Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response




 ,
,  and
and 
Abstract
1. Introduction
2. Results
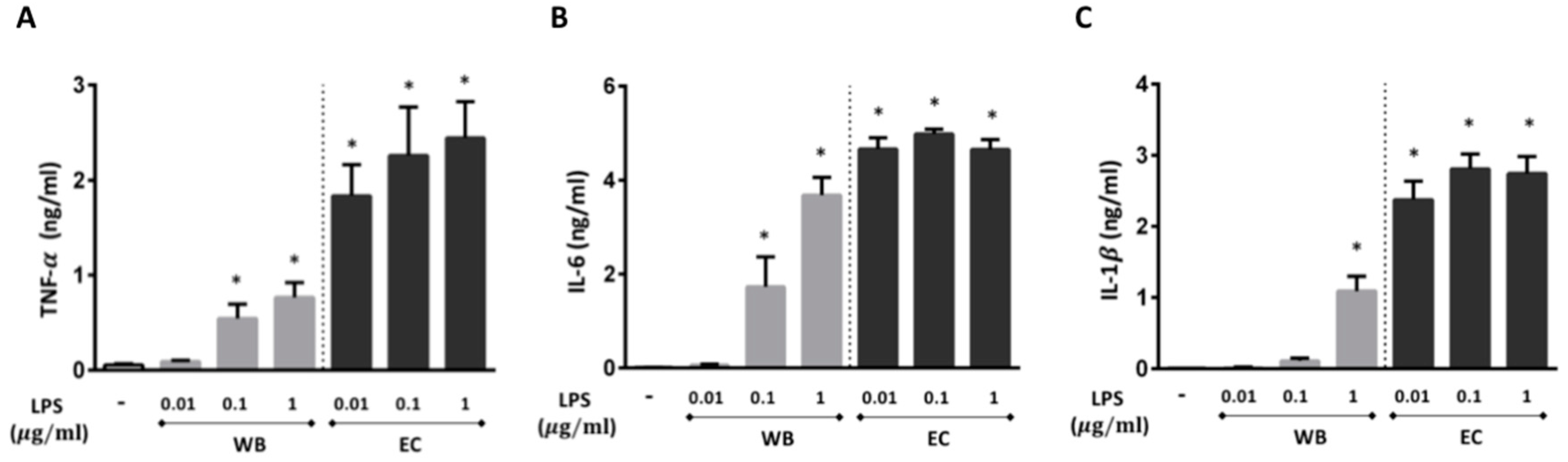
2.1. LPS Isolated from M. aeruginosa Water Bloom Induces the Production of Pro-Inflammatory Cytokines
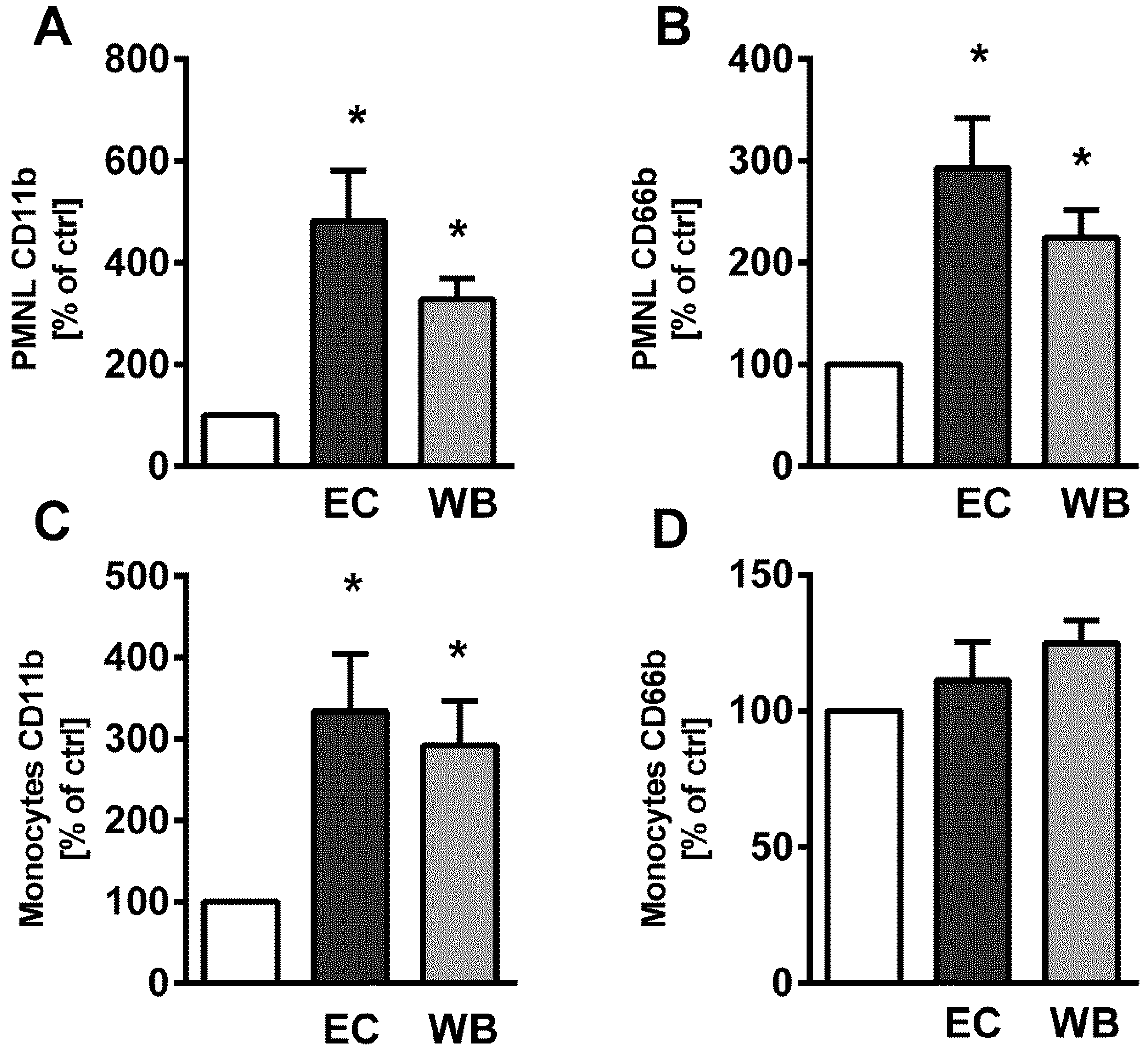
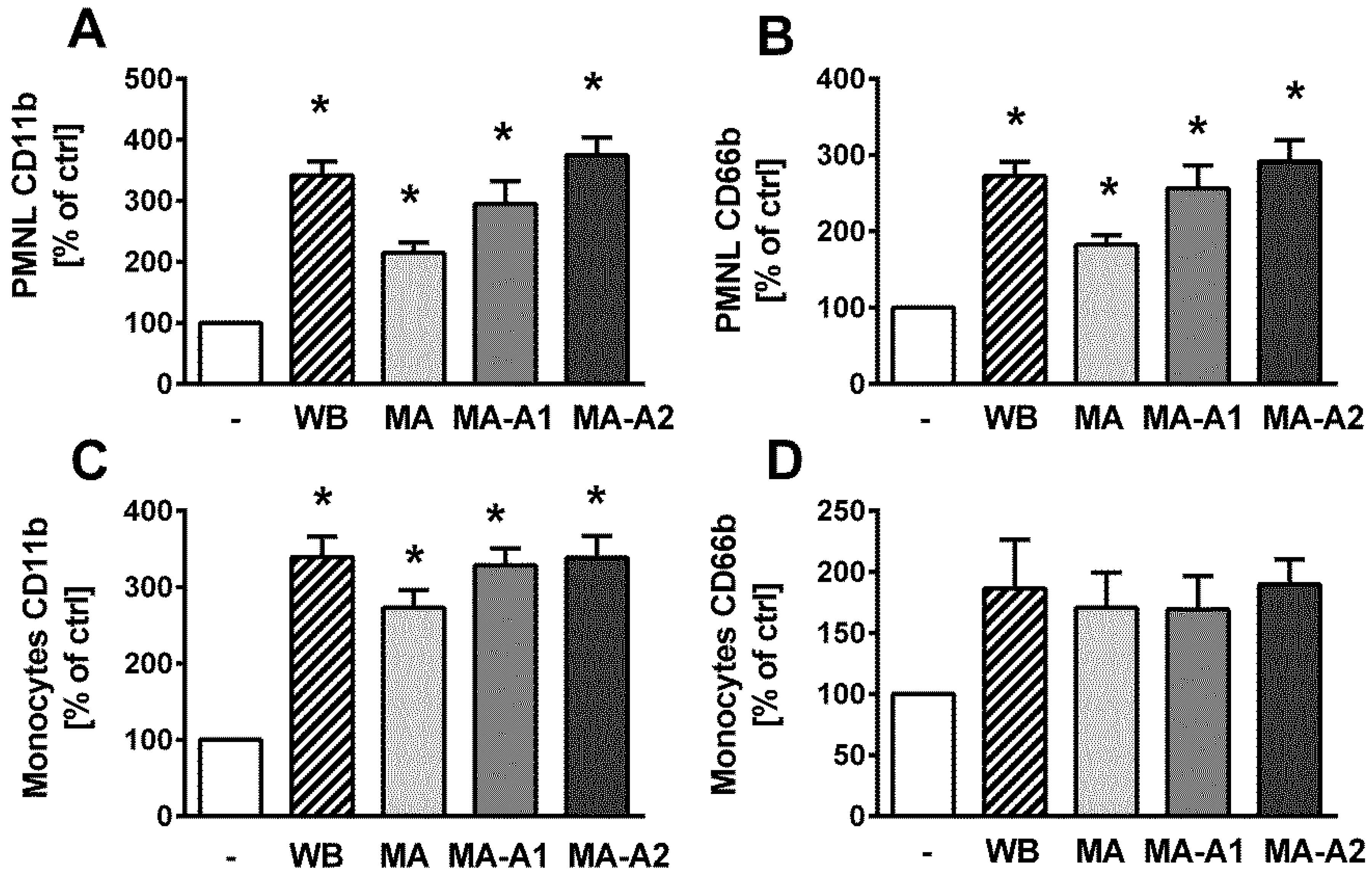
2.2. LPS Isolated from M. aeruginosa Water Bloom Activates Blood Phagocytes
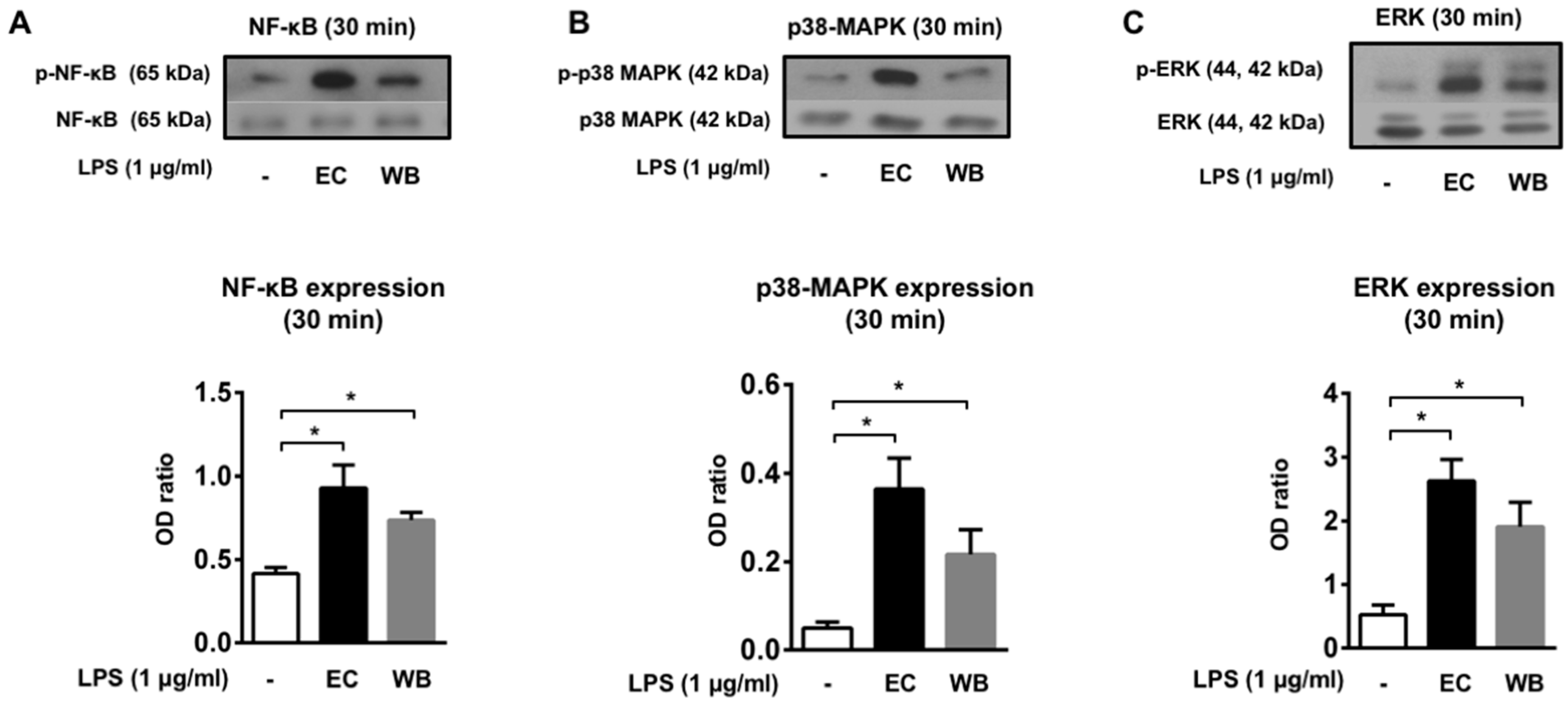
2.3. LPS Isolated from M. aeruginosa Water Bloom Activates Selected Signaling Pathways
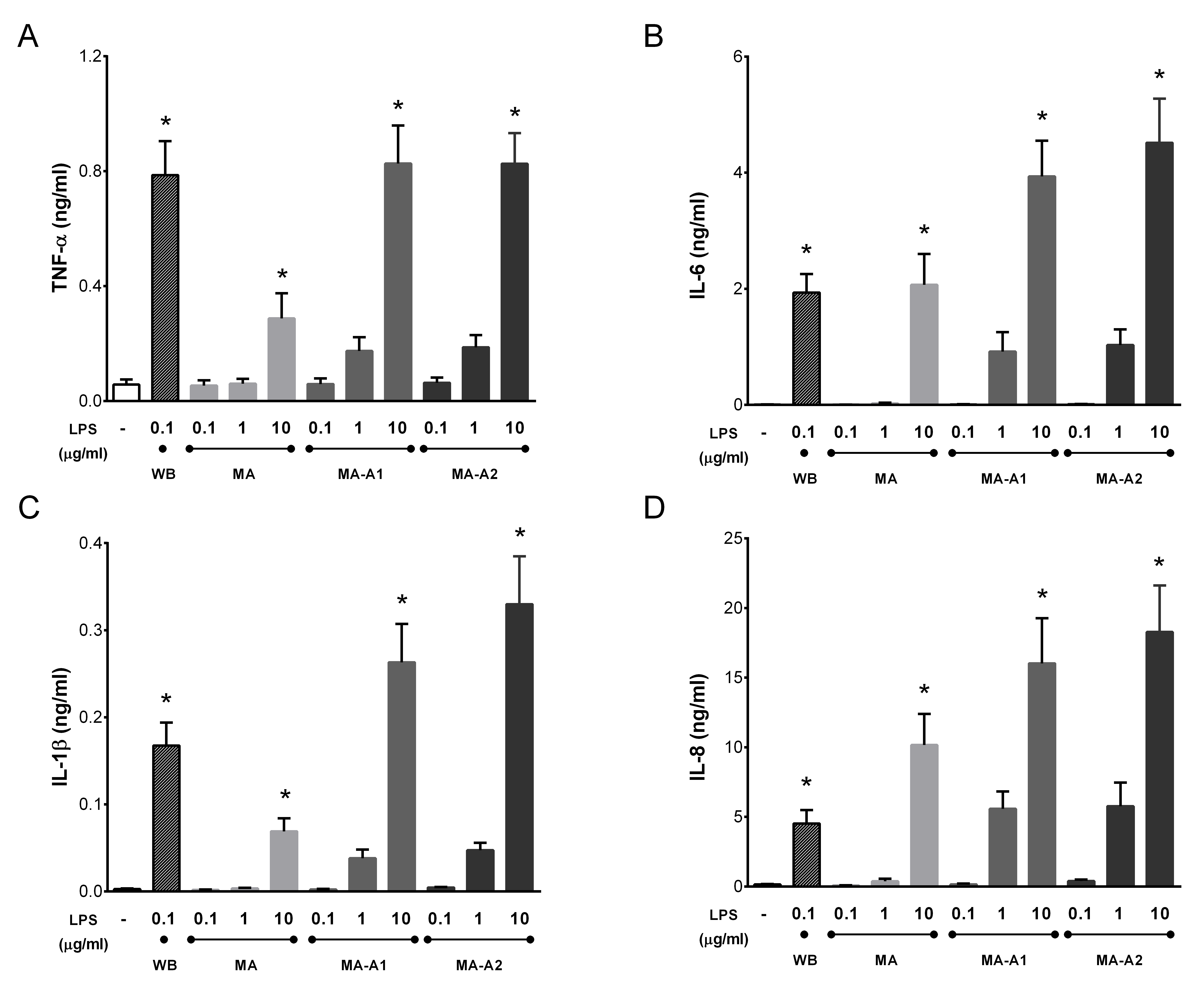
2.4. Cyanobacterial LPS Alone also Activates Blood Phagocytes
2.5. Pyrogenic Activity of Tested LPSs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Biomass Preparation
5.2. LPS Preparation
5.3. Pyrogenicity by the PyroGeneTM Recombinant Factor C Endpoint Fluorescent Assay
5.4. Preparation of Whole Blood Samples and Experimental Layout
5.5. Determination of Surface Expression of Receptors by Flow Cytometry (FC)
5.6. Detection of Cytokines by ELISA
5.7. Protein Analysis by Western Blot
5.8. Statistical Analysis of Data
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Blaha, L.; Babica, P.; Marsalek, B. Toxins produced in cyanobacterial water blooms—Toxicity and risks. Interdiscip. Toxicol. 2009, 2, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Svircev, Z.; Drobac, D.; Tokodi, N.; Mijovic, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Bourke, A.T.C.; Hawes, R.B.; Nelson, A.; Stallman, N.D. An outbreak of hepato-enteritis (the palm island mystery disease) possibly caused by algal intoxication. Toxicon 1983, 21, 45–48. [Google Scholar] [CrossRef]
- Griffiths, D.J.; Saker, M.L. The palm island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Anadotter, H.; Cronberg, G.; Lawton, L.A.; Hansson, H.-B.; Gothe, U.; Skulberg, O.M. An Extensive Outbreak of Gastroenteritis Associated with the Toxic Vyanobacterium Planktothrix Agardhii (Oscillatoriales, Cyanophyceae) in scania, South Sweden. In Cyanotoxins, Occurence, Causes, Conseqquences; Chorus, I., Ed.; Springer: Berlin, Germany, 2001. [Google Scholar]
- Hayman, J. Beyond the barcoo—Probable human tropical cyanobacterial poisoning in outback Australia. Med. J. Aust. 1992, 157, 794–796. [Google Scholar]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health Glob. Access Sci. Source 2006, 5, 7. [Google Scholar] [CrossRef]
- Durai, P.; Batool, M.; Choi, S. Structure and effects of cyanobacterial lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Billings, W.H. Water-Associated Human Illness in Northeast Pennsylvania and Its Suspected Association with Blue-Green Algae Blooms; Springer: New York, NY, USA, 1981. [Google Scholar]
- Dillenberg, H.O.; Dehnel, M.K. Toxic waterbloom in saskatchewan, 1959. Can. Med. Assoc. J. 1960, 83, 1151–1154. [Google Scholar]
- Rapala, J.; Robertson, A.; Negri, A.P.; Berg, K.A.; Tuomi, P.; Lyra, C.; Erkomaa, K.; Lahti, K.; Hoppu, K.; Lepisto, L. First report of saxitoxin in finnish lakes and possible associated effects on human health. Environ. Toxicol. 2005, 20, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.C.; Gammie, A.J.; Hollinrake, K.; Codd, G.A. Pneumonia associated with contact with cyanobacteria. BMJ 1990, 300, 1440–1441. [Google Scholar] [CrossRef] [PubMed]
- Adamovsky, O.; Moosova, Z.; Pekarova, M.; Basu, A.; Babica, P.; Svihalkova Sindlerova, L.; Kubala, L.; Blaha, L. Immunomodulatory potency of microcystin, an important water-polluting cyanobacterial toxin. Environ. Sci. Technol. 2015, 49, 12457–12464. [Google Scholar] [CrossRef] [PubMed]
- McLellan, N.L.; Manderville, R.A. Toxic mechanisms of microcystins in mammals. Toxicol. Res. 2017, 6, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Macagno, A.; Molteni, M.; Rinaldi, A.; Bertoni, F.; Lanzavecchia, A.; Rossetti, C.; Sallusto, F. A cyanobacterial LPS antagonist prevents endotoxin shock and blocks sustained TLR4 stimulation required for cytokine expression. J. Exp. Med. 2006, 203, 1481–1492. [Google Scholar] [CrossRef]
- Matiasovic, J.; Stepanova, H.; Volf, J.; Kubala, L.; Ovesna, P.; Rychlik, I.; Faldyna, M. Influence of the lipopolysaccharide structure of salmonella enterica serovar enteritidis on interactions with pig neutrophils. Vet. Microbiol. 2011, 150, 167–172. [Google Scholar] [CrossRef]
- Fujii, M.; Sato, Y.; Ito, H.; Masago, Y.; Omura, T. Monosaccharide composition of the outer membrane lipopolysaccharide and o-chain from the freshwater cyanobacterium microcystis aeruginosa nies-87. J. Appl. Microbiol. 2012, 113, 896–903. [Google Scholar] [CrossRef]
- Jurgens, U.J.; Martin, C.; Weckesser, J. Cell wall constituents of microcystis sp. Pcc 7806. FEMS Microbiol. Lett. 1989, 53, 47–51. [Google Scholar] [CrossRef]
- Martin, C.; Codd, G.A.; Siegelman, H.W.; Weckesser, J. Lipopolysaccharides and polysaccharides of the cell-envelope of toxic microcystis-aeruginosa strains. Arch. Microbiol. 1989, 152, 90–94. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Karima, R.; Matsumoto, S.; Higashi, H.; Matsushima, K. The molecular pathogenesis of endotoxic shock and organ failure. Mol. Med. Today 1999, 5, 123–132. [Google Scholar] [CrossRef]
- Pop-Began, V.; Paunescu, V.; Grigorean, V.; Pop-Began, D.; Popescu, C. Molecular mechanisms in the pathogenesis of sepsis. J. Med. Life 2014, 7, 38–41. [Google Scholar]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Vogt, K.L.; Summers, C.; Chilvers, E.R.; Condliffe, A.M. Priming and de-priming of neutrophil responses in vitro and in vivo. Eur. J. Clin. Investig. 2018, 48, e12967. [Google Scholar] [CrossRef]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef]
- Shiraishi, H. Association of heterotrophic bacteria with aggregated arthrospira platensis exopolysaccharides: Implications in the induction of axenic cultures. Biosci. Biotechnol. Biochem. 2015, 79, 331–341. [Google Scholar] [CrossRef]
- Ueda, H.; Otsuka, S.; Senoo, K. Community composition of bacteria co-cultivated with microalgae in non-axenic algal cultures. Microbiol. Cult. Coll. 2009, 25, 21–25. [Google Scholar]
- Blahova, L.; Adamovsky, O.; Kubala, L.; Svihalkova Sindlerova, L.; Zounkova, R.; Blaha, L. The isolation and characterization of lipopolysaccharides from microcystis aeruginosa, a prominent toxic water bloom forming cyanobacteria. Toxicon 2013, 76, 187–196. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Gantar, M.; Mayer, G.D.; Gibbs, P.D.; Berry, J.P. Effects of cyanobacterial lipopolysaccharides from microcystis on glutathione-based detoxification pathways in the zebrafish (danio rerio) embryo. Toxins 2012, 4, 390–404. [Google Scholar] [CrossRef]
- Mayer, A.M.; Clifford, J.A.; Aldulescu, M.; Frenkel, J.A.; Holland, M.A.; Hall, M.L.; Glaser, K.B.; Berry, J. Cyanobacterial microcystis aeruginosa lipopolysaccharide elicits release of superoxide anion, thromboxane B(2), cytokines, chemokines, and matrix metalloproteinase-9 by rat microglia. Toxicol. Sci. 2011, 121, 63–72. [Google Scholar] [CrossRef]
- Mayer, A.M.; Murphy, J.; MacAdam, D.; Osterbauer, C.; Baseer, I.; Hall, M.L.; Feher, D.; Williams, P. Classical and alternative activation of cyanobacterium oscillatoria sp. Lipopolysaccharide-treated rat microglia in vitro. Toxicol. Sci. 2016, 149, 484–495. [Google Scholar] [CrossRef]
- Ohkouchi, Y.; Tajima, S.; Nomura, M.; Itoh, S. Inflammatory responses and potencies of various lipopolysaccharides from bacteria and cyanobacteria in aquatic environments and water supply systems. Toxicon 2015, 97, 23–31. [Google Scholar] [CrossRef]
- Swanson-Mungerson, M.; Incrocci, R.; Subramaniam, V.; Williams, P.; Hall, M.L.; Mayer, A.M.S. Effects of cyanobacteria oscillatoria sp. Lipopolysaccharide on b cell activation and toll-like receptor 4 signaling. Toxicol. Lett. 2017, 275, 101–107. [Google Scholar] [CrossRef]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef]
- Eiler, A.; Bertilsson, S. Composition of freshwater bacterial communities associated with cyanobacterial blooms in four swedish lakes. Environ. Microbiol. 2004, 6, 1228–1243. [Google Scholar] [CrossRef]
- Coats, S.R.; Do, C.T.; Karimi-Naser, L.M.; Braham, P.H.; Darveau, R.P. Antagonistic lipopolysaccharides block E. Coli lipopolysaccharide function at human tlr4 via interaction with the human md-2 lipopolysaccharide binding site. Cell Microbiol. 2007, 9, 1191–1202. [Google Scholar] [CrossRef]
- Coats, S.R.; Pham, T.T.; Bainbridge, B.W.; Reife, R.A.; Darveau, R.P. Md-2 mediates the ability of tetra-acylated and penta-acylated lipopolysaccharides to antagonize escherichia coli lipopolysaccharide at the tlr4 signaling complex. J. Immunol. 2005, 175, 4490–4498. [Google Scholar] [CrossRef]
- Ernst, R.K.; Adams, K.N.; Moskowitz, S.M.; Kraig, G.M.; Kawasaki, K.; Stead, C.M.; Trent, M.S.; Miller, S.I. The pseudomonas aeruginosa lipid a deacylase: Selection for expression and loss within the cystic fibrosis airway. J. Bacteriol. 2006, 188, 191–201. [Google Scholar] [CrossRef]
- Suomalainen, M.; Lobo, L.A.; Brandenburg, K.; Lindner, B.; Virkola, R.; Knirel, Y.A.; Anisimov, A.P.; Holst, O.; Korhonen, T.K. Temperature-induced changes in the lipopolysaccharide of yersinia pestis affect plasminogen activation by the pla surface protease. Infect. Immun. 2010, 78, 2644–2652. [Google Scholar] [CrossRef] [PubMed]
- Bernardova, K.; Babica, P.; Marsalek, B.; Blaha, L. Isolation and endotoxin activities of lipopolysaccharides from cyanobacterial cultures and complex water blooms and comparison with the effects of heterotrophic bacteria and green alga. J. Appl. Toxicol. 2008, 28, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Dehus, O.; Hartung, T.; Hermann, C. Endotoxin evaluation of eleven lipopolysaccharides by whole blood assay does not always correlate with limulus amebocyte lysate assay. J. Endotoxin Res. 2006, 12, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Bartova, K.; Hilscherova, K.; Babica, P.; Marsalek, B.; Blaha, L. Effects of microcystin and complex cyanobacterial samples on the growth and oxidative stress parameters in green alga pseudokirchneriella subcapitata and comparison with the model oxidative stressor—Herbicide paraquat. Environ. Toxicol. 2011, 26, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar]
- Blaha, L.; Blahova, L.; Kovalova, L.; Adamovsky, O.; Babica, P.; Marsalek, B. Temporal and spatial variability of cyanobacterial toxins microcystins in three interconnected freshwater reservoirs. J. Serb. Chem. Soc. 2010, 75, 1303–1312. [Google Scholar] [CrossRef]
- Javůrek, J. Retinoid compounds associated with water bloomsdominated by microcystis species. Harmful Algae 2015, 47, 116–125. [Google Scholar] [CrossRef]
- Bláhová, L.; Babica, P.; Maršálková, E.; Maršálek, B.; Bláha, L. Concentrations and seasonal trends of extracellular microcystins in freshwaters of the czech republic—Results of the national monitoring program. CLEAN Soil Air Water 2007, 35, 348–354. [Google Scholar] [CrossRef]
- Jancula, D.; Strakova, L.; Sadilek, J.; Marsalek, B.; Babica, P. Survey of cyanobacterial toxins in czech water reservoirs—The first observation of neurotoxic saxitoxins. Environ. Sci. Pollut. Res. Int. 2014, 21, 8006–8015. [Google Scholar] [CrossRef]
- Jasa, L.; Sadilek, J.; Kohoutek, J.; Strakova, L.; Marsalek, B.; Babica, P. Application of passive sampling for sensitive time-integrative monitoring of cyanobacterial toxins microcystins in drinking water treatment plants. Water Res. 2019, 153, 108–120. [Google Scholar] [CrossRef]
- Znachor, P.; Jurczak, T.; Komarkova, J.; Jezberova, J.; Mankiewicz, J.; Kastovska, K.; Zapomelova, E. Summer changes in cyanobacterial bloom composition and microcystin concentration in eutrophic czech reservoirs. Environ. Toxicol. 2006, 21, 236–243. [Google Scholar] [CrossRef]
- Kolarova, H.; Klinke, A.; Kremserova, S.; Adam, M.; Pekarova, M.; Baldus, S.; Eiserich, J.P.; Kubala, L. Myeloperoxidase induces the priming of platelets. Free Radic. Biol. Med. 2013, 61, 357–369. [Google Scholar] [CrossRef]
- Gallova, L.; Kubala, L.; Ciz, M.; Lojek, A. Il-10 does not affect oxidative burst and expression of selected surface antigen on human blood phagocytes in vitro. Physiol. Res. 2004, 53, 199–208. [Google Scholar]
- Pekarova, M.; Koudelka, A.; Kolarova, H.; Ambrozova, G.; Klinke, A.; Cerna, A.; Kadlec, J.; Trundova, M.; Sindlerova Svihalkova, L.; Kuchta, R.; et al. Asymmetric dimethyl arginine induces pulmonary vascular dysfunction via activation of signal transducer and activator of transcription 3 and stabilization of hypoxia-inducible factor 1-alpha. Vasc. Pharmacol. 2015, 73, 138–148. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LPS Sample | Label | PyroGene (×104 EU/mg LPS) |
|---|---|---|
| E. coli (serotype O26:B6) | EC | 674 |
| Environmental water bloom sample | WB | 1210 |
| Non-axenic M. aeruginosa PCC 7806 | MA | 108 |
| Axenic M. aeruginosa PCC 7806 | MA-A1 | <1 |
| Axenic M. aeruginosa HAMBI/UHCC 130 | MA-A2 | <1 |
| Locality | Nové Mlýny I, Czech Republic |
|---|---|
| Sampling Site | 48°53′9.155″N, 16°35′41.663″E |
| Sampling Date | 14 August 2012 |
| Total phytoplankton concentration (cell/mL) | 10.2 × 106 |
| Percentage of cyanobacteria in total biomass (% of the total cell count) | 99.8 |
| Species composition (% of cyanobacterial cell count) | M. aeruginosa (100) |
| The concentration of major microcystins (MCs) in phytoplankton biomass (µg/g d.w.) | MC-RR: 329 MC-YR: 83 MC-LR: 373 Sum MCs: 785 |
| The concentration of major MCs in water (µg/L) | MC-RR: 2.22 MC-YR: 0.68 MC-LR: 1.29 Sum MCs: 4.19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moosová, Z.; Šindlerová, L.; Ambrůzová, B.; Ambrožová, G.; Vašíček, O.; Velki, M.; Babica, P.; Kubala, L. Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response. Toxins 2019, 11, 218. https://doi.org/10.3390/toxins11040218
Moosová Z, Šindlerová L, Ambrůzová B, Ambrožová G, Vašíček O, Velki M, Babica P, Kubala L. Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response. Toxins. 2019; 11(4):218. https://doi.org/10.3390/toxins11040218
Chicago/Turabian StyleMoosová, Zdena, Lenka Šindlerová, Barbora Ambrůzová, Gabriela Ambrožová, Ondřej Vašíček, Mirna Velki, Pavel Babica, and Lukáš Kubala. 2019. "Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response" Toxins 11, no. 4: 218. https://doi.org/10.3390/toxins11040218
APA StyleMoosová, Z., Šindlerová, L., Ambrůzová, B., Ambrožová, G., Vašíček, O., Velki, M., Babica, P., & Kubala, L. (2019). Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response. Toxins, 11(4), 218. https://doi.org/10.3390/toxins11040218