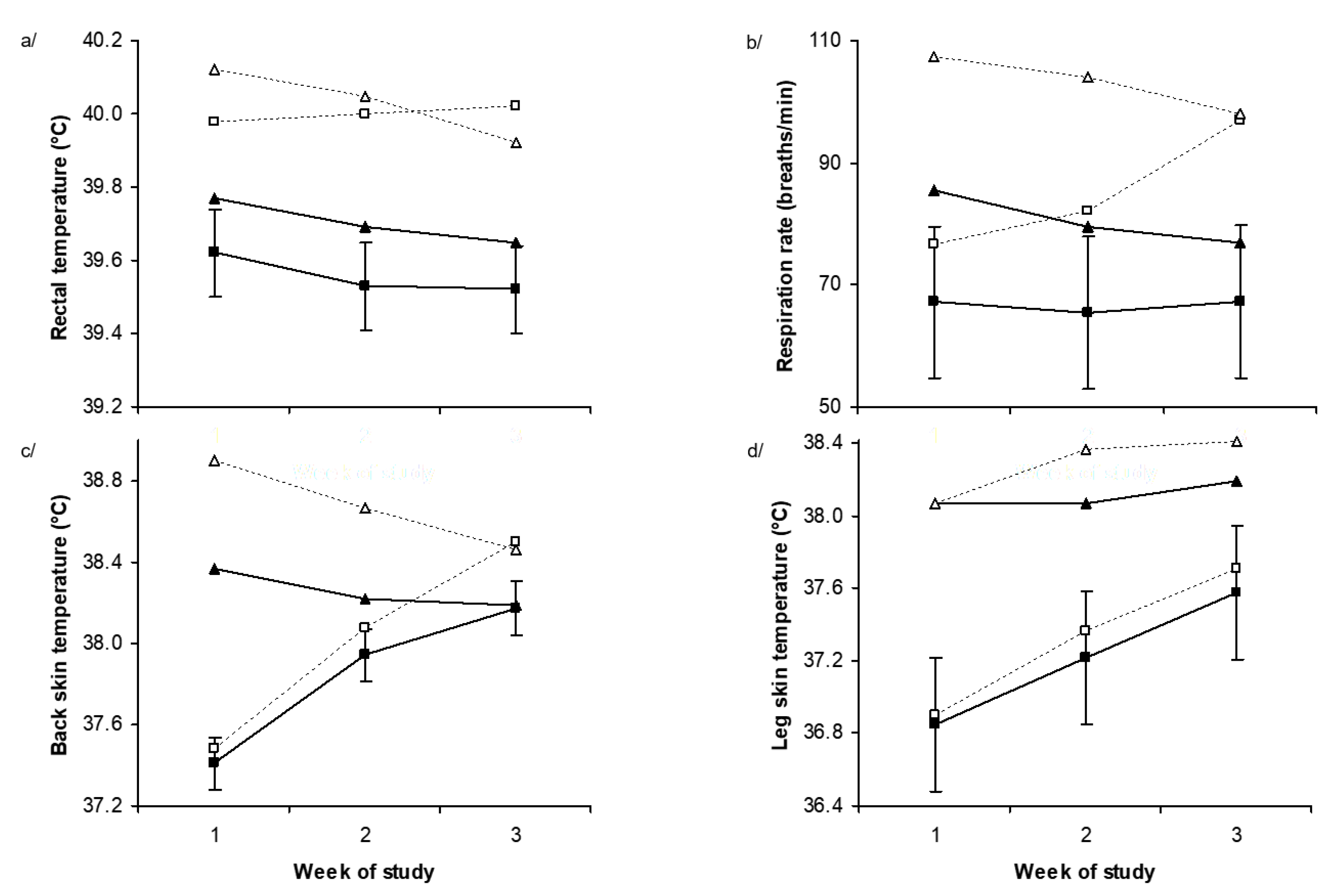
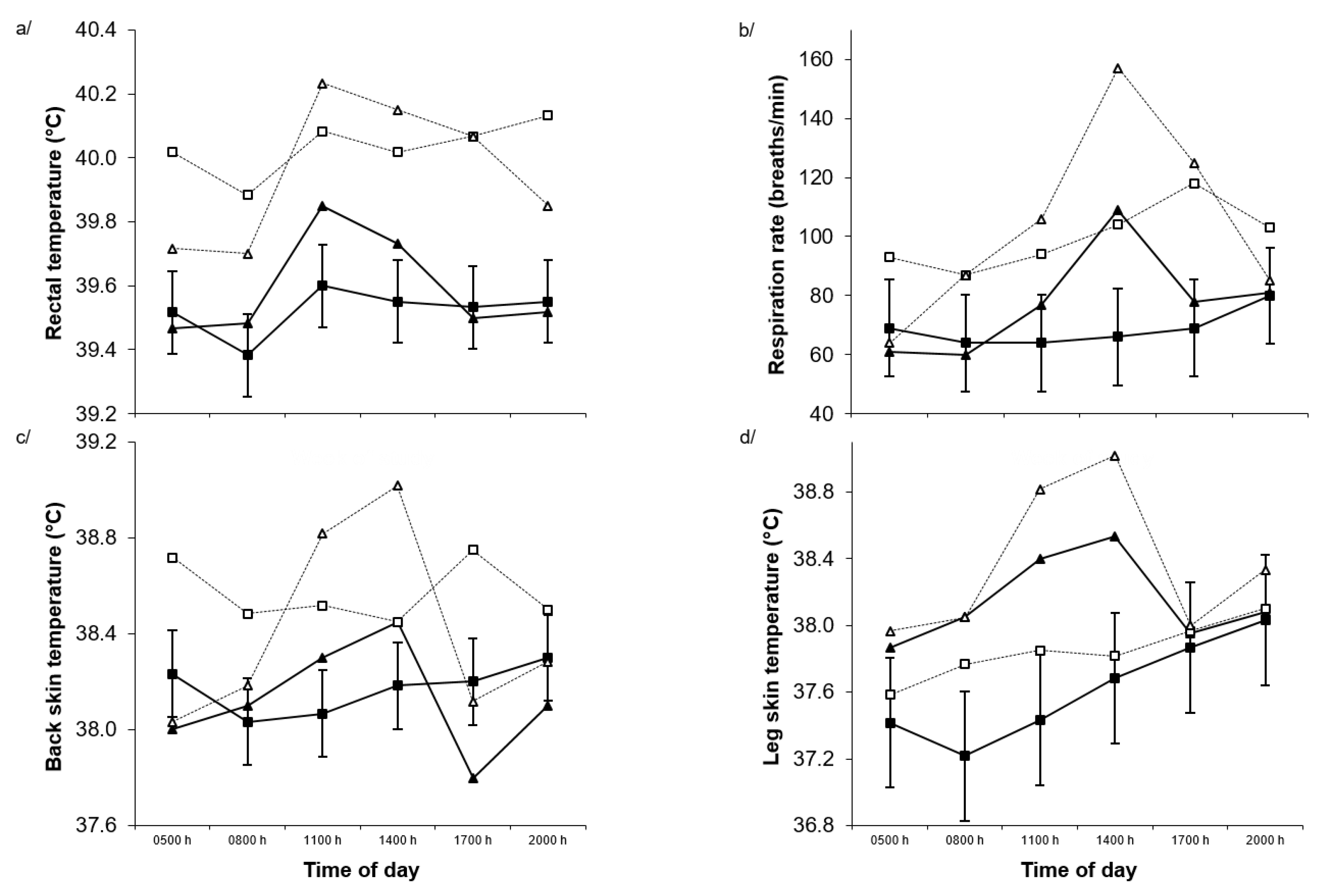
2.2. Physiological Responses
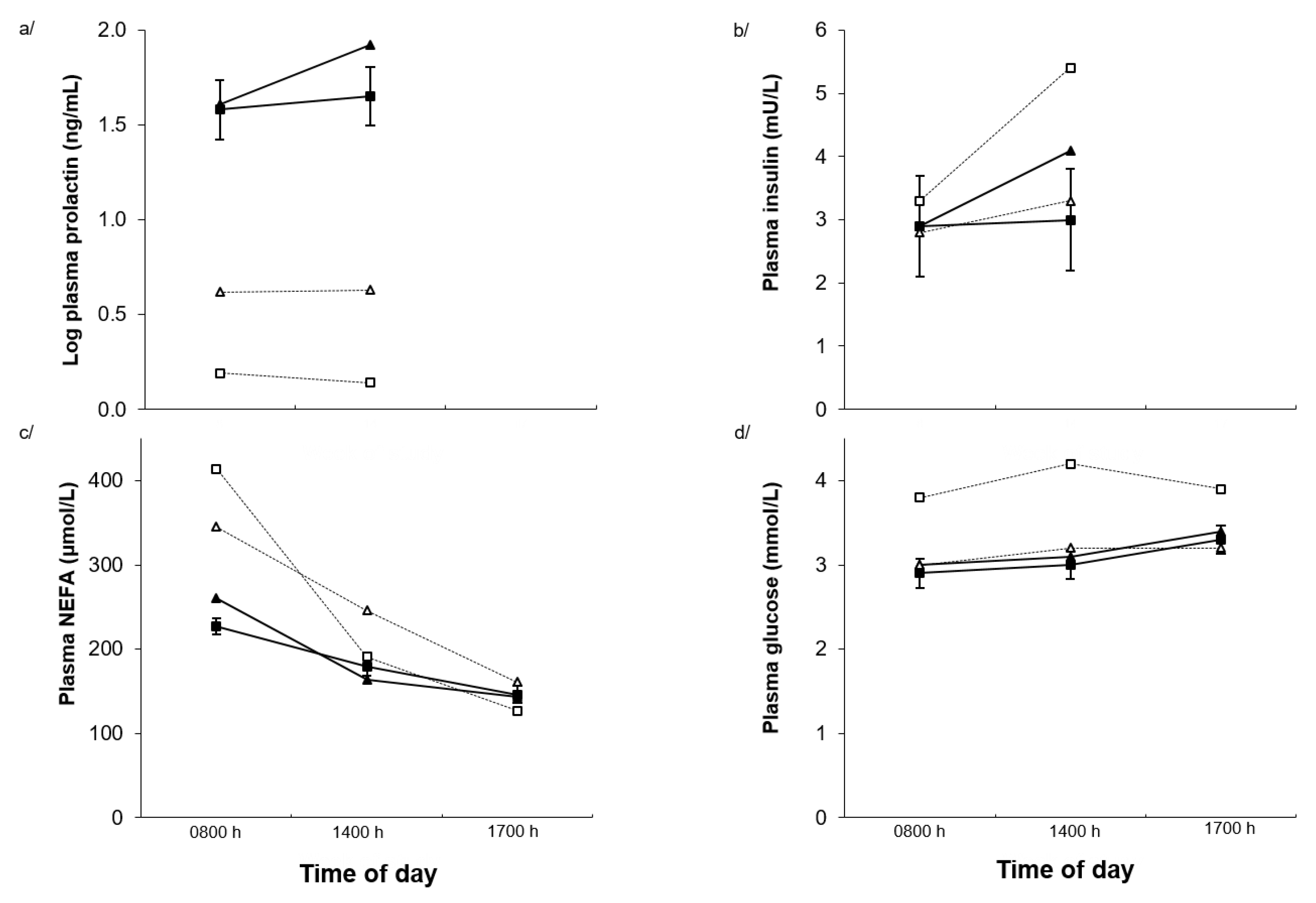
2.3. Plasma Hormones and Metabolites
3. Discussion
4. Conclusions
5. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reed, K.F.M. Perennial ryegrass toxins in Australian pasture. In Perennial Ryegrass Toxicosis in Australia; Reed, K.F.M., Page, S.W., Lean, I.J., Eds.; Meat and Livestock Australia: Attwood, Australia, 2005; pp. 11–17. [Google Scholar]
- McLeay, L.; Smith, B. Effects of the mycotoxins lolitrem B and paxilline on gastrointestinal smooth muscel, and cardiovascular and respiratory systems, and temperature in sheep. In Ryegrass Endophyte: An Essential New Zealand Symbiosis; Woodfield, D.R., Matthew, C., Eds.; New Zealand Grassland Association: Napier, New Zealand, 1999; pp. 69–76. [Google Scholar]
- Gallagher, R.T.; White, E.P.; Mortimer, P.H. Ryegrass staggers: Isolation of potent neurotoxins lolitrem A and lolitrem B from staggers-producing pastures. N. Z. Vet. J. 1981, 29, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.; Sutherland, B.; Fletcher, C. The impact of endophyte on the health and productivity of sheep grazing ryegrass-based pastures. In Ryegrass Endophyte: An Essential New Zealand Symbiosis; Woodfield, D.R., Matthew, C., Eds.; New Zealand Grassland Association: Napier, New Zealand, 1999; pp. 11–18. [Google Scholar]
- Henry, M.L.E.; Kemp, S.; Clarke, I.J.; Dunshea, F.R.; Leury, B.J. Production and physiological effects of perennial ryegrass alkaloids under thermoneutral conditions in merinos. Anim. Prod. Sci. 2016, 56, 1629–1636. [Google Scholar] [CrossRef]
- Henry, M.L.; Kemp, S.; Dunshea, F.R.; Leury, B.J. Physiological effects of ergot alkaloid and indole-diterpene consumption on sheep under hot and thermoneutral ambient temperature conditions. Animals 2016, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Combs, M.D.; Edwards, S.H.; Scherpenhuizen, J.M.; Narayan, E.J.; Kessell, A.E.; Piltz, J.; Raidal, S.R.; Ramsay, J.; Quinn, J.C. Development of a model for investigation of perennial ryegrass toxicosis in sheep. N. Z. Vet. J. 2018, 66, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Finch, S.; Imlach, W.; Dunlop, J.; Meredith, A.; Aldrich, R.; Dalziel, J. Mode of action of tremorgens. In Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses, Christchurch, New Zealand; 2006; pp. 363–364. [Google Scholar]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Osborn, T.G.; Schmidt, S.P.; Marple, D.N.; Rahe, C.H.; Steenstra, J.R. Effect of consuming fungus-infected and fungus-free tall fescue and ergotamine tartrate on selected physiological variables of cattle in environmentally controlled conditions. J. Anim. Sci. 1992, 70, 2501–2509. [Google Scholar] [CrossRef]
- Fletcher, L.; Barrell, G. Reduced liveweight gains and serum prolactin levels in hoggets grazing ryegrasses containing Lolium endophyte. N. Z. Vet. J. 1984, 32, 139–140. [Google Scholar] [CrossRef]
- Fletcher, L.; Sutherland, B.; Fletcher, C.; Easton, H. The impact of endophyte toxins on the health of grazing sheep—An evolving story. In Proceedings of the Second Pan Pacific Veterinary Conference, Massey University, Christchurch, New Zealand, 23–28 June 1996; pp. 31–42. [Google Scholar]
- Fletcher, L.; Easton, H. The evaluation and use of endophytes for pasture improvement. In Neotyphodium/Grass Interactions; Bacon, C.W., Hill, N.S., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 209–227. [Google Scholar]
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- Houpt, T. Water, electrolytes, and acid-base balance. In Dukes’ Physiology of Domestic Animals, 10th ed.; Swenson, M., Ed.; Cornell University Press: Ithaca, NY, USA, 1984; pp. 486–506. [Google Scholar]
- Fiorito, I.M.; Bunting, L.D.; Davenport, G.M.; Boling, J.A. Metabolic and endocrine responses of lambs fed Acremonium Coenophialum-infected or noninfected tall fescue hay at equivalent nutrient intake. J. Anim. Sci. 1991, 69, 2108–2114. [Google Scholar] [CrossRef]
- Keeler, R.; Wilson, N. Vasopressin contamination as a cause of some apparent renal actions of prolactin. Can. J. Physiol. Pharmacol. 1976, 54, 887–890. [Google Scholar] [CrossRef]
- Burstyn, P.G. Sodium and water metabolism under the influence of prolactin, aldosterone, and antidiuretic hormone. J. Physiol. 1978, 275, 39–50. [Google Scholar] [CrossRef]
- Hannah, S.M.; Paterson, J.A.; Williams, J.E.; Kerley, M.S.; Miner, J.L. Effects of increasing dietary levels of endophyte-infected tall fescue seed on diet digestibility and ruminal kinetics in sheep. J. Anim. Sci. 1990, 68, 1693–1701. [Google Scholar] [CrossRef]
- Aldrich, C.G.; Paterson, J.A.; Tate, J.; Kerley, M.S. The effects of endophyte-infected tall fescue consumption on diet utilization and thermal regulation in cattle. J. Anim. Sci. 1993, 71, 164–170. [Google Scholar] [CrossRef]
- Matthews, A.K.; Poore, M.H.; Huntington, G.B.; Green, J.T. Intake, digestion, and N metabolism in steers fed endophyte-free, ergot alkaloid-producing endophyte-infected, or nonergot alkaloid-producing endophyte-infected fescue hay. J. Anim. Sci. 2005, 83, 1179–1185. [Google Scholar] [CrossRef]
- Waghorn, G.C. Dags in Sheep Grazing Temperate Pastures: Causes and Consequences of Faecal Adhesion to Wool; Fundacao Estudos Agrarios Luiz Queiroz (FEALQ): Piracicaba, Brazil, 2001; pp. 440–441. [Google Scholar]
- Chauhan, S.S.; Celi, P.; Leury, B.J.; Clarke, I.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. J. Anim. Sci. 2014, 92, 3364–3374. [Google Scholar] [CrossRef]
- Britt, J.L.; Greene, M.A.; Bridges, W.C.; Klotz, J.L.; Aiken, G.E.; Andrae, J.G.; Pratt, S.L.; Long, N.M.; Schrick, F.N.; Strickland, J.R.; et al. Ergot alkaloid exposure during gestation alters. I. Maternal characteristics and placental development of pregnant ewes. J. Anim. Sci. 2019, 97, 1874–1890. [Google Scholar] [CrossRef]
- Beede, D.K.; Collier, R.J. Potential nutritional strategies for intensively managed cattle during thermal stress. J. Anim. Sci. 1986, 62, 543–554. [Google Scholar] [CrossRef]
- Gonzalez-Rivas, P.A.; DiGiacomo, K.; Giraldo, P.A.; Leury, B.J.; Cottrell, J.J.; Dunshea, F.R. Reducing rumen starch fermentation of wheat with three percent sodium hydroxide has the potential to ameliorate the effect of heat stress in grain-fed wethers. J. Anim. Sci. 2017, 95, 5547–5562. [Google Scholar] [CrossRef]
- Salah, M.S.; Alshaikh, M.A.; Alsaiadi, M.Y.; Mogawer, H.H. Effect of prolactin inhibition on thermoregulation, water and food intakes in heat-stressed fat-tailed male lambs. Anim. Sci. 1995, 60, 87–91. [Google Scholar] [CrossRef]
- Alamer, M. The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. As. J. Anim. Vet. Adv. 2011, 6, 1153–1169. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; Hugo, E.R.; Brandebourg, T.D.; LaPensee, C.R. Focus on prolactin as a metabolic hormone. Trends Endocrinol. Metab. 2006, 17, 110–116. [Google Scholar] [CrossRef]
- Oliver, J.W.; Schultze, A.E.; Rohrbach, B.W.; Fribourg, H.A.; Ingle, T.; Waller, J.C. Alterations in hemograms and serum biochemical analytes of steers after prolonged consumption of endophyte-infected tall fescue. J. Anim. Sci. 2000, 78, 1029–1035. [Google Scholar] [CrossRef]
- Browning, R., Jr.; Thompson, F.N. Endocrine and respiratory responses to ergotamine in Brahman and Hereford steers. Vet. Hum. Toxicol. 2002, 44, 149–154. [Google Scholar]
- McCracken, B.A.; Krysl, L.J.; Park, K.K.; Holcombe, D.W.; Judkins, M.B. Steers grazing endophyte-free tall fescue: Seasonal changes in nutrient quality, forage intake, digesta kinetics, ruminal fermentation, and serum hormones and metabolites. J. Anim. Sci. 1993, 71, 1588–1595. [Google Scholar] [CrossRef]
- Looper, M.L.; Edrington, T.S.; Flores, R.; Burke, J.M.; Callaway, T.R.; Aiken, G.E.; Schrick, F.N.; Rosenkrans, C.F., Jr. Influence of dietary endophyte (Neotyphodium coenophialum)-infected tall fescue (Festuca arundinacea) seed on fecal shedding of antibiotic resistance-selected Escherichia coli O157:H7 in ewes. J. Anim. Sci. 2007, 85, 1102–1108. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Bell, A.W.; Trigg, T.E. Relations between plasma non-esterified fatty acid metabolism and body tissue mobilization during chronic undernutrition in goats. Br. J. Nutr. 1988, 60, 633–644. [Google Scholar] [CrossRef]
- Johnson, M.M.; Peters, J.P. Technical note: An improved method to quantify nonesterified fatty acids in bovine plasma. J. Anim. Sci. 1993, 71, 753–756. [Google Scholar] [CrossRef]
- Harrington, K.C.; Beskow, W.B.; Hodgson, J. Recovery and viability of seeds ingested by goats. Pasture Pests Weeds 2011, 64, 75–80. [Google Scholar]
- Tindal, J.S.; Knaggs, G.S.; Hart, I.C.; Blake, L.A. Release of growth hormone in lactating and non-lactating goats in relation to behaviour, stages of sleep, electroencephalograms, environmental stimuli and levels of prolactin, insulin, glucose and free fatty acids in the circulation. J. Endocrinol. 1978, 76, 333–346. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Johnson-Barrett, J.J.; Butler, J.H.; Edgar, B.W.; Gunn, T.R. Studies of insulin-like growth factor -I and -II by specific radioligand assays in umbilical cord blood. Clin. Endocrinol. 1983, 19, 405–413. [Google Scholar] [CrossRef]
- Downing, J.A.; Joss, J.; Connell, P.; Scaramuzzi, R.J. Ovulation rate and the concentrations of gonadotrophic and metabolic hormones in ewes fed lupin grain. J. Reprod. Fertil. 1995, 103, 137–145. [Google Scholar] [CrossRef]
- Blache, D.; Tellam, R.L.; Chagas, L.M.; Blackberry, M.A.; Vercoe, P.E.; Martin, G.B. Level of nutrition affects leptin concentrations in plasma and cerebrospinal fluid in sheep. J. Endocrinol. 2000, 165, 625–637. [Google Scholar] [CrossRef]
- Clarke, I.J.; Mitchelhill, K.; Zachariah, E.; Findlay, J.K.; Funder, J.W. Analysis of androgen action on pituitary gonadotropin and prolactin secretion in ewes. Biol. Reprod. 1982, 26, 777–786. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkaloid (Alk) | NilAlk | Alk | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Temperature (T) | TN | Heat | TN | Heat | Sed | Alk | T | Alk × T |
| Average daily gain, g/d | −8.0 | −50.0 | −146 | −187 | 41.6 | <0.001 | 0.19 | 0.98 |
| Dry matter intake, g/d | 717 | 687 | 473 | 413 | 40.4 | <0.001 | 0.020 | 0.60 |
| Water intake, L/d | 1.97 | 2.22 | 2.34 | 2.74 | 0.432 | 0.15 | 0.44 | 0.81 |
| Water intake, L/kg DMI | 2.99 | 3.41 | 5.68 | 6.85 | 1.191 | 0.002 | 0.36 | 0.66 |
| Dry matter digestibility, % | 66.5 | 67.8 | 65.1 | 55.0 | 6.70 | 0.009 | 0.55 | 0.030 |
| Alkaloid (Alk) | NilAlk | Alk | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Temperature (T) | TN | Heat | TN | Heat | Sed | Alk | T | Alk × T |
| IGF-I, ng/mL | 25.0 1 | 25.8 | 36.2 | 19.3 | 10.61 | 0.76 | 0.29 | 0.25 |
| Somatotropin, ng/mL | 1.72 | 2.48 | 1.60 | 1.69 | 0.851 | 0.98 | 0.14 | 0.90 |
| Leptin, ng/mL | 1.20 | 1.30 | 1.21 | 1.09 | 0.092 | 0.28 | 0.89 | 0.23 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henry, M.L.E.; Kemp, S.; Clarke, I.J.; Dunshea, F.R.; Leury, B.J. Perennial Ryegrass Alkaloids Increase Respiration Rate and Decrease Plasma Prolactin in Merino Sheep under Both Thermoneutral and Mild Heat Conditions. Toxins 2019, 11, 479. https://doi.org/10.3390/toxins11080479
Henry MLE, Kemp S, Clarke IJ, Dunshea FR, Leury BJ. Perennial Ryegrass Alkaloids Increase Respiration Rate and Decrease Plasma Prolactin in Merino Sheep under Both Thermoneutral and Mild Heat Conditions. Toxins. 2019; 11(8):479. https://doi.org/10.3390/toxins11080479
Chicago/Turabian StyleHenry, Michelle L. E., Stuart Kemp, Iain J. Clarke, Frank R. Dunshea, and Brian J. Leury. 2019. "Perennial Ryegrass Alkaloids Increase Respiration Rate and Decrease Plasma Prolactin in Merino Sheep under Both Thermoneutral and Mild Heat Conditions" Toxins 11, no. 8: 479. https://doi.org/10.3390/toxins11080479
APA StyleHenry, M. L. E., Kemp, S., Clarke, I. J., Dunshea, F. R., & Leury, B. J. (2019). Perennial Ryegrass Alkaloids Increase Respiration Rate and Decrease Plasma Prolactin in Merino Sheep under Both Thermoneutral and Mild Heat Conditions. Toxins, 11(8), 479. https://doi.org/10.3390/toxins11080479