Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in a Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography


Abstract
1. Introduction
2. Results
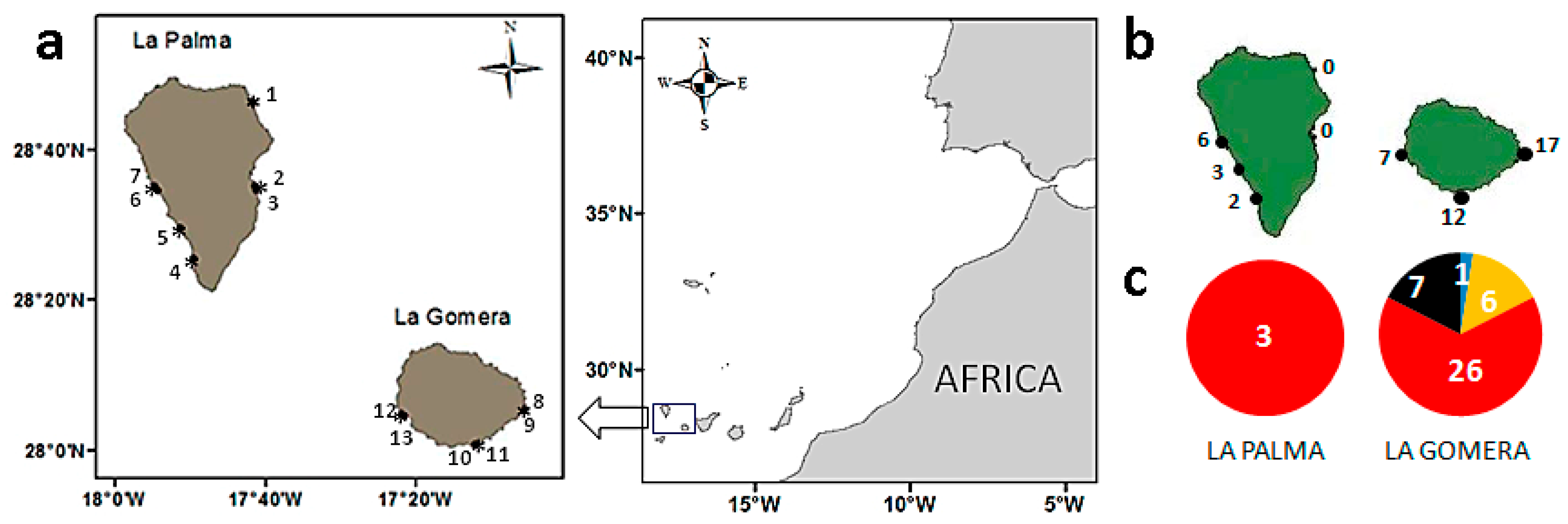
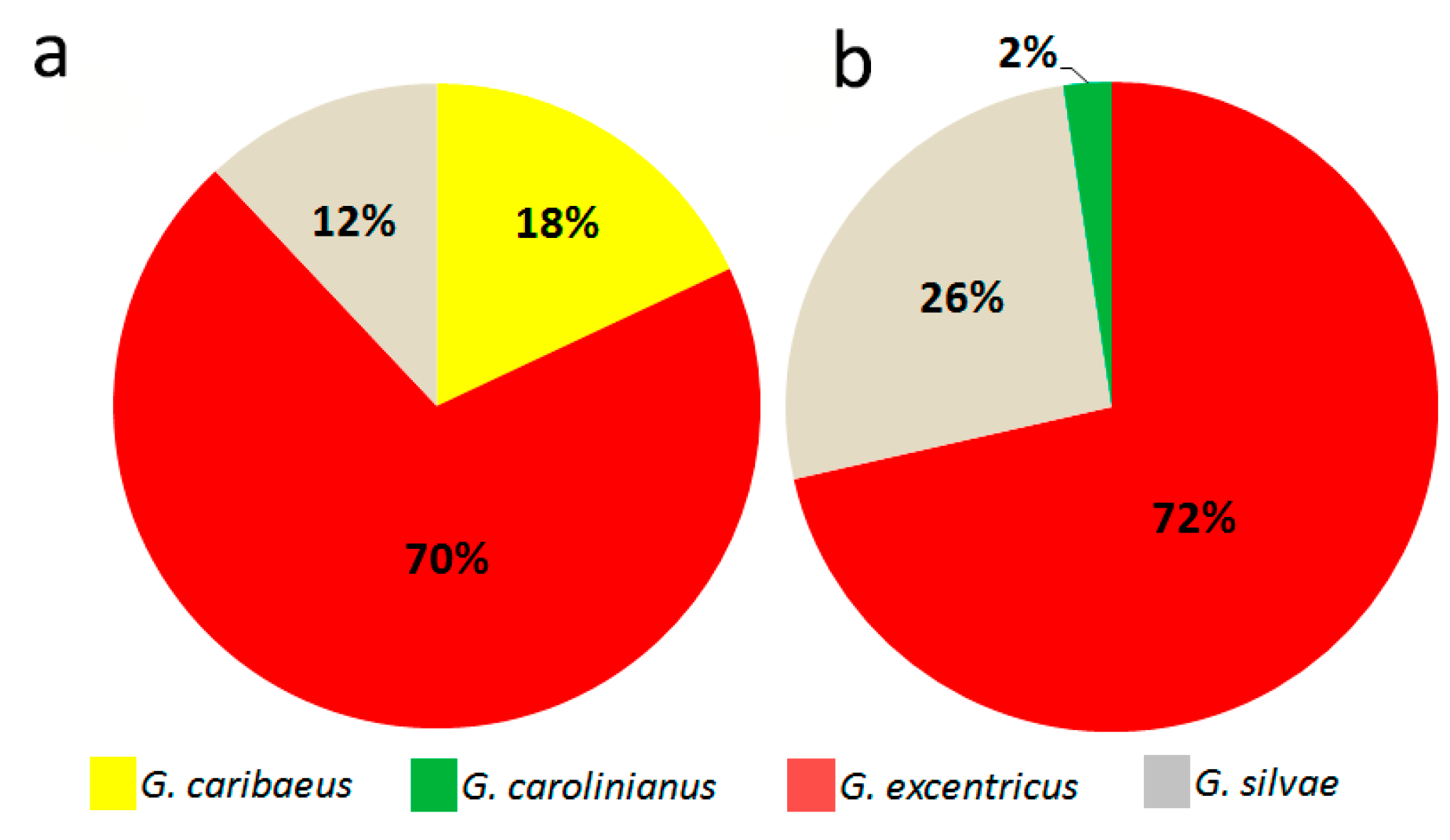
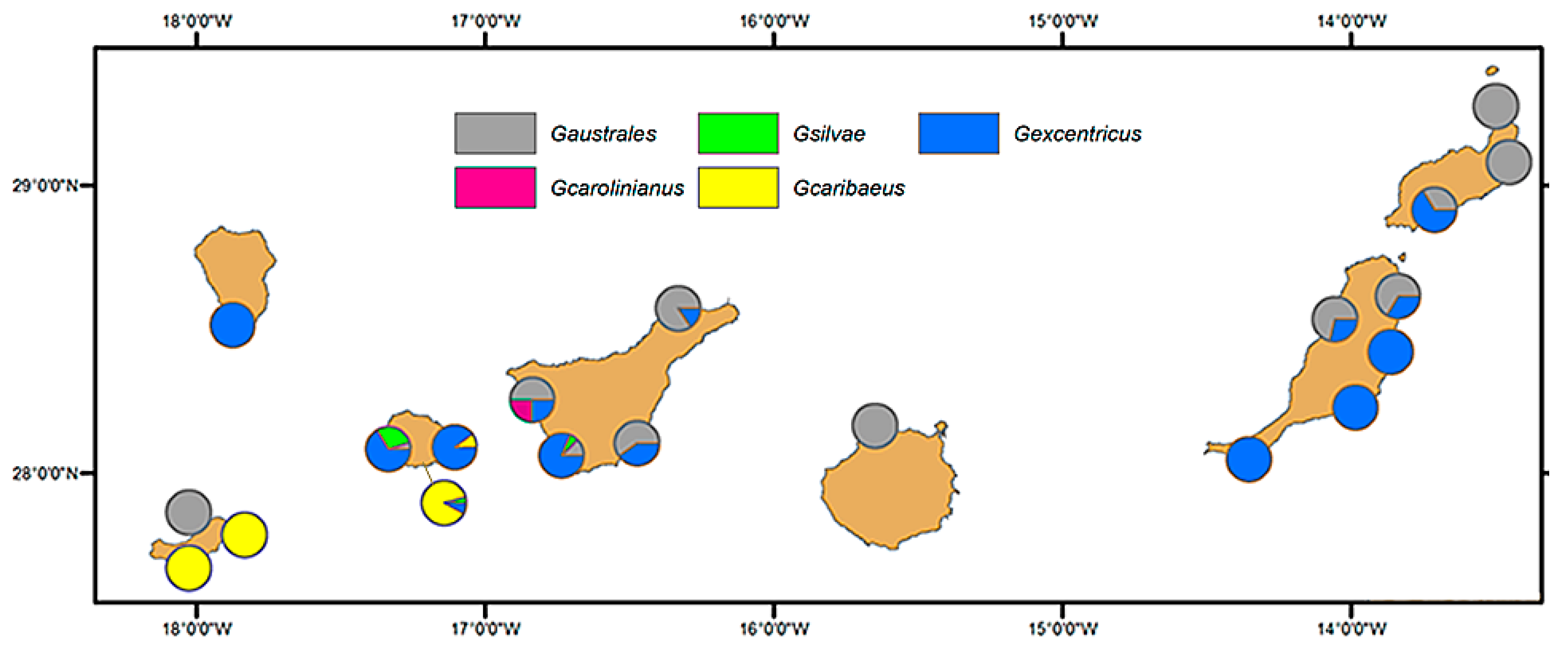
2.1. Diversity and Abundance of Gambierdiscus from La Palma and La Gomera
2.2. Epiphytic Dinoflagellate Composition
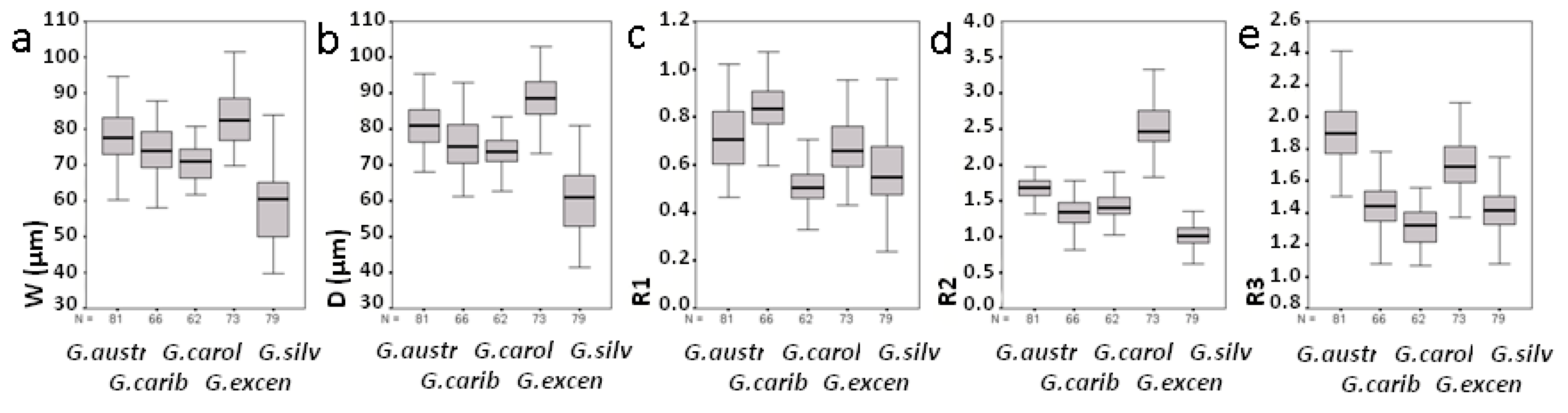
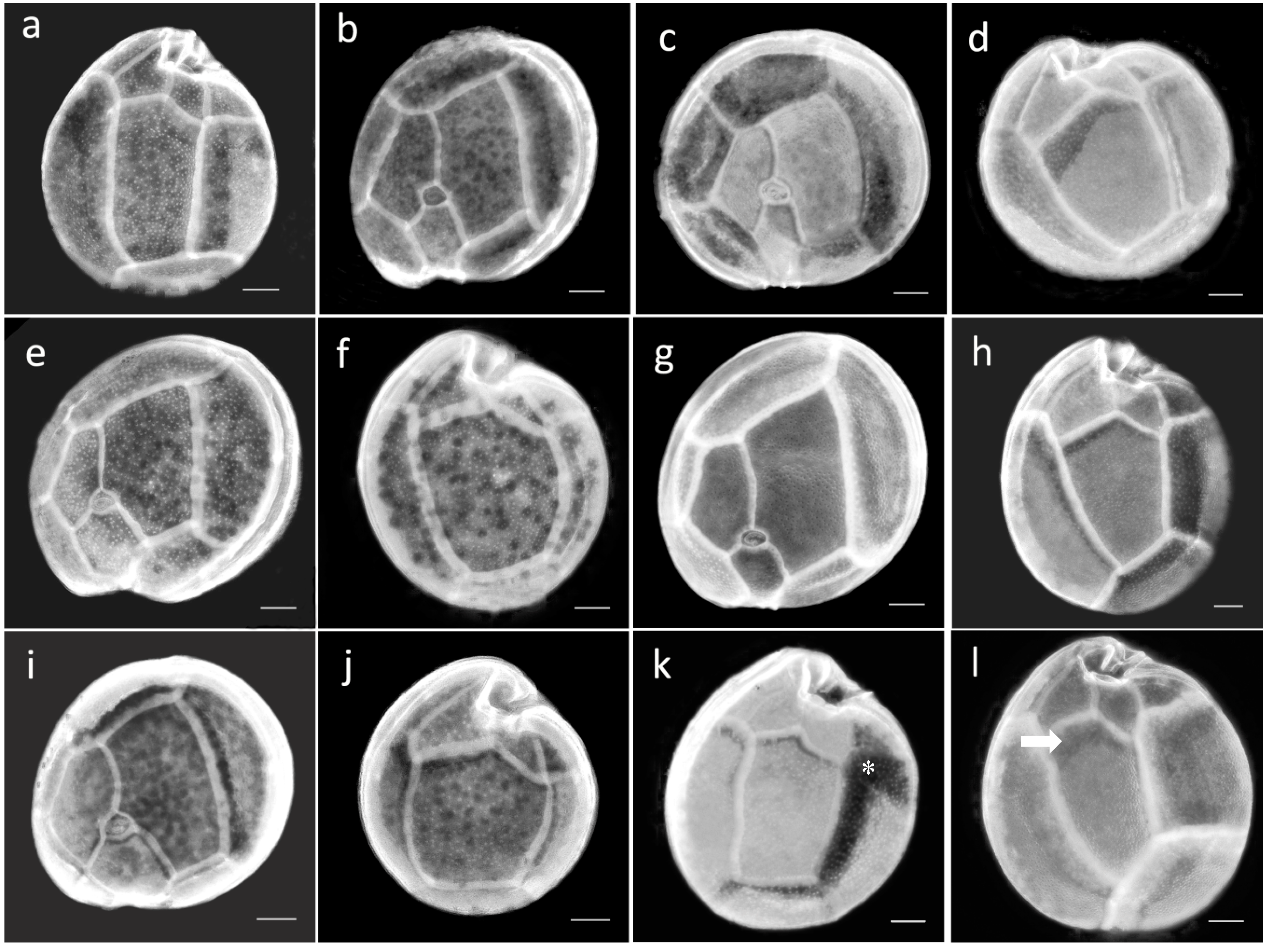
2.3. Morphological Study of Cultured Gambierdiscus Cells
2.4. Morphological Study of Field Cells of Gambierdiscus
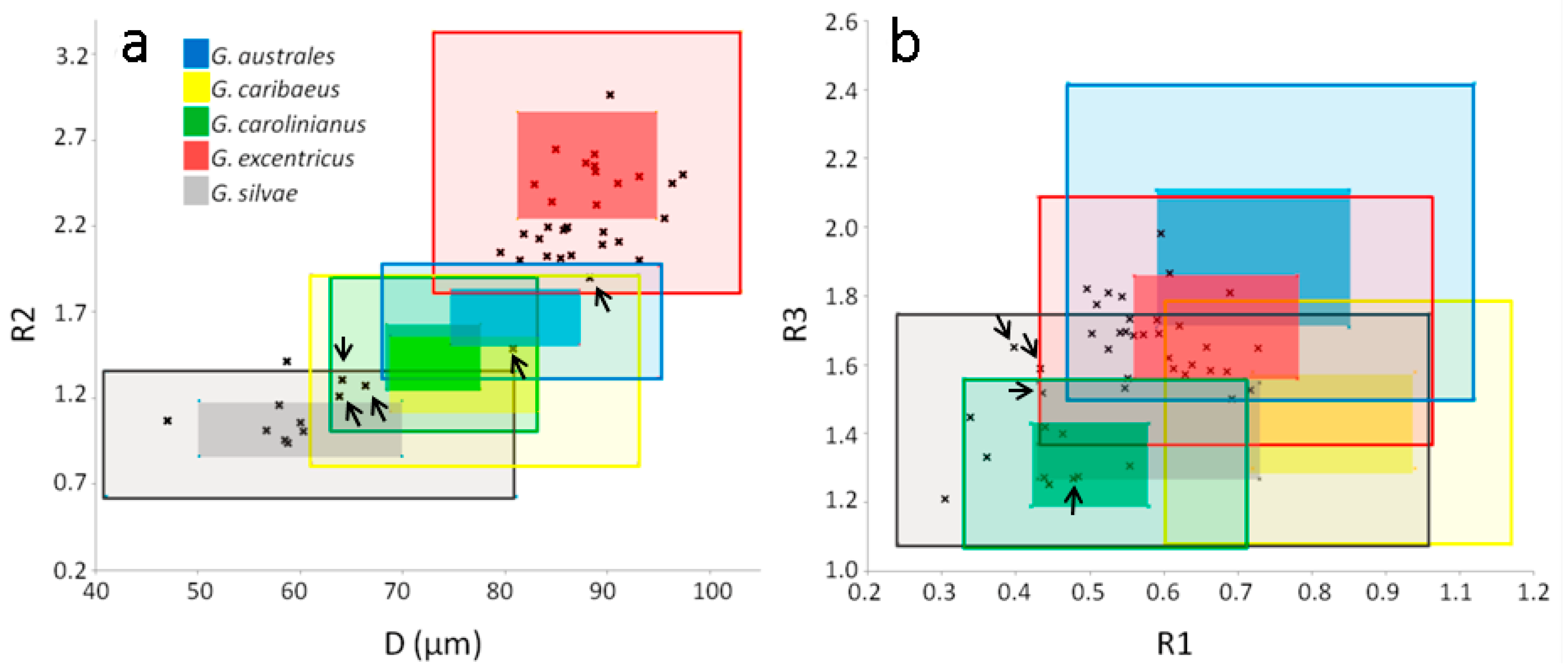
2.5. Gambierdiscus Diversity: Genetics vs. Morphology
3. Discussion
3.1. Geographical Distribution of the Species of Gambierdiscus Detected in the Archipelago of Canary Islands
3.2. Morphological Study
3.3. Comparisons of the Results: Genetics vs. Morphology
4. Materials and Methods
4.1. Field Sampling and Processing
4.2. Cell Enumeration and Light Microscopy
4.3. DNA Extraction, PCR Amplification, and DNA Sequencing
4.4. Morphological Study of Gambierdiscus
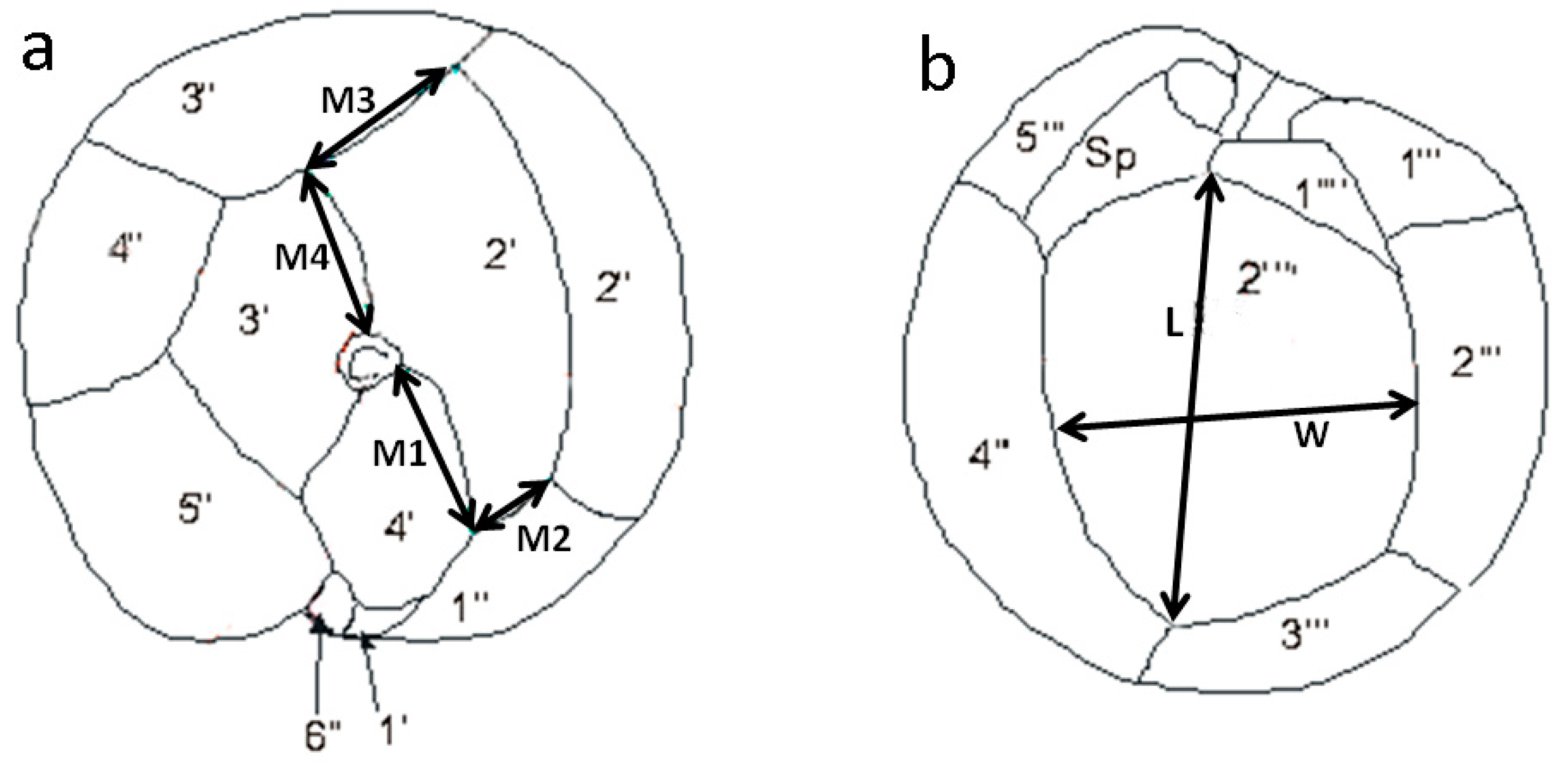
4.4.1. Morphometric Analysis
4.4.2. Cultured Cells
4.4.3. Field Samples
4.5. Comparison of the Morphological versus Genetic Data
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T. Chemistry, etiology, and food chain dynamics of marine toxins. Proc. Jpn. Acad. Ser. B 2005, 81, 43–51. [Google Scholar] [CrossRef]
- Ledreux, A.; Brand, H.; Chinain, M.; Bottein, M.-Y.D.; Ramsdell, J.S. Dynamics of ciguatoxins from Gambierdiscus polynesiensis in the benthic herbivore Mugil cephalus: Trophic transfer implications. Harmful Algae 2014, 39, 165–174. [Google Scholar] [CrossRef]
- Stewart, I.; Eaglesham, G.K.; Poole, S.; Graham, G.; Paulo, C.; Wickramasinghe, W.; Sadler, R.; Shaw, G.R. Establishing a public health analytical service based on chemical methods for detecting and quantifying Pacific ciguatoxin in fish samples. Toxicon 2010, 56, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Tester, P.A.; Vandersea, M.W.; Buckel, C.A.; Kibler, S.R.; Holland, W.C.; Davenport, E.D.; Clark, R.D.; Edwards, K.F.; Taylor, J.C.; Vander Pluym, J.L.; et al. Gambierdiscus (Dinophyceae) species diversity in the Flower Garden Banks National Marine Sanctuary, Northern Gulf of Mexico, USA. Harmful Algae 2013, 29, 1–9. [Google Scholar] [CrossRef]
- Lewis, R.J. The changing face of ciguatera. Toxicon 2001, 39, 97–106. [Google Scholar] [CrossRef]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera Fish Poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef] [PubMed]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-González, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.; Dickey, R.W. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.; Suárez, F.; Ramírez, A.; Acosta, F. Ciguatera, an Emerging Human Poisoning in Europe. J. Aquac. Mar. Boil. 2015, 3, 00053. [Google Scholar]
- Aligizaki, K.; Nikolaidis, G. Morphological identification of two tropical dinoflagellates of the genera Gambierdiscus and Sinophysis in the Mediterranean Sea. J. Biol. Res. Thessalon. 2008, 9, 75–82. [Google Scholar]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Yamaguchi, H.; Adachi, M. Morphology of G ambierdiscus scabrosus sp. nov. (Gonyaulacales): A new epiphytic toxic dinoflagellate from coastal areas of Japan. J. Phycol. 2014, 50, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.I.; Riobó, P.; Bravo, I. Canary Islands (NE Atlantic) as a biodiversity ’hotspot’ of Gambierdiscus: Implications for future trends of ciguatera in the area. Harmful Algae 2017, 67, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Villareal, T.; Hanson, S.; Qualia, S.; Jester, E.; Granade, H.; Dickey, R.; Villareal, T. Petroleum production platforms as sites for the expansion of ciguatera in the northwestern Gulf of Mexico. Harmful Algae 2007, 6, 253–259. [Google Scholar] [CrossRef]
- Tester, P.A.; Feldman, R.L.; Nau, A.W.; Kibler, S.R.; Litaker, R.W. Ciguatera fish poisoning and sea surface temperatures in the Caribbean Sea and the West Indies. Toxicon 2010, 56, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Model. 2015, 316, 194–210. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with Description of Gambierdiscus silvae sp. nov., a New Potentially Toxic Epiphytic Benthic Dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef]
- Pérez-Arellano, J.-L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera Fish Poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef]
- Costa, P.; Estevez, P.; Castro, D.; Solino, L.; Gouveia, N.; Santos, C.; Rodrigues, S.; Leao, J.; Gago-Martinez, A. New Insights into the Occurrence and Toxin Profile of Ciguatoxins in Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Leao, J.M.; Vilariño, O.; Diogène, J.; Gago-Martínez, A. An Attempt to Characterize the Ciguatoxin Profile in Seriola fasciata Causing Ciguatera Fish Poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Leao, J.M.; Yasumoto, T.; Dickey, R.; Gago-Martinez, A. Implementation of liquid chromatography tandem mass spectrometry for the analysis of ciguatera fish poisoning in contaminated fish samples from Atlantic coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus, sp. nov., G. australes, sp. nov., and G. polynesiensis, sp. nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Chinain, M.; Holmes, M.J.; Holland, W.C.; Tester, P.A. Taxonomy of Gambierdiscus including four new species, Gambierdiscus caribaeus, Gambierdiscus carolinianus, Gambierdiscus carpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dinophyceae). Phycologia 2009, 48, 344–390. [Google Scholar] [CrossRef]
- Vandersea, M.W.; Kibler, S.R.; Holland, W.C.; Tester, P.A.; Schultz, T.F.; Faust, M.A.; Holmes, M.J.; Chinain, M.; Litaker, R.W. Development Of Semi-Quantitative Pcr Assays For The Detection And Enumeration Of Gambierdiscus Species (Gonyaulacales, Dinophyceae)(1). J. Phycol. 2012, 48, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Tester, P.A.; Faust, M.A.; Vandersea, M.W.; Kibler, S.R.; Chinain, M.; Holmes, M.; Holland, C.; Litaker, R.W. Gambierdiscus toxicus: Taxonomic uncertainties concerning Gambierdiscus toxicus: Proposed epitype. In 12th International Conference on Harmful Algae; International Society for the Study of Harmful Algae and the Intergovernmental Oceanographic Commission of UNESCO: Copenhagen, Denmark, 2008; pp. 269–271. [Google Scholar]
- Munir, S.; Siddiqui, P.; Morton, S.L.; Munir, S. The occurrence of the ciguatera fish poisoning producing dinoflagellate genus Gambierdiscus in Pakistan waters. ALGAE 2011, 26, 317–325. [Google Scholar] [CrossRef]
- Tudó, A.; Toldrá, A.; Andree, K.; Rey, M.; Fernández-Tejedor, M.; Campás, M.; Diogène, J. First report of Gambierdiscus in the Western Mediterranean Sea (Balearic Islands). Harmful Algae News 2018, 59, 22–23. [Google Scholar]
- Kaufmann, M.; Böhm-Beck, M. Gambierdiscus and related benthic dinoflagellates from Madeira archipelago (NE Atlantic). Harmful Algae News 2013, 47, 18–19. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-Wide Electronic Publication, National University of Ireland: Galway, Ireland, 2010; Available online: http://www.algaebase.org (accessed on 2 December 2010).
- Ennaffah, B.; Chaira, K. First report of Gambierdiscus in Moroccan Atlantic waters. Harmful Algae News 2015, 50, 20. [Google Scholar]
- Nascimento, S.M.; Melo, G.; Salgueiro, F.; Diniz, B.D.S.; Fraga, S. Morphology of Gambierdiscus excentricus (Dinophyceae) with emphasis on sulcal plates. Phycologia 2015, 54, 628–639. [Google Scholar] [CrossRef]
- Soler-Onís, E.; Fernandez-Zabala, J.; Ojeda-Rodriguez, A.; Amorin, A. Bloom of Gambierdicus caribaeus in the temperate-subtropical waters of El Hierro, Canary Islands (North East Atlantic). Harmful Algae News 2016, 55, 14–17. [Google Scholar]
- Silva, E.S. Contribution a L’étude du microplancton de Dakar et des régions maritimes voisines. Bull. Int. Law Soc. 1956, 18, 335–371. [Google Scholar]
- Rhodes, L.; Smith, K.F.; Verma, A.; Curley, B.G.; Harwood, D.T.; Murray, S.; Kohli, G.S.; Solomona, D.; Rongo, T.; Munday, R.; et al. A new species of Gambierdiscus (Dinophyceae) from the south-west Pacific: Gambierdiscus honu sp. nov. Harmful Algae 2017, 65, 61–70. [Google Scholar] [CrossRef]
- Adachi, R.; Fukuyo, Y. The thecal structure of the marine toxic dinoflagellate Gambierdiscus toxicus gen. et. sp. nov. collected in a ciguatera endemic area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef]
- Nishimura, T.; Hariganeya, N.; Tawong, W.; Sakanari, H.; Yamaguchi, H.; Adachi, M. Quantitative PCR assay for detection and enumeration of ciguatera-causing dinoflagellate Gambierdiscus spp. (Gonyaulacales) in coastal areas of Japan. Harmful Algae 2016, 52, 11–22. [Google Scholar] [CrossRef]
- Smith, K.F.; Biessy, L.; Argyle, P.A.; Trnski, T.; Halafihi, T.; Rhodes, L.L. Molecular Identification of Gambierdiscus and Fukuyoa (Dinophyceae) from Environmental Samples. Mar. Drugs 2017, 15, 243. [Google Scholar] [CrossRef]
- Fritz, L.; Triemer, R.E. A rapid simple technique utilizing calcofluor white M2R for the visualization of dinoflagellate thecal plates. J. Phycol. 1985, 21, 662–664. [Google Scholar] [CrossRef]
- Lenaers, G.; Maroteaux, L.; Michot, B.; Herzog, M. Dinoflagellates in evolution. A molecular phylogenetic analysis of large subunit ribosomal RNA. J. Mol. Evol. 1989, 29, 40–51. [Google Scholar] [CrossRef]
- Kofoid, C.A. On the Peridinium steini Jörgensen, with a note on the nomenclature of the skeleton of the Peridinidae. Arch. Protistenkd. 1909, 16, 25–47. [Google Scholar]
- Besada, E.G.; Loeblich, L.A.; Loeblich, A.R., III. Observations on tropical, benthic dinoflagellates form ciguatera-endemic areas: Coolia, Gambierdiscus, and Ostreopsis. Bull. Mar. Sci. 1982, 32, 723–735. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Island | Locality | Coordinates | Type of Sample |
|---|---|---|---|---|
| 1 | La Palma | Charco azul | 28.81089, −17.7642 | macrophyte |
| 2 | La Palma | Cancajos beach | 28.65232, −17.75951 | macrophyte |
| 3 | La Palma | Caleta Ballena | 28.64465, −17.7573 | macrophyte |
| 4 | La Palma | La Zamora beach | 28.51485, −17.87486 | macrophyte |
| 5 | La Palma | Charco verde | 28.57261, −17.90008 | macrophyte |
| 6 | La Palma | Tazacorte harbor | 28.6437, −17.94325 | net |
| 7 | La Palma | Tazacorte beach | 28.65157, −17.94887 | macrophyte |
| 8 | La Gomera | La Cueva beach | 28.09081, −17.10565 | macrophyte |
| 9 | La Gomera | San Sebastián harbor | 28.089, −17.10767 | net |
| 10 | La Gomera | Santiago beach | 28.02732, −17.1979 | macrophyte |
| 11 | La Gomera | Santiago harbor | 28.02629, −17.19755 | net |
| 12 | La Gomera | Valle Rey harbor | 28.08069, −17.33259 | net |
| 13 | La Gomera | Charco Condesa | 28.08397, −17.33653 | macrophyte |
| Component | ||
|---|---|---|
| 1 | 2 | |
| Prorocentrum | 0.568 | - |
| Coolia | 0.583 | - |
| Sinophysis | 0.709 | - |
| Ostreopsis | - | 0.685 |
| Vulcanodinium | 0.524 | - |
| Heterocapsa | - | 0.792 |
| Scripsiella | - | 0.625 |
| Gambierdiscus | 0.706 | - |
| Parameter | Species | Mean | Std | Max | Min |
|---|---|---|---|---|---|
| D (cell depth) | G. australes | 81 | 6.3 | 95 | 68 |
| G. caribaeus | 76 | 7.2 | 93 | 61 | |
| G. carolinianus | 73 | 4.6 | 83 | 63 | |
| G. excentricus | 88 | 6.8 | 103 | 73 | |
| G. silvae | 60 | 9.9 | 81 | 41 | |
| W (cell width) | G. australes | 78 | 7.5 | 95 | 60 |
| G. caribaeus | 74 | 6.7 | 88 | 58 | |
| G. carolinianus | 71 | 4.9 | 81 | 62 | |
| G. excentricus | 83 | 7.9 | 101 | 70 | |
| G. silvae | 59 | 9.7 | 84 | 40 | |
| R1 | G. australes | 0.72 | 0.13 | 1.02 | 0.47 |
| G. caribaeus | 0.83 | 0.11 | 1.07 | 0.60 | |
| G. carolinianus | 0.50 | 0.08 | 0.71 | 0.33 | |
| G. excentricus | 0.67 | 0.11 | 0.96 | 0.43 | |
| G. silvae | 0.58 | 0.15 | 0.96 | 0.24 | |
| R2 | G. australes | 1.67 | 0.16 | 1.97 | 1.32 |
| G. caribaeus | 1.34 | 0.22 | 1.92 | 0.82 | |
| G. carolinianus | 1.44 | 0.19 | 1.90 | 1.02 | |
| G. excentricus | 2.55 | 0.31 | 3.33 | 1.82 | |
| G. silvae | 1.02 | 0.16 | 1.36 | 0.63 | |
| R3 | G. australes | 1.91 | 0.20 | 2.42 | 1.50 |
| G. caribaeus | 1.44 | 0.14 | 1.78 | 1.08 | |
| G. carolinianus | 1.31 | 0.12 | 1.56 | 1.07 | |
| G. excentricus | 1.71 | 0.15 | 2.09 | 1.37 | |
| G. silvae | 1.41 | 0.14 | 1.75 | 1.08 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo, I.; Rodriguez, F.; Ramilo, I.; Rial, P.; Fraga, S. Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in a Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography. Toxins 2019, 11, 423. https://doi.org/10.3390/toxins11070423
Bravo I, Rodriguez F, Ramilo I, Rial P, Fraga S. Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in a Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography. Toxins. 2019; 11(7):423. https://doi.org/10.3390/toxins11070423
Chicago/Turabian StyleBravo, Isabel, Francisco Rodriguez, Isabel Ramilo, Pilar Rial, and Santiago Fraga. 2019. "Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in a Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography" Toxins 11, no. 7: 423. https://doi.org/10.3390/toxins11070423
APA StyleBravo, I., Rodriguez, F., Ramilo, I., Rial, P., & Fraga, S. (2019). Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) in a Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography. Toxins, 11(7), 423. https://doi.org/10.3390/toxins11070423