Isolation and Characterization of Insecticidal Toxins from the Venom of the North African Scorpion, Buthacus leptochelys

 ,
,
Abstract
1. Introduction
2. Results and Discussion
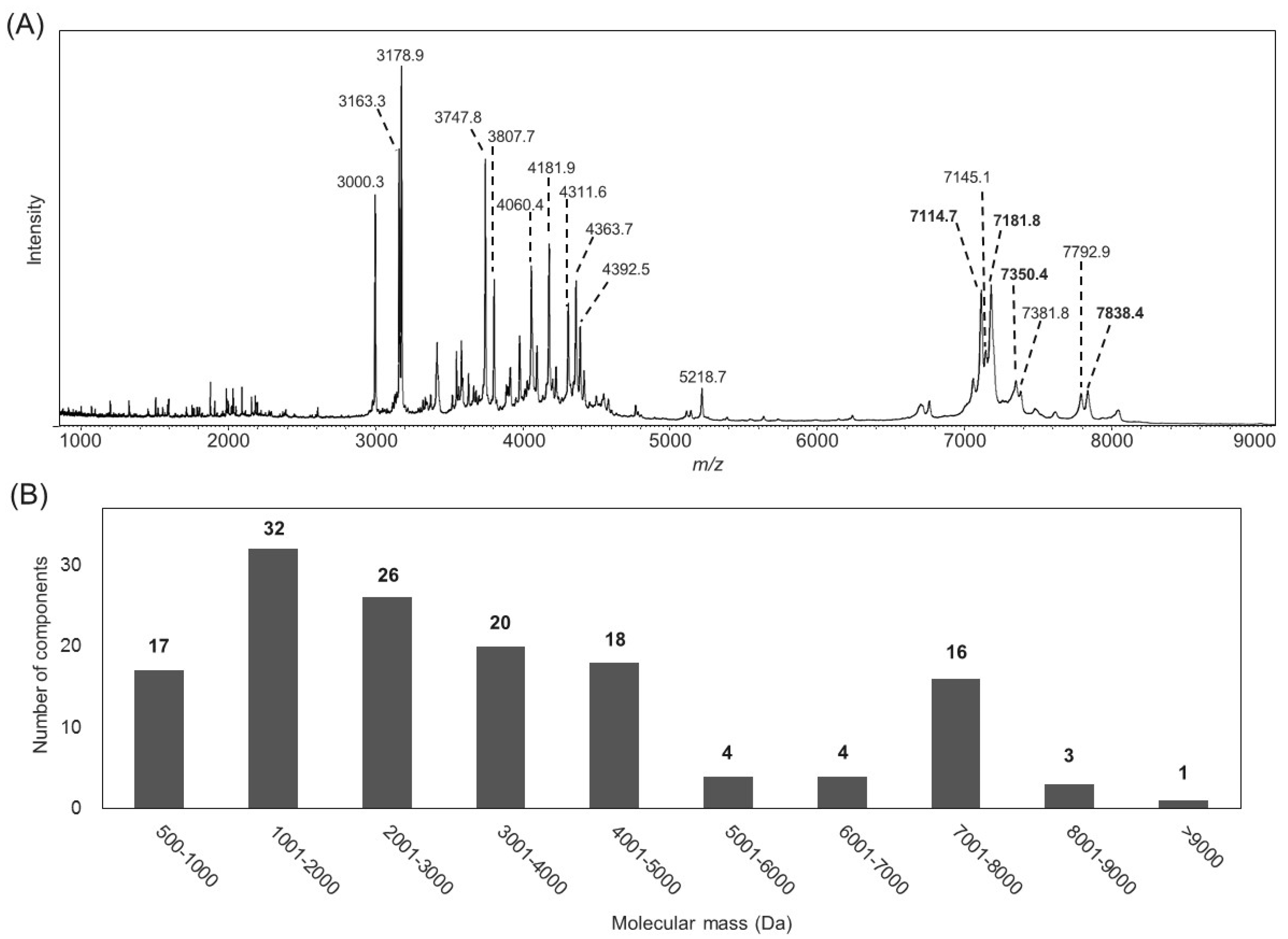
2.1. Characterization of the B. leptochelys Venom
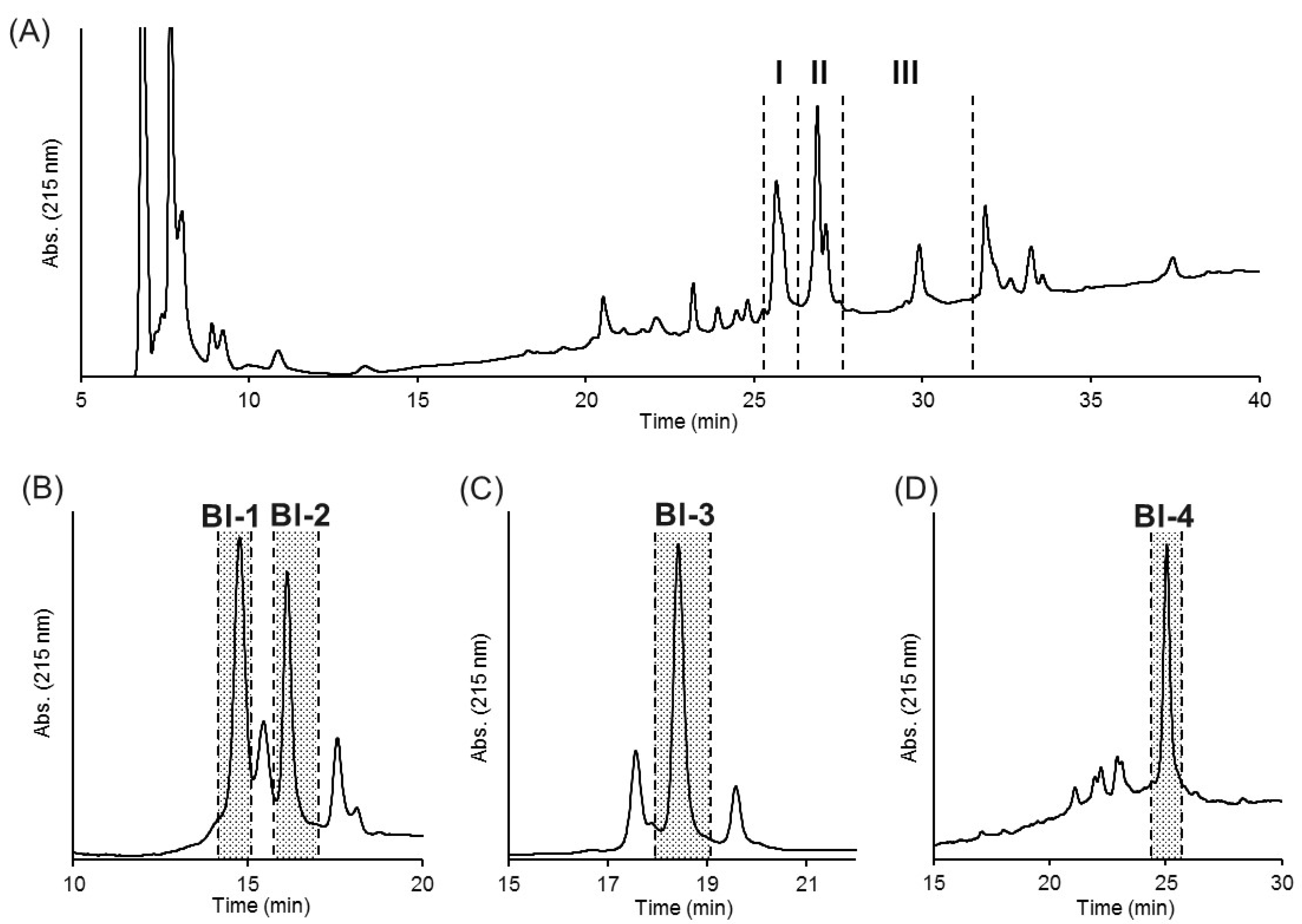
2.2. Purification of Insecticidal Peptides
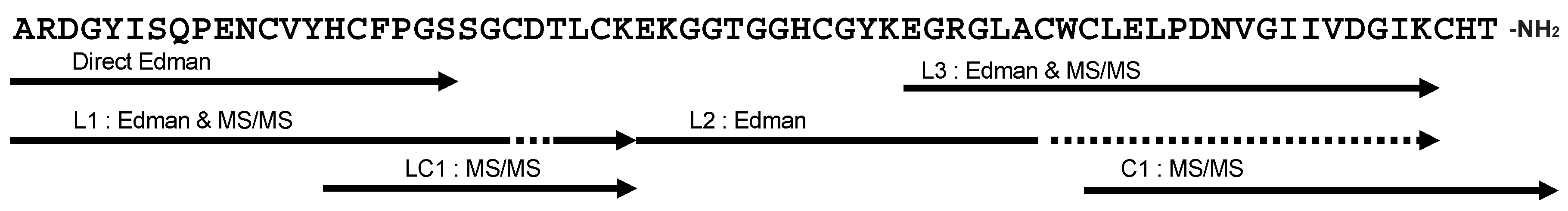
2.3. Primary Structure of Bl-1
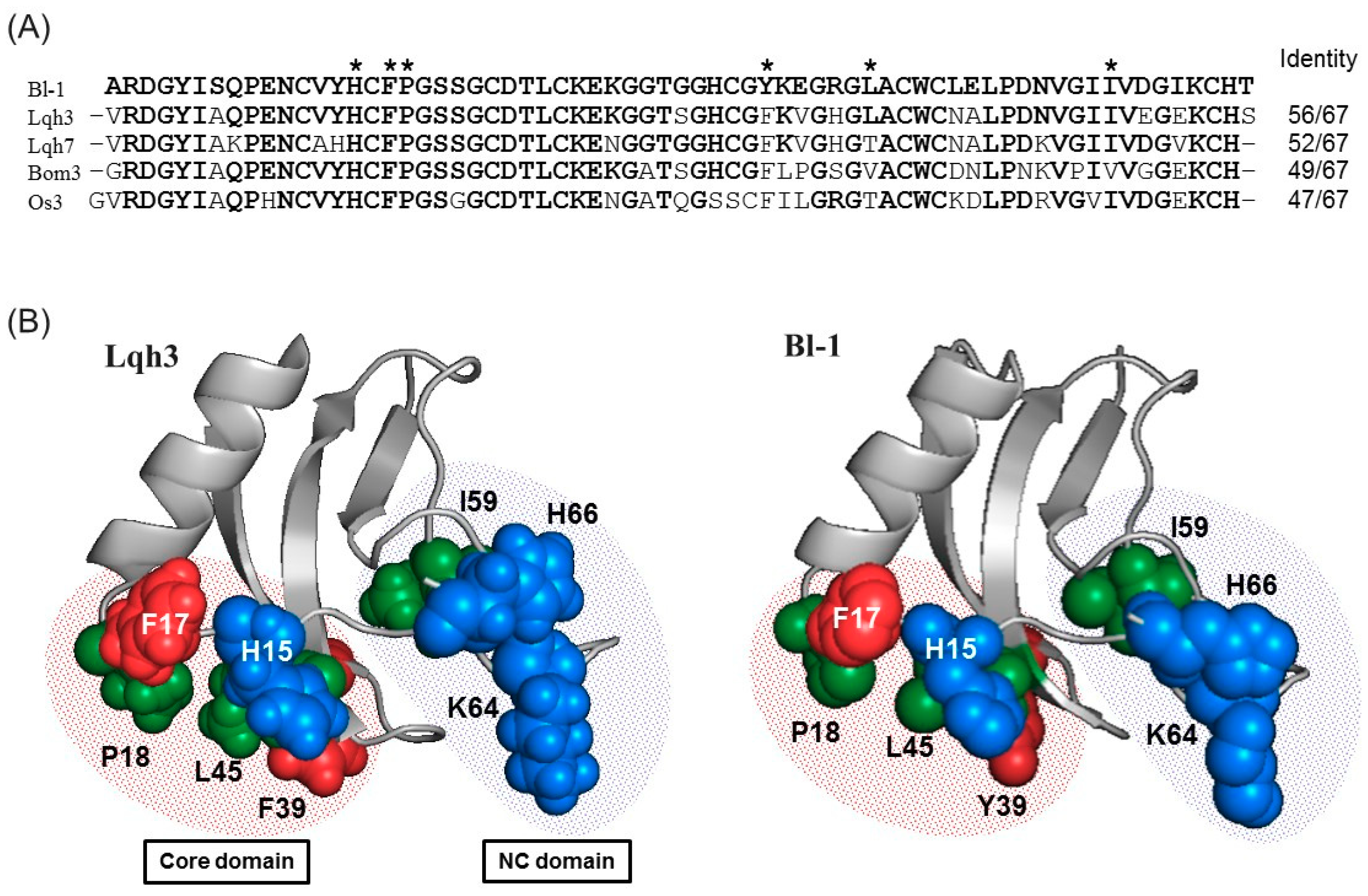
2.4. Sequence Comparison
3. Conclusions
4. Materials and Methods
4.1. Collection of Venom
4.2. Bioassay
4.3. Mass Spectrometric Analysis
4.4. HPLC Purification
4.5. Determination of N-terminal Sequence
4.6. Enzymatic Digestion
4.7. Homology Modeling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dunlop, J.A. Geological history and phylogeny of Chelicerata. Arthropod. Struct. Dev. 2010, 39, 124–142. [Google Scholar] [CrossRef]
- Waddington, J.; Rudkin, D.M.; Dunlop, J.A. A new mid-Silurian aquatic scorpion-one step closer to land? Biol. Lett. 2015, 11, 20140815. [Google Scholar] [CrossRef] [PubMed]
- Santibanez-Lopez, C.E.; Francke, O.F.; Ureta, C.; Possani, L.D. Scorpions from Mexico: From species diversity to venom complexity. Toxins 2015, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- The Scorpion Files. Available online: https://www.ntnu.no/ub/scorpion-files/intro.php (accessed on 20 March 2019).
- Zlotkin, E. Scorpion venoms. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier B.V.: Oxford, UK, 2005; Volume 5, pp. 173–220. [Google Scholar]
- Santibanez-Lopez, C.E.; Possani, L.D. Overview of the Knottin scorpion toxin-like peptides in scorpion venoms: Insights on their classification and evolution. Toxicon 2015, 107, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Almaaytah, A.; Albalas, Q. Scorpion venom peptides with no disulfide bridges: A review. Peptides 2014, 51, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Zhijian, C.; Feng, L.; Yingliang, W.; Xin, M.; Wenxin, L. Genetic mechanisms of scorpion venom peptide diversification. Toxicon 2006, 47, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Housley, D.M.; Housley, G.D.; Liddell, M.J.; Jennings, E.A. Scorpion toxin peptide action at the ion channel subunit level. Neuropharmacology 2017, 127, 46–78. [Google Scholar] [CrossRef]
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion toxins specific for Na+-channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef]
- Rodriguez de la Vega, R.C.; Possani, L.D. Overview of scorpion toxins specific for Na+ channels and related peptides: Biodiversity, structure-function relationships and evolution. Toxicon 2005, 46, 831–844. [Google Scholar] [CrossRef]
- Yu, F.H.; Catterall, W.A. Overview of the voltage-gated sodium channel family. Genome Biol. 2003, 4, 207. [Google Scholar] [CrossRef]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Cestele, S.; Stankiewicz, M.; Mansuelle, P.; De Waard, M.; Dargent, B.; Gilles, N.; Pelhate, M.; Rochat, H.; Martin-Eauclaire, M.F.; Gordon, D. Scorpion alpha-like toxins, toxic to both mammals and insects, differentially interact with receptor site 3 on voltage-gated sodium channels in mammals and insects. Eur. J. Neurosci. 1999, 11, 975–985. [Google Scholar] [CrossRef]
- Bergeron, Z.L.; Bingham, J.P. Scorpion toxins specific for potassium (K+) channels: A historical overview of peptide bioengineering. Toxins 2012, 4, 1082–1119. [Google Scholar] [CrossRef]
- Rodriguez de la Vega, R.C.; Possani, L.D. Current views on scorpion toxins specific for K+-channels. Toxicon 2004, 43, 865–875. [Google Scholar] [CrossRef]
- Chagot, B.; Pimentel, C.; Dai, L.; Pil, J.; Tytgat, J.; Nakajima, T.; Corzo, G.; Darbon, H.; Ferrat, G. An unusual fold for potassium channel blockers: NMR structure of three toxins from the scorpion Opisthacanthus madagascariensis. Biochem. J. 2005, 388, 263–271. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, F.; Feng, J.; Yang, W.; Zeng, D.; Zhao, R.; Cao, Z.; Liu, M.; Li, W.; Jiang, L.; et al. Genomic and structural characterization of Kunitz-type peptide LmKTT-1a highlights diversity and evolution of scorpion potassium channel toxins. PLoS ONE 2013, 8, e60201. [Google Scholar] [CrossRef]
- DeBin, J.A.; Maggio, J.E.; Strichartz, G.R. Purification and characterization of chlorotoxin, a chloride channel ligand from the venom of the scorpion. Am. J. Physiol. 1993, 264, C361–C369. [Google Scholar] [CrossRef]
- Froy, O.; Sagiv, T.; Poreh, M.; Urbach, D.; Zilberberg, N.; Gurevitz, M. Dynamic diversification from a putative common ancestor of scorpion toxins affecting sodium, potassium, and chloride channels. J. Mol. Evol. 1999, 48, 187–196. [Google Scholar] [CrossRef]
- Quintero-Hernandez, V.; Jimenez-Vargas, J.M.; Gurrola, G.B.; Valdivia, H.H.; Possani, L.D. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial peptides from scorpion venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef]
- Scorpiones.pl. Available online: http://scorpiones.pl/maps/#africa (accessed on 20 March 2019).
- Loret, E.P.; Hammock, B.D. Structure and neurotoxicity of venoms. In Scorpion Biology and Research; Brownell, P.H., Polis, G.A., Eds.; Oxford University Press, Inc.: New York, NY, USA, 2001; pp. 204–233. [Google Scholar]
- Caliskan, F.; Quintero-Hernandez, V.; Restano-Cassulini, R.; Batista, C.V.F.; Zarnudio, F.Z.; Coronas, F.I.; Possani, L.D. Turkish scorpion Buthacus macrocentrus: General characterization of the venom and description of Bu1, a potent mammalian Na+-channel alpha-toxin. Toxicon 2012, 59, 408–415. [Google Scholar] [CrossRef]
- Cestele, S.; Kopeyan, C.; Oughideni, R.; Mansuelle, P.; Granier, C.; Rochat, H. Biochemical and pharmacological characterization of a depressant insect toxin from the venom of the scorpion Buthacus arenicola. Eur. J. Biochem. 1997, 243, 93–99. [Google Scholar] [CrossRef]
- Miyashita, M.; Sakai, A.; Matsushita, N.; Hanai, Y.; Nakagawa, Y.; Miyagawa, H. A novel amphipathic linear peptide with both insect toxicity and antimicrobial activity from the venom of the scorpion Isometrus maculatus. Biosci. Biotechnol. Biochem. 2010, 74, 364–369. [Google Scholar] [CrossRef]
- Manzoil-Palma, M.F.; Gobbi, N.; Palma, M.S. Insects as biological models to assay spider and scorpion venom toxicity. Venom. Anim. Toxins incl. Trop. Dis. 2003, 9, 174–185. [Google Scholar] [CrossRef]
- De Lima, M.E.; Figueiredo, S.G.; Pimenta, A.M.; Santos, D.M.; Borges, M.H.; Cordeiro, M.N.; Richardson, M.; Oliveira, L.C.; Stankiewicz, M.; Pelhate, M. Peptides of arachnid venoms with insecticidal activity targeting sodium channels. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 264–279. [Google Scholar] [CrossRef]
- Gurevitz, M.; Karbat, I.; Cohen, L.; Ilan, N.; Kahn, R.; Turkov, M.; Stankiewicz, M.; Stuhmer, W.; Dong, K.; Gordon, D. The insecticidal potential of scorpion beta-toxins. Toxicon 2007, 49, 473–489. [Google Scholar] [CrossRef]
- Sautiere, P.; Cestele, S.; Kopeyan, C.; Martinage, A.; Drobecq, H.; Doljansky, Y.; Gordon, D. New toxins acting on sodium channels from the scorpion Leiurus quinquestriatus hebraeus suggest a clue to mammalian vs insect selectivity. Toxicon 1998, 36, 1141–1154. [Google Scholar] [CrossRef]
- Vargas, O.; Martin, M.F.; Rochat, H. Characterization of six toxins from the venom of the Moroccan scorpion Buthus occitanus mardochei. Eur. J. Biochem. 1987, 162, 589–599. [Google Scholar] [CrossRef]
- Kopeyan, C.; Martinez, G.; Rochat, H. Amino acid sequence of neurotoxin V from scorpion Leiurus quinquestriatus quinquestriatus. FEBS Lett. 1978, 89, 54–58. [Google Scholar] [CrossRef]
- Rochat, H.; Rochat, C.; Lissitzky, S.; Sampieri, F.; Miranda, F. Amino acid sequence of neurotoxin II of Androctonus australis Hector. Eur. J. Biochem. 1972, 28, 381–388. [Google Scholar] [CrossRef]
- Jalali, A.; Bosmans, F.; Amininasab, M.; Clynen, E.; Cuypers, E.; Zaremirakabadi, A.; Sarbolouki, M.N.; Schoofs, L.; Vatanpour, H.; Tytgat, J. OD1, the first toxin isolated from the venom of the scorpion Odonthobuthus doriae active on voltage-gated Na(+) channels. FEBS Lett. 2005, 579, 4181–4186. [Google Scholar] [CrossRef]
- Loret, E.P.; Mansuelle, P.; Rochat, H.; Granier, C. Neurotoxins active on insects—Amino acid sequences, chemical modifications, and secondary structure estimation by circular dichroism of toxins from the scorpion Androctonus australis Hector. Biochemistry 1990, 29, 1492–1501. [Google Scholar] [CrossRef]
- Kopeyan, C.; Mansuelle, P.; Sampieri, F.; Brando, T.; Bahraoui, E.M.; Rochat, H.; Granier, C. Primary structure of scorpion antiinsect toxins isolated from the venom of Leiurus quinquestriatus quinquestriatus. FEBS Lett. 1990, 261, 423–426. [Google Scholar] [CrossRef]
- Zhao, R.M.; Ma, Y.B.; He, Y.W.; Di, Z.Y.; Wu, Y.L.; Cao, Z.J.; Li, W.X. Comparative venom gland transcriptome analysis of the scorpion Lychas mucronatus reveals intraspecific toxic gene diversity and new venomous components. BMC Genom. 2010, 11. [Google Scholar] [CrossRef]
- Ma, Y.B.; He, Y.W.; Zhao, R.M.; Wu, Y.L.; Li, W.X.; Cao, Z.J. Extreme diversity of scorpion venom peptides and proteins revealed by transcriptomic analysis: Implication for proteome evolution of scorpion venom arsenal. J. Proteom. 2012, 75, 1563–1576. [Google Scholar] [CrossRef]
- Ichiki, Y.; Kawachi, T.; Miyashita, M.; Nakagawa, Y.; Miyagawa, H. Isolation and characterization of a novel non-selective beta-toxin from the venom of the scorpion Isometrus maculatus. Biosci. Biotechnol. Biochem. 2012, 76, 2089–2092. [Google Scholar] [CrossRef][Green Version]
- Kawachi, T.; Miyashita, M.; Nakagawa, Y.; Miyagawa, H. Isolation and characterization of an anti-insect beta-toxin from the venom of the scorpion Isometrus maculatus. Biosci. Biotechnol. Biochem. 2013, 77, 205–207. [Google Scholar] [CrossRef][Green Version]
- Karbat, I.; Kahn, R.; Cohen, L.; Ilan, N.; Gilles, N.; Corzo, G.; Froy, O.; Gur, M.; Albrecht, G.; Heinemann, S.H.; et al. The unique pharmacology of the scorpion alpha-like toxin Lqh3 is associated with its flexible C-tail. FEBS J. 2007, 274, 1918–1931. [Google Scholar] [CrossRef]
- Strohalm, M.; Kavan, D.; Novak, P.; Volny, M.; Havlicek, V. mMass 3: A cross-platform software environment for precise analysis of mass spectrometric data. Anal. Chem. 2010, 82, 4648–4651. [Google Scholar] [CrossRef]
- Ogata, K.; Umeyama, H. An automatic homology modeling method consisting of database searches and simulated annealing. J. Mol. Graph. Model. 2000, 18, 258–272. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | N-Terminal Sequence (U = Unknown) | Similar Peptides | Toxin Classification |
|---|---|---|---|
| Bl-1 | ARDGYISQPENCVYHCFPGS | Lqh3, Bom3 | α-like insect and mammal toxin |
| Bl-2 | URDGYLVDDUNCTFFCG | Lqh2, AaH2 | α-mammal toxin |
| Bl-3 | UVRDAYIADDKNCVYTCASN | OD1, Bu1 | α-insect and mammal toxin |
| Bl-4 | UKNGYAVDSSGKAPECILSNYCNNECTKV | AaHIT1, LqqIT1 | β-insect toxin |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshimoto, Y.; Miyashita, M.; Abdel-Wahab, M.; Sarhan, M.; Nakagawa, Y.; Miyagawa, H. Isolation and Characterization of Insecticidal Toxins from the Venom of the North African Scorpion, Buthacus leptochelys. Toxins 2019, 11, 236. https://doi.org/10.3390/toxins11040236
Yoshimoto Y, Miyashita M, Abdel-Wahab M, Sarhan M, Nakagawa Y, Miyagawa H. Isolation and Characterization of Insecticidal Toxins from the Venom of the North African Scorpion, Buthacus leptochelys. Toxins. 2019; 11(4):236. https://doi.org/10.3390/toxins11040236
Chicago/Turabian StyleYoshimoto, Yusuke, Masahiro Miyashita, Mohammed Abdel-Wahab, Moustafa Sarhan, Yoshiaki Nakagawa, and Hisashi Miyagawa. 2019. "Isolation and Characterization of Insecticidal Toxins from the Venom of the North African Scorpion, Buthacus leptochelys" Toxins 11, no. 4: 236. https://doi.org/10.3390/toxins11040236
APA StyleYoshimoto, Y., Miyashita, M., Abdel-Wahab, M., Sarhan, M., Nakagawa, Y., & Miyagawa, H. (2019). Isolation and Characterization of Insecticidal Toxins from the Venom of the North African Scorpion, Buthacus leptochelys. Toxins, 11(4), 236. https://doi.org/10.3390/toxins11040236