Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins



Abstract
1. Introduction
2. ABC Transporters are One of the Primary Receptors for Several 3D-Cry Toxins
2.1. ABC Transporters Facilitate Induction of Cell Swelling by 3D-Cry Toxins
2.2. BmABCC2 Facilitates Cation-Permeable Pore Formation by Cry1A Toxins
2.3. Binding to 3D-Cry Toxins
3. Role of ABC Transporters as Receptors for 3D-Cry Toxins
4. Receptor Functions of ABCC2, Cadherin-Like Receptor, and Aminopeptidase N
5. Relationship between Binding Affinity to 3D-Cry Toxins and Receptor Activity
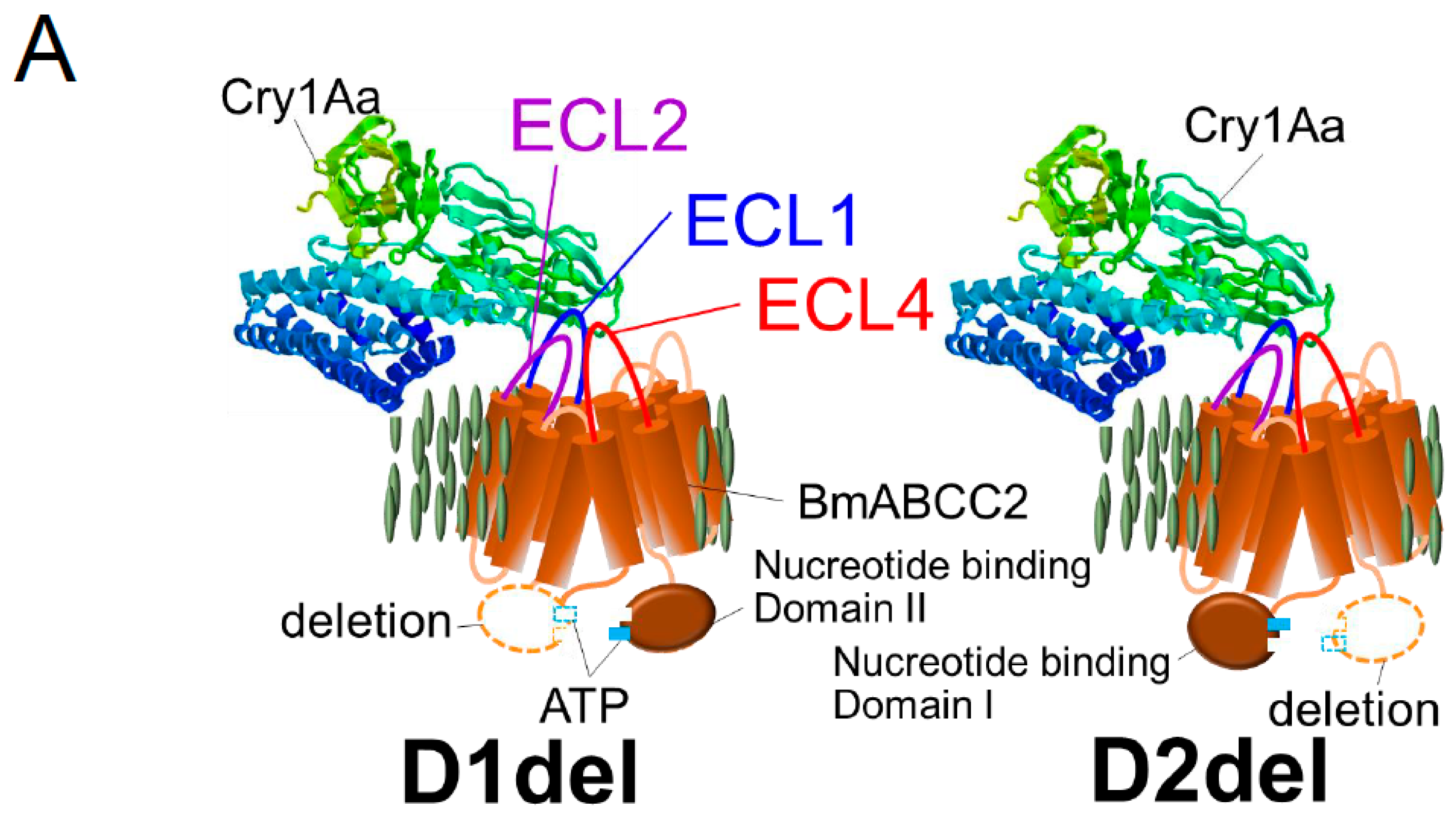
6. Structures of BmABCC2 Responsible for Its Function as a Cry1Aa Toxin Receptor
7. ABCC2-Binding Sites in Cry1Aa
8. Synergism of ABCCs and Cadherin-Like Receptor and Roles in Susceptibility Determination
9. Production of Multiple Toxins by B. Thuringiensis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABC transporters | ATP-binding cassette transporters |
| 3D-Cry toxins | 3-domain Cry toxins |
| ABCC2 | ABC transporter family C2 |
| ABCB1 | ABC transporter family B1 |
| ABCA2 | ABC transporter family A2 |
| ECL | extracellular loop |
| BtR175-TBR | BtR175 toxin-binding region |
| SPR | surface plasmon resonance |
References
- Fred, G.; Arne, A.; Alan, R.; Lara, B.; William, M. Selection and genetic analysis of a Heliothis virescens (Lepidoptera: Noctuidae) strain with high levels of resistance to Bacillus thuringiensis toxins. J. Econ. Entomol. 1995, 88, 1545–1559. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Liu, Y.-B.; Finson, N.; Masson, L.; Heckel, D.G. One gene in diamondback moth confers resistance to four Bacillus thuringiensis toxins. Proc. Natl. Acad. Sci. USA 1997, 94, 1640–1644. [Google Scholar] [CrossRef] [PubMed]
- Gahan, L.J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS Genet. 2010, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, N.; Badenes-Pérez, F.R.; Baxter, S.W.; Heckel, D.G.; Jiggins, C.D.; Kain, W.; Morrison, A.; Vogel, H.; Wang, P.; Badenes-Pérez, F.R.; et al. Parallel evolution of Bt toxin resistance in lepidoptera. Genetics 2011, 189, 1–19. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, T.T.; Liu, C.; Heckel, D.G.; Li, X.; Tabashnik, B.E.; Wu, K. Mis-Splicing of the ABCC2 gene linked with Bt toxin resistance in Helicoverpa armigera. Sci. Rep. 2014, 4, 6184. [Google Scholar] [CrossRef]
- Atsumi, S.; Miyamoto, K.; Yamamoto, K.; Narukawa, J.; Kawai, S.; Sezutsu, H.; Kobayashi, I.; Uchino, K.; Tamura, T.; Mita, K.; et al. Single amino acid mutation in an ATP-binding cassette transporter gene causes resistance to Bt toxin Cry1Ab in the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2012, 109, E1591–E1598. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Adang, M.J. Characterization of a Cry1Ac-receptor alkaline phosphatase in susceptible and resistant Heliothis virescens Larvae. Eur. J. Biochem. 2004, 271, 3127–3135. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, J.-Z.; Rodrigo-simo, A.; Kain, W.; Janmaat, A.F.; Shelton, A.M.; Ferre, J.; Myers, J. Mechanism of resistance to Bacillus thuringiensis toxin Cry1Ac in a greenhouse population of the cabbage looper, Trichoplusia ni. Appl. Environ. Microbiol. 2007, 73, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, Y.; Koike, T.; Sasaki, K.; Yoshimoto, A.; Furukawa, Y. The cadherin-like protein is essential to specificity determination and cytotoxic action of the Bacillus thuringiensis insecticidal CryIAa toxin. FEBS Lett. 1999, 460, 385–390. [Google Scholar] [CrossRef]
- Gahan, L.J.; Gould, F.; Heckel, D.G. Identification of a gene associated with Bt resistance in Heliothis Virescens. Science 2001, 293, 857–860. [Google Scholar] [CrossRef]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac δ-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. Microbiol. 2005, 71, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carrière, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Finson, N.; Groeters, F.R.; Moart, W.J.; Johnson, M.W.; Luo, K.; Adang, M.J. Reversal of resistance to Bacillus thuringiensis in Plutella xylostella. Proc. Natl. Acad. Sci. USA 1994, 91, 4120–4124. [Google Scholar] [CrossRef] [PubMed]
- Mahon, A.R.J.; Olsen, K.M.; Garsia, K.A.; Young, S.R. Resistance to Bacillus thuringiensis toxin Cry2Ab in a strain of Helicoverpa armigera (Lepidoptera: Noctuidae) in Australia. J. Econ. Entomol. 2007, 100, 894–902. [Google Scholar] [CrossRef]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Wslsh, T.K.; Downes, S.; James, W.J.; Lee, S.-F.; Reineke, A.; Williams, A.K.; et al. Insect resistance to Bacillus thuringiensis toxin Cry2Ab is conferred by mutations in an ABC transporter subfamily A protein. PLoS Genet. 2015, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Liu, S.; Liu, L.; Tay, T.W.; Walsh, T.K.; Yang, Y.; Wu, Y. CRISPR/Cas9 mediated genome editing of helicoverpa armigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Pauchet, Y.; Bretschneider, A.; Augustin, S.; Heckel, D.G. A P-glycoprotein is linked to resistance to the Bacillus thuringiensis Cry3Aa toxin in a leaf beetle. Toxins 2016, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of P-glycoprotein reveals a molecular basis for poly-specific drug binding. Science 2009, 323, 1718–1722. [Google Scholar] [CrossRef] [PubMed]
- Gazit, E.; La Rocca, P.; Sansom, M.S.; Shai, Y. The structure and organization within the membrane of the helices composing the pore-forming domain of Bacillus thuringiensis delta-endotoxin are consistent with an “umbrella-like” structure of the pore. Proc. Natl. Acad. Sci. USA 1998, 95, 12289–12294. [Google Scholar] [CrossRef]
- Schwartz, J.-L.; Lu, Y.-J.; Söhnlein, P.; Brousseau, R.; Laprade, R.; Masson, L.; Adang, M.J. Ion channels formed in planar lipid bilayers by Bacillus thuringiensis toxins in the presence of Manduca sexta midgut receptors. FEBS Lett. 1997, 412, 270–276. [Google Scholar] [CrossRef]
- Sangadala, S.; Walters, F.S.; English, L.H.; Adangt, M.J. A mixture of Manduca sexta aminopeptidase and phosphatase enhances Bacillus thuringiensis insecticidal CrylA(c) toxin binding and 86Rb(+)-K+ efflux in vitro. J. Biol. Chem. 1994, 269, 10088–10092. [Google Scholar]
- Luo, K.; Sangadala, S.; Masson, L.; Mazza, A.; Brousseau, R.; Adang, M.J. The Heliothis virescens 170 KDa aminopeptidase functions as “receptor A” by mediating specific Bacillus thuringiensis Cry1A δ-endotoxin binding and pore formation. Insect Biochem. Mol. Biol. 1997, 27, 735–743. [Google Scholar] [CrossRef]
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Physiol. 2012, 104, 103–110. [Google Scholar] [CrossRef]
- Tanaka, S.; Endo, H.; Adegawa, S.; Iizuka, A.; Imamura, K.; Kikuta, S.; Sato, R. Bombyx mori ABC transporter C2 structures responsible for the receptor function of Bacillus thuringiensis Cry1Aa toxin. Insect Biochem. Mol. Biol. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Nishiitsutsuji-Uwo, J. Mode of action of Bacillus thuringiensis δ-endotoxin: Histopathological changes in the silkworm midgut. J. Insect Physiol. 1980, 36, 90–103. [Google Scholar] [CrossRef]
- Tanaka, S.; Yoshizawa, Y.; Sato, R. Response of midgut epithelial cells to Cry1Aa is toxin-dependent and depends on the interplay between toxic action and the host apoptotic response. FEBS J. 2012, 279, 1071–1079. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Rose-Young, L.; Bulla, L.A., Jr. Cytotoxicity of Bacillus thuringiensis Cry1Ab toxin depends on specific binding of the toxin to the cadherin receptor BT-R1 expressed in insect cells. Cell Death Differ. 2005, 12, 1407–1416. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef]
- Tanaka, S.; Miyamoto, K.; Noda, H.; Jurat-Fuentes, J.L.; Yoshizawa, Y.; Endo, H.; Sato, R. The ATP-binding cassette transporter subfamily C member 2 in Bombyx mori larvae is a functional receptor for cry toxins from Bacillus thuringiensis. FEBS J. 2013, 280, 1782–1794. [Google Scholar] [CrossRef]
- Endo, H.; Tanaka, S.; Imamura, K.; Adegawa, S.; Kikuta, S.; Sato, R. Cry toxin specificities of insect ABCC transporters closely related to lepidopteran ABCC2 transporters. Peptides 2017, 98, 1–7. [Google Scholar] [CrossRef]
- Obata, F.; Tanaka, S.; Kashio, S.; Tsujimura, H.; Sato, R.; Miura, M. Induction of Rapid and selective cell necrosis in Drosophila using Bacillus thuringiensis Cry toxin and its silkworm receptor. BMC Biol. 2015, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Bretschneider, A.; Heckel, D.G.; Vogel, H. Know your ABCs: Characterization and gene expression dynamics of ABC transporters in the polyphagous herbivore Helicoverpa armigera. Insect Biochem. Mol. Biol. 2016, 72, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.; Song, S.; Bruning, J.B.; Choo, A.; Baxter, S.W. Expressing a moth Abcc2 Gene in transgenic Drosophila causes susceptibility to Bt Cry1Ac without requiring a cadherin-like protein receptor. Insect Biochem. Mol. Biol. 2017, 80, 61–70. [Google Scholar] [CrossRef]
- Carroll, J.; Ellar, D.J. Analysis of the large aqueous pores produced by a Bacillus thuringiensis protein insecticide in manduca sexta midgut-brush-border-membrane vesicles. Eur. J. Biochem. 1997, 245, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Liebig, B.; Stetson, D.L.; Dean, D.H. Quantification of the effect of Bacillus thuringiensis toxins on short-circuit current in the midgut of Bombyx mori. J. Insect Physiol. 1995, 41, 17–22. [Google Scholar] [CrossRef]
- Hubbard, T.J.P.; Murzin, A.G.; Brenner, S.E.; Chothia, C. SCOP: A structural classification of proteins database. Nucleic Acids Res. 1997, 25, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Ridleya, H.; Johnson, C.L.; Lakey, J.H. Interfacial interactions of pore-forming colicins. In Proteins Membrane Binding and Pore Formation; Anderluh, G., Lakey, J., Eds.; Springer: New York, NY, USA, 2010; pp. 81–90. [Google Scholar] [CrossRef]
- London, E. The mechanism of diphtheria toxin translocation across membranes. In Membrane Protein Transport; Rothman, S.S., Ed.; Elsevier BV: Amsterdam, The Netherlands, 1995; Volume 1, pp. 201–227. [Google Scholar] [CrossRef]
- Endo, H.; Azuma, M.; Adegawa, S.; Kikuta, S.; Sato, R. Water influx via aquaporin directly determines necrotic cell death induced by the Bacillus Thuringiensis Cry toxin. FEBS Lett. 2016, 591, 56–64. [Google Scholar] [CrossRef]
- Knowles, B.H.; Ellar, D.J. Colloid-osmotic liysis is a general feature of the mechanism of action of Bacillus thuringiensis δ-endotoxins with different insect specificity. BBA-Gen. Subj. 1987, 924, 509–518. [Google Scholar] [CrossRef]
- Tanaka, S.; Endo, H.; Adegawa, S.; Kikuta, S.; Sato, R. Functional characterization of Bacillus thuringiensis Cry toxin receptors explains resistance in insects. FEBS J. 2016, 1–17. [Google Scholar] [CrossRef]
- Vadlamudi, R.K.; Ji, T.H.; Bulla, L.A. A specific binding protein from Manduca sexta for the insecticidal toxin of Bacillus thuringiensis Subsp. Berliner. J. Biol. Chem. 1993, 268, 12334–12340. [Google Scholar]
- Yaoi, K.; Nakanishi, K.; Kadotani, T.; Imamura, M.; Koizumi, N.; Iwahana, H.; Sato, R. Bacillus thuringiensis Cry1Aa toxin-binding region of Bombyx mori aminopeptidase N. FEBS Lett. 1999, 463, 221–224. [Google Scholar] [CrossRef]
- Mcnall, R.J.; Adang, M.J. Identification of novel Bacillus thuringiensis Cry1Ac binding proteins in Manduca sexta midgut through proteomic analysis. Insect Biochem. Mol. Biol. 2003, 33, 999–1010. [Google Scholar] [CrossRef]
- Hossain, D.M.; Shitomi, Y.; Moriyama, K.; Higuchi, M.; Hayakawa, T.; Mitsui, T.; Sato, R.; Hori, H. Characterization of a novel plasma membrane protein, expressed in the midgut epithelia of Bombyx mori, that binds to Cry1A toxins. Appl. Environ. Microbiol. 2004, 70, 4604–4612. [Google Scholar] [CrossRef]
- Pandian, G.N.; Ishikawa, T.; Vaijayanthi, T.; Hossain, D.M. Formation of macromolecule complex with Bacillus thuringiensis Cry1A Toxins and chlorophyllide binding 252-KDa lipocalin-like protein locating on Bombyx mori midgut membrane. J. Membr. Biol. 2010, 125–136. [Google Scholar] [CrossRef]
- Valaitis, A.P.; Jenkins, J.L.; Lee, M.K.; Dean, D.H.; Garner, K.J. Isolation and partial characterization of gypsy moth BTR-270, an anionic brush border membrane glycoconjugate that binds Bacillus thuringiensis Cry1A toxins with high affinity. Collab. Entomol. Soc. Am. 2001, 46, 2001. [Google Scholar] [CrossRef]
- Galián, C.; Manon, F.; Dezi, M.; Torres, C.; Ebel, C.; Lévy, D.; Jault, J.-M. Optimized purification of a heterodimeric ABC transporter in a highly stable form amenable to 2-D crystallization. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Adegawa, S.; Nakama, Y.; Endo, H.; Shinkawa, N.; Kikuta, S.; Sato, R. The domain II loops of Bacillus thuringiensis Cry1Aa form an overlapping interaction site for two Bombyx mori larvae functional receptors, ABC transporter C2 and cadherin-like receptor. BBA Proteins Proteom. 2017, 1865, 220–231. [Google Scholar] [CrossRef]
- Park, Y.; González-Martínez, R.M.; Navarro-Cerrillo, G.; Chakroun, M.; Kim, Y.; Ziarsolo, P.; Blanca, J.; Cañizares, J.; Ferré, J.; Herrero, S. ABCC transporters mediate insect resistance to multiple Bt toxins revealed by bulk segregant analysis. BMC Biol. 2014, 12, 46. [Google Scholar] [CrossRef]
- Guo, Z.; Kang, S.; Zhu, X.; Xia, J.; Wu, Q.; Wang, S.; Xie, W.; Zhang, Y. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management. Sci. Rep. 2015, 5, 13728. [Google Scholar] [CrossRef]
- Chen, Z.; He, F.; Xiao, Y.; Liu, C.; Li, J.; Yang, Y.; Ai, H.; Peng, J.; Hong, H.; Liu, K. Endogenous expression of a Bt toxin receptor in the Cry1Ac-susceptible insect cell line and its synergistic effect with cadherin on cytotoxicity of activated Cry1Ac. Insect Biochem. Mol. Biol. 2015, 59, 1–17. [Google Scholar] [CrossRef]
- Endo, H.; Tanaka, S.; Adegawa, S.; Ichino, F.; Tabunoki, H.; Kikuta, S.; Sato, R. Cro extracellular loop structures in silkworm ABCC transporters determine their specificities for Bacillus thuringiensis Cry. J. Biol. Chem. 2018, 293, 8569–8577. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Cheng, T.; Wang, G.; Duan, J.; Niu, W.; Xia, Q. Genome-wide analysis of the ATP-binding cassette (abc) transporter gene family in the silkworm, Bombyx mori. Mol. Biol. Rep. 2012, 39, 7281–7291. [Google Scholar] [CrossRef] [PubMed]
- Augustin, S.; Courtin, C.; Rejasse, A.; Lorme, P.; Genissel, A.; Bourguet, D. Genetics of resistance to transgenic Bacillus thuringiensis poplars in Chrysomela tremulae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2004, 97, 1058–1064. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role ininsecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef]
- Tsuda, Y.; Nakatani, F.; Hashimoto, K.; Ikawa, S.; Matsuura, C.; Fukada, T.; Sugimoto, K.; Himeno, M. Cytotoxic activity of Bacillus thuringiensis Cry Proteins on mammalian cells transfected with cadherin-like Cry receptor gene of Bombyx mori (silkworm). Biochem. J. 2003, 369, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Atsumi, S.; Yaoi, K.; Nakanishi, K.; Higurashi, S.; Miura, N.; Tabunoki, H.; Sato, R. A cadherin-like protein functions as a receptor for Bacillus thuringiensis Cry1Aa and Cry1Ac toxins on midgut epithelial cells of Bombyx Mori larvae. FEBS Lett. 2003, 538, 29–34. [Google Scholar] [CrossRef]
- Banks, D.J.; Hua, G.; Adang, M.J. Cloning of a Heliothis Virescens 110 KDa aminopeptidase N and expression in drosophila S2 cells. Insect Biochem. Mol. Biol. 2003, 33, 499–508. [Google Scholar] [CrossRef]
- Crava, C.M.; Bel, Y.; Jakubowska, A.K.; Ferré, J.; Escriche, B. Midgut aminopeptidase N isoforms from Ostrinia nubilalis: Activity characterization and differential binding to Cry1Ab and Cry1Fa proteins from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2013, 43, 924–935. [Google Scholar] [CrossRef]
- Soberón, M.; Portugal, L.; Garcia-Gómez, B.-I.; Sánchez, J.; Onofre, J.; Gómez, I.; Pacheco, S.; Bravo, A. Cell lines as models for the study of Cry toxins from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2018, 93, 66–78. [Google Scholar] [CrossRef]
- Chen, J.; Aimanova, K.G.; Pan, S.; Gill, S.S. Identification and characterization of Aedes aegypti aminopeptidase N as a putative receptor of Bacillus thuringiensis Cry11A toxin. Insect Biochem. Mol. Biol. 2009, 39, 688–696. [Google Scholar] [CrossRef]
- Gill, M.; Ellar, D. Transgenic Drosophila reveals a functional in vivo receptor for the Bacillus thuringiensis toxin Cry1Ac1. Insect Mol. Biol. 2002, 11, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Kitami, M.; Kadotani, T.; Nakanishi, K.; Atsumi, S.; Higurashi, S.; Ishizaka, T.; Watanabe, A.; Sato, R. Bacillus Thuringiensis Cry toxins bound specifically to various proteins via domain III, which had a galactose-binding domain-like fold. Biosci. Biotechnol. Biochem. 2011, 7, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Gómez, I.; Rausell, C.; Sánchez, J.; Soberón, M.; Bravo, A. Structural changes of the Cry1Ac oligomeric pre-pore from Bacillus thuringiensis induced by N-Acetylgalactosamine facilitates toxin membrane insertion †. Biochemistry 2006, 45, 10329–10336. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Adang, M.J. The Heliothis virescens cadherin protein expressed in Drosophila S2 cells functions as a receptor for Bacillus thuringiensis Cry1A but not Cry1Fa toxins. Biochemistry 2006, 45, 9688–9695. [Google Scholar] [CrossRef]
- Ren, X.-L.; Ma, Y.; Cui, J.-J.; Li, G.-Q. RNA interference-mediated knockdown of three putative aminopeptidases N affects susceptibility of Spodoptera exigua larvae to Bacillus thuringiensis Cry1Ca. J. Insect Physiol. 2014, 67, 28–36. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, P.; Zhang, B.; Liu, L.; Wang, X.; Lei, C.; Lin, Y.; Ma, W. Proteomic analysis of Cry1Ac and Cry2Aa binding proteins in brush border membrane vesicles of the Chilo suppressalis midgut. J. Asia. Pac. Entomol. 2017, 20, 1165–1169. [Google Scholar] [CrossRef]
- Peña-Cardeña, A.; Grande, R.; Sánchez, J.; Tabashnik, B.E.; Bravo, A.; Soberón, M.; Gómez, I. The C-Terminal protoxin domain of Bacillus thuringiensis Cry1Ab toxin has a functional role in binding to GPI-anchored receptors in the insect midgut. J. Biol. Chem. 2018, 2016, 1–21. [Google Scholar] [CrossRef]
- Tanaka, S.; Miyamoto, K.; Noda, H.; Endo, H.; Kikuta, S.; Sato, R. Single amino acid insertions in extracellular loop 2 of Bombyx mori ABCC2 disrupt its receptor function for Bacillus thuringiensis Cry1Ab and Cry1Ac but not Cry1Aa toxins. Peptides 2016, 78, 99–108. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Z.; Yang, Y.; Xiao, Y.; Liu, C.; Ma, Y.; Soberón, M.; Bravo, A.; Yang, Y.; Liu, K. A single amino acid polymorphism in ABCC2 loop 1 is responsible for differential toxicity of Bacillus thuringiensis Cry1Ac toxin in different Spodoptera (Noctuidae) species. Insect Biochem. Mol. Biol. 2018, 100, 59–65. [Google Scholar] [CrossRef]
- Gómez, I.; Sánchez, J.; Miranda, R.; Bravo, A.; Soberón, M. Cadherin-like receptor binding facilitates proteolytic cleavage of helix α-1 in domain i and oligomer pre-pore formation of Bacillus thuringiensis Cry1Ab toxin. FEBS Lett. 2002, 513, 242–246. [Google Scholar] [CrossRef]
- Gómez, I.; Dean, D.H.; Bravo, A.; Soberón, M. Molecular basis for Bacillus Thuringiensis Cry1Ab toxin specificity: Two structural determinants in the Manduca sexta Bt-R1 receptor interact with loops α-8 and 2 in domain II of Cy1Ab toxin †. Biochemistry 2003, 42, 10482–10489. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Zhuang, M.; Ross, L.S.; Gomez, I.; Oltean, D.I.; Bravo, A.; Soberon, M.; Gill, S.S. Single amino acid mutations in the cadherin receptor from Heliothis virescens affect its toxin binding ability to Cry1A toxins. J. Biol. Chem. 2005, 280, 8416–8425. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kain, W.; Wang, P. Bacillus thuringiensis Cry1A toxins exert toxicity by multiple pathways in insects. Insect Biochem. Mol. Biol. 2018, 102, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Ocelotl, J.; Sánchez, J.; Gómez, I.; Tabashnik, B.E.; Bravo, A.; Soberón, M. ABCC2 is associated with Bacillus thuringiensis Cry1Ac toxin oligomerization and membrane insertion in diamondback moth. Sci. Rep. 2017, 7, 2386. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Adegawa, S.; Kikuta, S.; Sato, R. The intracellular region of silkworm cadherin-like protein is not necessary to mediate the toxicity of Bacillus thuringiensis Cry1Aa and Cry1Ab toxins. Insect Biochem. Mol. Biol. 2018, 94, 36–41. [Google Scholar] [CrossRef]
- Chen, J.; Hua, G.; Jurat-Fuentes, J.L.; Abdullah, M.A.; Adang, M.J. Synergism of Bacillus thuringiensis toxins by a fragment of a toxin-binding cadherin. Proc. Natl. Acad. Sci. USA 2007, 104, 13901–13906. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Hua, G.; Abdullah, M.A.F.; Rahman, K.; Adang, M.J. Cadherin fragments from Anopheles gambiae synergize Bacillus thuringiensis Cry4Ba’s toxicity against Aedes aegypti larvae. Appl. Environ. Microbiol. 2009, 75, 7280–7282. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.; Park, Y.; Adang, M.J. Cadherin AdCad1 in alphitobius diaperinus larvae is a receptor of Cry3Bb toxin from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2014, 45, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Dorsch, J.A.; Candas, M.; Griko, N.B.; Maaty, W.S.A.; Midboe, E.G.; Vadlamudi, R.K.; Bulla, L.A., Jr. Cry1A toxins of Bacillus thuringiensis bind specifically to a region adjacent to the membrane-proximal extracellular domain of BT-R1in Manduca sexta: Involvement of a cadherin in the-entomopathogenicity of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2002, 32, 1025–1036. [Google Scholar] [CrossRef]
- Bravo, A.; Sarabia, S.; Lopez, L.; Ontiveros, H.; Abarca, C.; Ortiz, A.; Ortiz, M.; Lina, L.; Villalobos, F.J.; Pen, G.; et al. Characterization of Cry genes in a Mexican Bacillus thuringiensis strain collection. Appl. Environ. Microbiol. 1998, 64, 4965–4972. [Google Scholar] [PubMed]
- Martínez, C.; Caballero, P. Contents of cry genes and insecticidal toxicity of Bacillus thuringiensis strains from terrestrial and aquatic habitats. J. Appl. Entomol. 2002, 92, 745–752. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Malvar, T.; Liu, Y.B.; Finson, N.; Borthakur, D.; Shin, B.S.; Park, S.H.; Masson, L.; de Maagd, R.A.; Bosch, D. Cross-resistance of the diamondback moth indicates altered interactions with domain II of Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 1996, 62, 2839–2844. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Z.; Li, Y.-X.; Collins, H.L.; Cao, J.; Earle, E.D.; Shelton, A.M. Different cross-resistance patterns in the diamondback moth (Lepidoptera: Plutellidae) resistant to Bacillus thuringiensis toxin Cry1C. J. Econ. Entomol. 2001, 94, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Paris, M.; Tetreau, G.; Laurent, F.; Lelu, M.; Despres, L.; David, J.-P. Persistence of Bacillus thuringiensis israelensis (Bti) in the environment induces resistance to multiple Bti toxins in mosquitoes. Pest Manag. Sci. 2011, 67, 122–128. [Google Scholar] [CrossRef]
- Paul, A.; Harrington, L.H.; Zhang, L.; Scott, J.G. Insecticide resistance in Culex pipiens from New York. J. Am. Mosq. Control Assoc. 2005, 21, 2005. [Google Scholar]
- Li, H.; Bouwer, G. Evaluation of the synergistic activities of Bacillus thuringiensis Cry proteins against Helicoverpa armigera (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2014, 121, 7–13. [Google Scholar] [CrossRef]
- López-Diaz, J.A.; Cantón, P.E.; Gill, S.S.; Soberón, M.; Bravo, A. Oligomerization is a key step in Cyt1Aa membrane insertion and toxicity but not necessary to synergize Cry11Aa toxicity in Aedes aegypti larvae. Environ. Microbiol. 2013, 15, 3030–3039. [Google Scholar] [CrossRef] [PubMed]
- Lemes, A.N.A.; Figueiredo, C.S.; Sebastião, I.; da Silva, L.M.; da Costa Alves, R.; de Siqueira, H.Á.A.; Franco Lemos, M.V.; Fernandes, O.A.; Desidério, J.A. Cry1Ac and Vip3Aa proteins from Bacillus thuringiensis targeting Cry toxin resistance in Diatraea flavipennella and Elasmopalpus lignosellus from sugarcane. PeerJ 2017, 5, e2866. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor. | Toxin | KD (M) 1 | Effective Conc. (nM) in Cell Swelling Assay 2 |
|---|---|---|---|
| BmABCC2 | Cry1Aa | 3.1 × 10−10 | 0.1 |
| Cry1Ab | 2.6 × 10−10 | 100 | |
| Cry1Ca | 1.7 × 10−7 | >10,000 | |
| Cry1Da | 2.3 × 10−6 | >2000 | |
| Cry3Bb | 2.0 × 10−5 | >500 | |
| Cry8Ca | 1.9 × 10−5 | 1000 | |
| BmABCC2 from resistant line | Cry1Aa | 2.8 × 10−10 | 10 |
| Cry1Ab | 2.4 × 10−8 | >1000 | |
| BmABCC3 | Cry1Aa | 3.4 × 10−8 | 100 |
| Cry1Ab | 6.9 × 10−8 | >4500 | |
| Cry1Ca | 3.9 × 10−7 | >1000 | |
| Cry1Da | 4.2 × 10−4 | >1000 | |
| Cry3Bb | 4.0 × 10−8 | >1000 | |
| TcABCC4A | Cry1Aa | not detectable | >1000 |
| Cry1Ca | not detectable | >2500 | |
| Cry1Da | not detectable | >2000 | |
| Cry3Bb | 1.8 × 10−5 | >1700 | |
| Cry8Ca | 4.0 × 10−8 | 100 | |
| BtR175(BmCad) | Cry1Aa | 7.2 × 10−10 | 200 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, R.; Adegawa, S.; Li, X.; Tanaka, S.; Endo, H. Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins. Toxins 2019, 11, 124. https://doi.org/10.3390/toxins11020124
Sato R, Adegawa S, Li X, Tanaka S, Endo H. Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins. Toxins. 2019; 11(2):124. https://doi.org/10.3390/toxins11020124
Chicago/Turabian StyleSato, Ryoichi, Satomi Adegawa, Xiaoyi Li, Shiho Tanaka, and Haruka Endo. 2019. "Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins" Toxins 11, no. 2: 124. https://doi.org/10.3390/toxins11020124
APA StyleSato, R., Adegawa, S., Li, X., Tanaka, S., & Endo, H. (2019). Function and Role of ATP-Binding Cassette Transporters as Receptors for 3D-Cry Toxins. Toxins, 11(2), 124. https://doi.org/10.3390/toxins11020124