l-Proline Alleviates Kidney Injury Caused by AFB1 and AFM1 through Regulating Excessive Apoptosis of Kidney Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
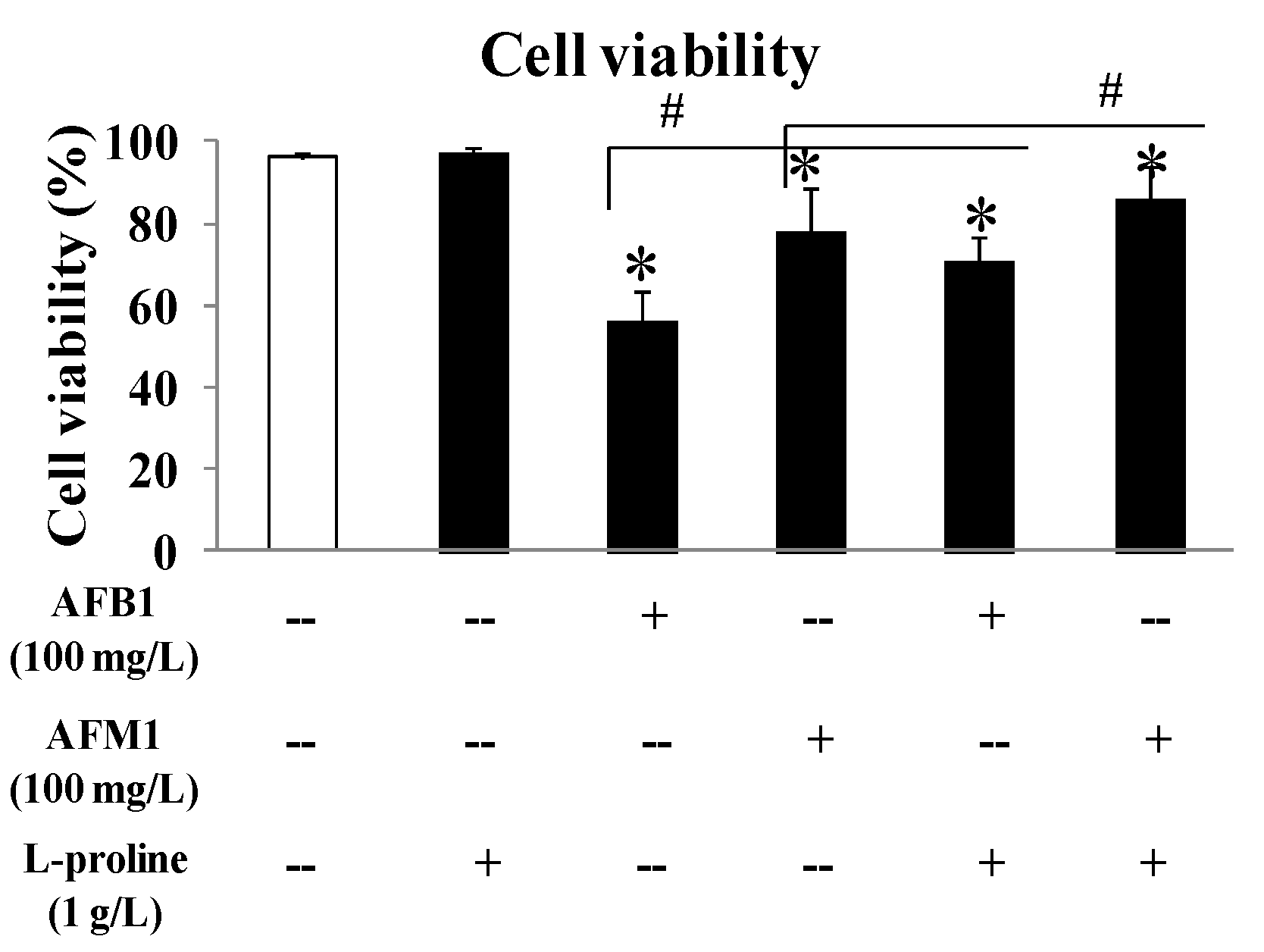
2.1. l-Proline Alleviates the Decreased Viability of HEK 293 cells Caused by AFB1 and AFM1
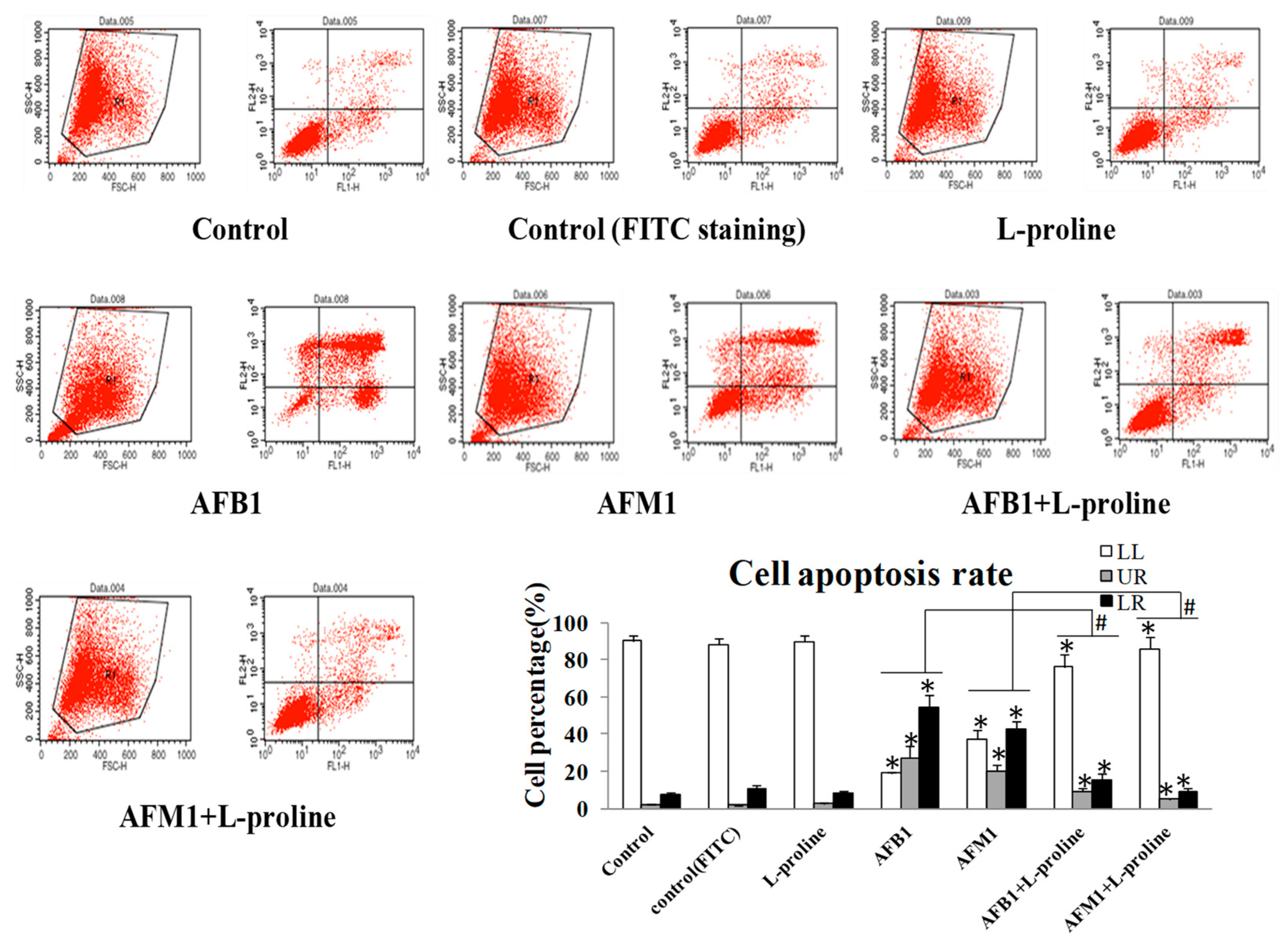
2.2. l-Proline Decreases Apoptosis Rate of HEK 293 Cells Induced by AFB1 and AFM1
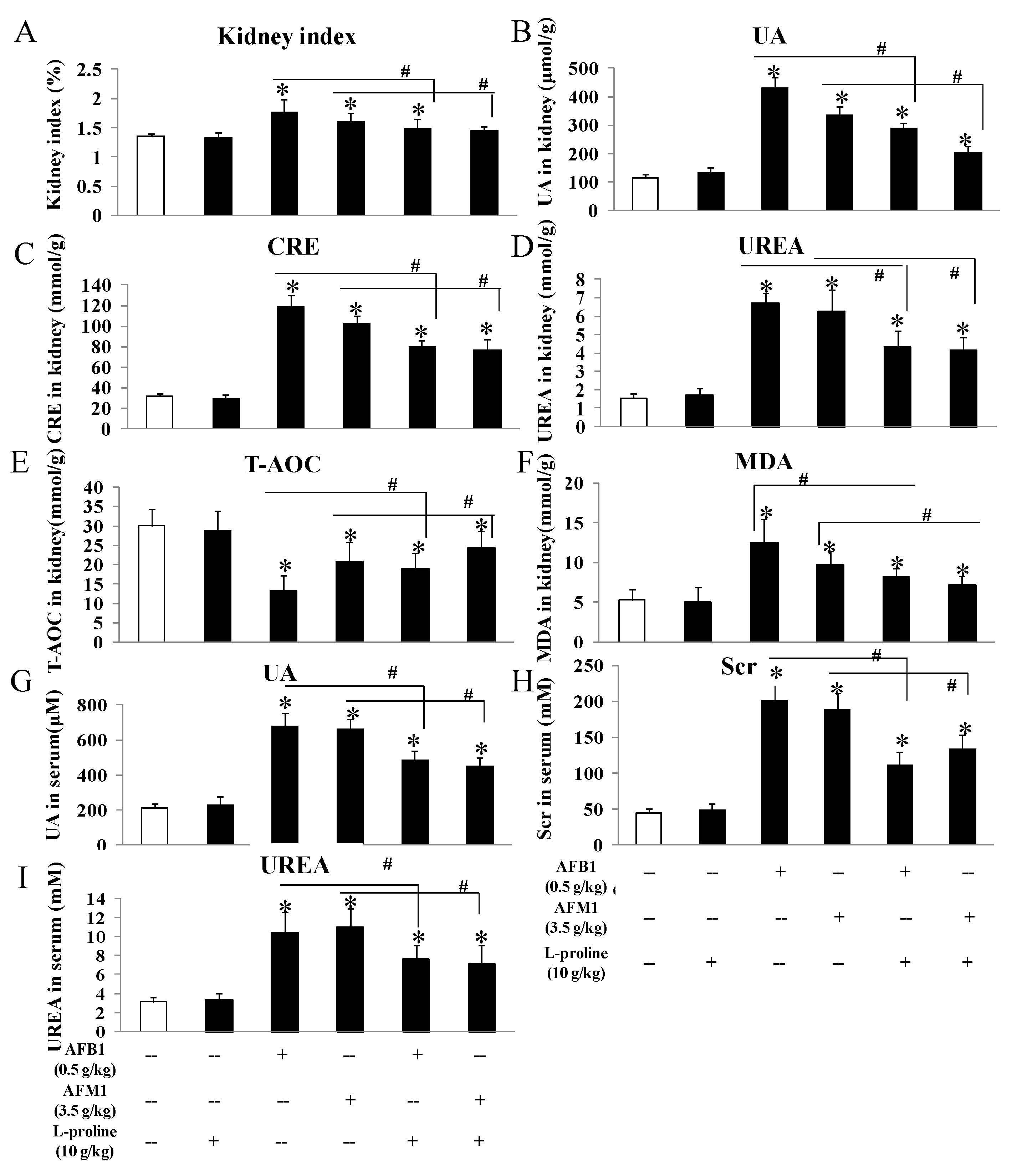
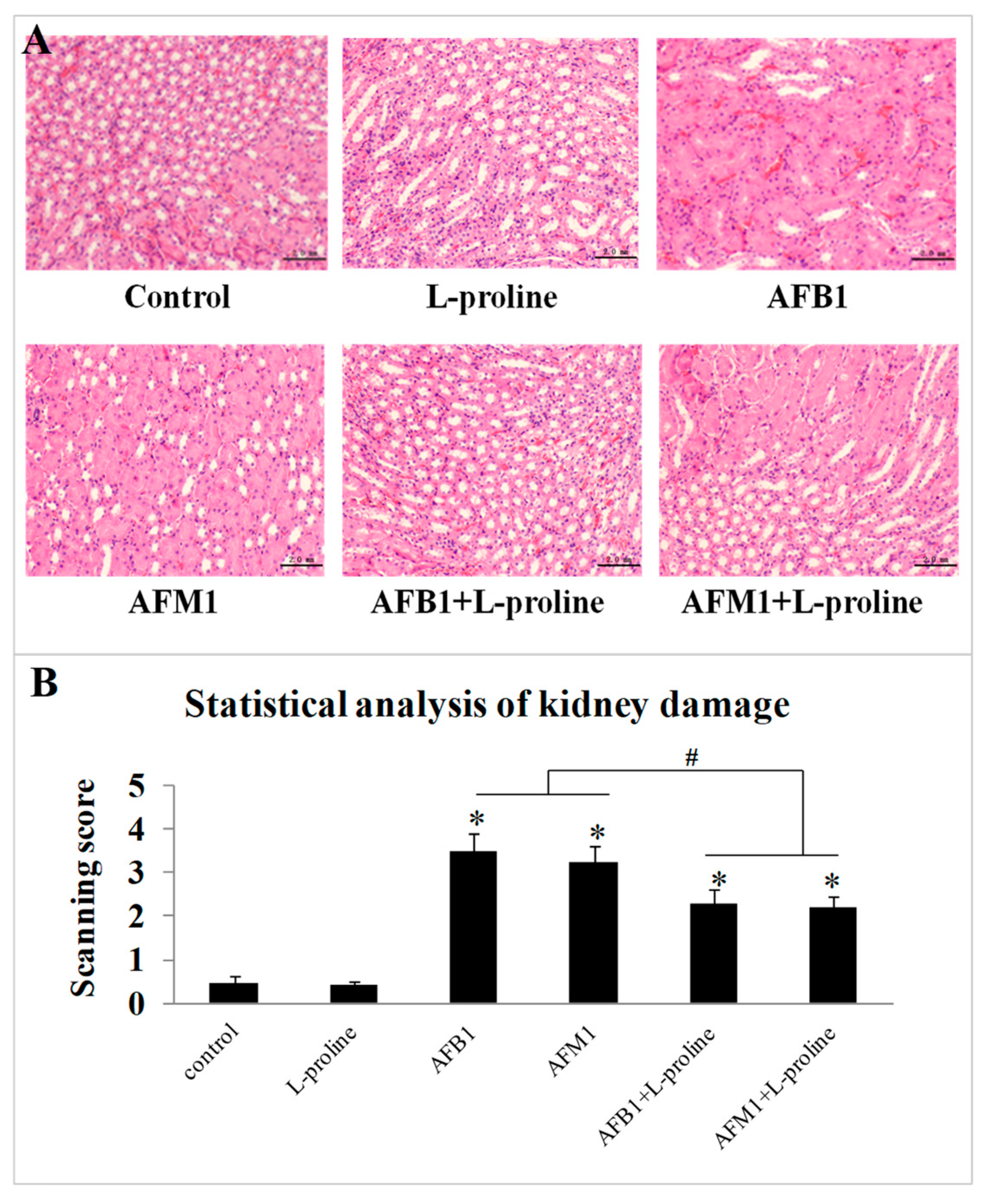
2.3. l-Proline Alleviates the Kidney Damage Caused by AFB1 and AFM1, Referring to Both Biochemical Parameters and Pathology Condition
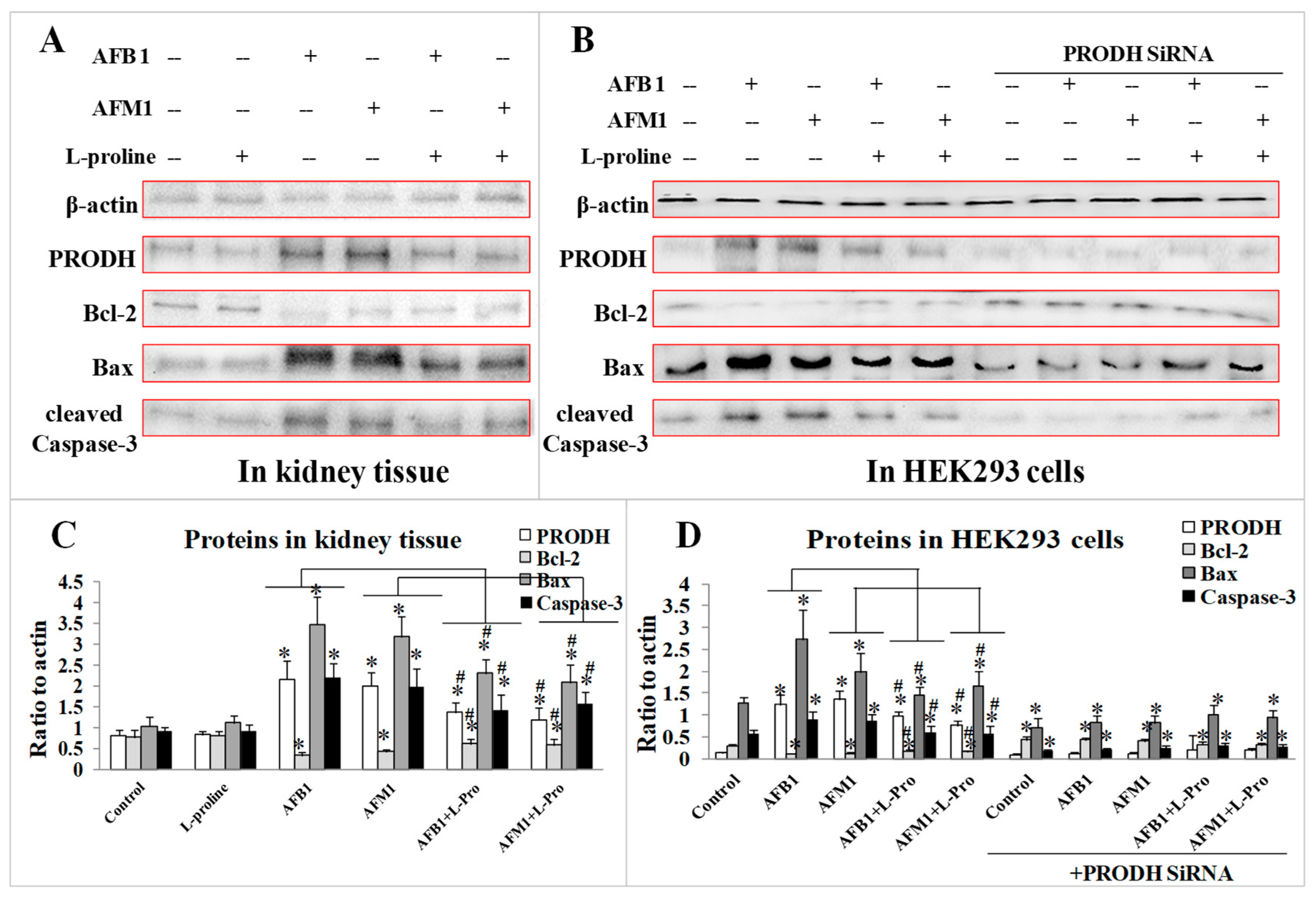
2.4. Validation of the Protective Effect of l-Proline in Regulating Apoptosis Factors via PRODH
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture, Viability Detection and SiRNA Treatment
4.3. Cell Apoptosis Detection
4.4. Animal Model
4.5. Histopathological Analyses
4.6. Biochemical Analysis
4.7. Western Blotting Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Battacone, G.; Nudda, A.; Palomba, M.; Pascale, M.; Nicolussi, P.; Pulina, G. Transfer of aflatoxin B1 from feed to milk and from milk to curd and whey in dairy sheep fed artificially contaminated concentrates. J. Dairy Sci. 2005, 88, 3063–3069. [Google Scholar] [CrossRef]
- Ronchi, B.; Danieli, P.; Vitali, A.; Sabatini, A.; Bernabucci, U.; Nardone, A. Evaluation of AFB1/AFM1 carry-over in lactating goats exposed to different levels of AFB1 contamination. In Book of Abstracts of the 56th Annual Meeting of the European Association for Animal Production, Uppsala, Sweden; European Association for Animal Production: Roma, Italy, 2005; pp. 1–10. [Google Scholar]
- Van Egmond, H.P.; Dragacci, S. Liquid chromatographic method for aflatoxin M1 in milk. Methods Mol. Biol. 2001, 157, 59–69. [Google Scholar]
- Bedard, L.L.; Massey, T.E. Aflatoxin B1-induced DNA damage and its repair. Cancer Lett. 2006, 241, 174–183. [Google Scholar] [CrossRef]
- Tchana, A.N.; Moundipa, P.F.; Tchouanguep, F.M. Aflatoxin contamination in food and body fluids in relation to malnutrition and cancer status in Cameroon. Int. J. Environ. Res. Public Health 2010, 7, 178–188. [Google Scholar] [CrossRef]
- Pong, R.S.; Wogan, G.N. Toxicity and biochemical and fine structural effects of synthetic aflatoxins M 1 and B 1 in rat liver. J. Natl. Cancer Inst. 1971, 47, 585–592. [Google Scholar] [PubMed]
- Chiavaro, E.; Cacchioli, C.; Berni, E.; Spotti, E. Immunoaffinity clean-up and direct fluorescence measurement of aflatoxins B1 and M1 in pig liver: Comparison with high-performance liquid chromatography determination. Food Addit. Contam. 2005, 22, 1154–1161. [Google Scholar] [CrossRef]
- Cui, X.; Muhammad, I.; Li, R.; Jin, H.; Guo, Z.; Yang, Y.; Hamid, S.; Li, J.; Cheng, P.; Zhang, X. Development of a UPLC-FLD Method for Detection of Aflatoxin B1 and M1 in Animal Tissue to Study the Effect of Curcumin on Mycotoxin Clearance Rates. Front. Pharmacol. 2017, 8, 650. [Google Scholar] [CrossRef]
- Hayes, R.B.; van Nieuwenhuize, J.P.; Raatgever, J.W.; ten Kate, F.J. Aflatoxin exposures in the industrial setting: An epidemiological study of mortality. Food Chem. Toxicol. 1984, 22, 39–43. [Google Scholar] [PubMed]
- Kelly, J.D.; Eaton, D.L.; Guengerich, F.P.; Coulombe, R.A., Jr. Aflatoxin B1 activation in human lung. Toxicol. Appl. Pharmacol. 1997, 144, 88–95. [Google Scholar] [CrossRef]
- Dvorackova, I.; Stora, C.; Ayraud, N. Evidence of aflatoxin B1 in two cases of lung cancer in man. J. Cancer Res. Clin. Oncol. 1981, 100, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Ibrahim, A.; El-Nekeety, A.A.; Hassan, N.S. Panax ginseng CA Meyer extract counteracts the oxidative stress in rats fed multi-mycotoxins-contaminated diet. Comun. Sci. 2012, 3, 143–153. [Google Scholar]
- Polychronaki, N.; Wild, C.P.; Mykkanen, H.; Amra, H.; Abdel-Wahhab, M.; Sylla, A.; Diallo, M.; EL-Nezami, H.; Turner, P.C. Urinary biomarkers of aflatoxin exposure in young children from Egypt and Guinea. Food Chem. Toxicol. 2008, 46, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.C.; Collinson, A.C.; Cheung, Y.B.; Gong, Y.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Aflatoxin exposure in utero causes growth faltering in Gambian infants. Int. J. Epidemiol. 2007, 36, 1119–1125. [Google Scholar] [CrossRef]
- Li, H.; Xing, L.; Zhang, M.; Wang, J.; Zheng, N. The Toxic Effects of Aflatoxin B1 and Aflatoxin M1 on Kidney through Regulating l-Proline and Downstream Apoptosis. Biomed. Res. Int. 2018, 2018, 9074861. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tang, L.; Glenn, T.C.; Wang, J.S. Aflatoxin B1 Induced Compositional Changes in Gut Microbial Communities of Male F344 Rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef]
- Zhang, Y.; Stefanovic, B. Akt mediated phosphorylation of LARP6; critical step in biosynthesis of type I collagen. Sci. Rep. 2016, 6, 22597. [Google Scholar] [CrossRef]
- Madan, E.; Gogna, R.; Pati, U. p53 Ser15 phosphorylation disrupts the p53-RPA70 complex and induces RPA70-mediated DNA repair in hypoxia. Biochem. J. 2012, 443, 811–820. [Google Scholar] [CrossRef]
- Sabino, M.; Purchio, A.; Milanez, T.V. Aflatoxins B1, M1 and aflatoxicol in tissues and urine of calves receiving aflatoxin. Food Addit. Contam. 1995, 12, 467–472. [Google Scholar] [CrossRef]
- Ribichini, F.; Graziani, M.; Gambaro, G.; Pasoli, P.; Pighi, M.; Pesarini, G.; Brunelleschi, T.S.; Rizzotti, P.; Lupo, A.; Vassanelli, C. Early creatinine shifts predict contrast-induced nephropathy and persistent renal damage after angiography. Am. J. Med. 2010, 123, 755–763. [Google Scholar] [CrossRef]
- Jolly, P.; Jiang, Y.; Ellis, W.; Awuah, R.; Nnedu, O.; Phillips, T.; Wang, J.S.; Afriyie-Gyawu, E.; Tang, L.L.; Person, S.; et al. Determinants of aflatoxin levels in Ghanaians: Sociodemographic factors, knowledge of aflatoxin and food handling and consumption practices. Int. J. Hyg. Environ. Health 2006, 209, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Alpsoy, L.; Yildirim, A.; Agar, G. The antioxidant effects of vitamin A, C, and E on aflatoxin B1-induced oxidative stress in human lymphocytes. Toxicol. Ind. Health 2009, 25, 121–127. [Google Scholar] [CrossRef]
- Wang, L. The significance of Cys-C UREA and Scr tests in early renal damage assessment of acute glomerulonephritis. Labeled Immunoass. Clin. Med. 2017, 4, 422–424. [Google Scholar]
- Olearczyk, J.J.; Quigley, J.E.; Mitchell, B.C.; Yamamoto, T.; Kim, I.H.; Newman, J.W.; Ayala, L.; Bruce, D.H.; John, D.I. Administration of a substituted adamantyl urea inhibitor of soluble epoxide hydrolase protects the kidney from damage in hypertensive Goto-Kakizaki rats. Clin. Sci. (Lond.) 2009, 116, 61–70. [Google Scholar] [CrossRef]
- Liu, C.L.; Shu-Bao, L.I. Correlation of serum uric acid and early renal damage in patients with type 2 diabetes mellitus. Anhui Med. Pharm. J. 2012, 27, 237–239. [Google Scholar]
- Kwak, H.S.; Lee, Y.H.; Han, Y.M.; Jin, G.Y.; Kim, W.; Chung, G.H. Comparison of renal damage by iodinated contrast or gadolinium in an acute renal failure rat model based on serum creatinine levels and apoptosis degree. J. Korean Med. Sci. 2005, 20, 841–847. [Google Scholar] [CrossRef]
- Liu, S.X.; Li, C.G.; Dai, D.L. Effect of ligustrazine on MDA, SOD and T-AOC in erythrocyte membrane in patients with hemoglobin H disease. J. Clin. Exp. Med. 2010, 11. [Google Scholar]
- Kanski, J.; Aksenova, M.; Schoneich, C.; Butterfield, D.A. Substitution of isoleucine-31 by helical-breaking proline abolishes oxidative stress and neurotoxic properties of Alzheimer’s amyloid beta-peptide. Free Radic. Biol. Med. 2002, 32, 1205–1211. [Google Scholar] [CrossRef]
- Xiang, Y.; Chen, G.; Wang, S.J.; Fang, Y.J.; Cai, R.J.; Deng, Q.K. The effect of various pulse amplitudes on the amount of SOD, MDA, and T-AOC in the diaphagm of rabbit atter diaphragm pacing. Chinese J. Phys. Med. Rehabil. 2004, 26, 269–271. [Google Scholar]
- Son, Y.O.; Hitron, J.A.; Shi, X. Chromium (VI), Oxidative Cell Damage; Springer: New York, NY, USA, 2013. [Google Scholar]
- Tripathi, B.N.; Gaur, J.P. Relationship between copper- and zinc-induced oxidative stress and proline accumulation in Scenedesmus sp. Planta 2004, 219, 397–404. [Google Scholar] [CrossRef]
- Khan, M.I.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant. Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef]
- Chen, C.; Wanduragala, S.; Becker, D.F.; Dickman, M.B. Tomato QM-like protein protects Saccharomyces cerevisiae cells against oxidative stress by regulating intracellular proline levels. Appl. Environ. Microbiol. 2006, 72, 4001–4006. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef]
- Sumithra, K.; Jutur, P.P.; Carmel, B.D.; Reddy, A.R. Salinity-induced changes in two cultivars of Vigna radiata: Responses of antioxidative and proline metabolism. J. Plant Growth Regul. 2006, 50, 11–22. [Google Scholar] [CrossRef]
- Zhang, L.; Alfano, J.R.; Becker, D.F. Proline metabolism increases katG expression and oxidative stress resistance in Escherichia coli. J. Bacteriol. 2015, 197, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; Schuermann, J.P.; White, T.A.; Krishnan, N.; Sanyal, N.; Hura, G.L.; Anmin, T.; Michael, T.H.; Donald, F.B.; John, J.T. Crystal structure of the bifunctional proline utilization A flavoenzyme from Bradyrhizobium japonicum. Proc. Natl. Acad. Sci. USA 2010, 107, 2878–2883. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Li, S.; Yang, H.; Wang, Y.; Wang, J.; Zheng, N. l-Proline Alleviates Kidney Injury Caused by AFB1 and AFM1 through Regulating Excessive Apoptosis of Kidney Cells. Toxins 2019, 11, 226. https://doi.org/10.3390/toxins11040226
Li H, Li S, Yang H, Wang Y, Wang J, Zheng N. l-Proline Alleviates Kidney Injury Caused by AFB1 and AFM1 through Regulating Excessive Apoptosis of Kidney Cells. Toxins. 2019; 11(4):226. https://doi.org/10.3390/toxins11040226
Chicago/Turabian StyleLi, Huiying, Songli Li, Huaigu Yang, Yizhen Wang, Jiaqi Wang, and Nan Zheng. 2019. "l-Proline Alleviates Kidney Injury Caused by AFB1 and AFM1 through Regulating Excessive Apoptosis of Kidney Cells" Toxins 11, no. 4: 226. https://doi.org/10.3390/toxins11040226
APA StyleLi, H., Li, S., Yang, H., Wang, Y., Wang, J., & Zheng, N. (2019). l-Proline Alleviates Kidney Injury Caused by AFB1 and AFM1 through Regulating Excessive Apoptosis of Kidney Cells. Toxins, 11(4), 226. https://doi.org/10.3390/toxins11040226