Occurrence and Ear Damage of Helicoverpa zea on Transgenic Bacillus thuringiensis Maize in the Field in Texas, U.S. and Its Susceptibility to Vip3A Protein

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Plant Injury and Occurrence of H. zea on Different Hybrids of Non-Bt and Bt Maize in the Field
2.2. Susceptibility of Two Different Field Populations of H. zea to Vip3A Protein in Diet Bioassays
2.3. Larval Survival and Development of H. zea Populations on Cotton Leaf Tissues
3. Discussion
4. Materials and Methods
4.1. Source of Bt and Non-Bt Maize Hybrids
4.2. Field Planting
4.3. Dose-Response Bioassays
4.4. Cotton Leaf Tissue Bioassays
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- James, C. Global Status of Commercialized Biotech/GM Crops: 2017; Brief No. 53; ISAAA: Ithaca, NY, USA, 2018; Available online: https://www.isaaa.org/resources/publications/briefs/53/download/isaaa-brief-53-2017.pdf (accessed on 8 February 2019).
- Carpenter, J.E. Peer-reviewed surveys indicate positive impact of commercialized GM crops. Nat. Biotechnol. 2010, 28, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Edgerton, M.D.; Fridgen, J.; Anderson, J.R., Jr.; Ahlgrim, J.; Criswell, M.; Dhungana, P.; Gocken, T.; Li, Z.; Mariappan, S.; Pilcher, C.D.; et al. Transgenic insect resistance traits increase corn yield and yield stability. Nat. Biotechnol. 2012, 30, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Guo, Y.Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Kathage, J.; Qaim, M. Economic impacts and impact dynamics of Bt (Bacillus thuringiensis) cotton in India. Proc. Natl. Acad. Sci. USA 2012, 109, 11652–11656. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]
- Grimi, D.A.; Parody, B.; Ramos, M.L.; Machado, M.; Ocampo, F.; Willse, A.; Martinelli, S.; Head, G. Field-evolved resistance to Bt maize in sugarcane borer (Diatraea saccharalis) in Argentina. Pest Manag. Sci. 2018, 74, 905–913. [Google Scholar] [CrossRef]
- Smith, J.L.; Lepping, M.D.; Rule, D.M.; Farhan, Y.; Schaafsma, A.W. Evidence for field-evolved resistance of Striacosta albicosta (Lepidoptera: Noctuidae) to Cry1F Bacillus thuringiensis protein and transgenic corn hybrids in Ontario, Canada. J. Econ. Entomol. 2017, 110, 2217–2228. [Google Scholar] [CrossRef]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis-derived Cry1F δ-endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef]
- Matten, S.R.; Frederick, R.J.; Reynolds, A.H. United States Environmental Protection Agency insect resistance management programs for plant-incorporated protectants and use of simulation modeling. In Regulation of Agricultural Biotechnology: The United States and Canada; McHughen, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 175–267. [Google Scholar]
- Zhao, J.Z.; Cao, J.; Li, Y.; Collins, H.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat. Biotechnol. 2003, 21, 1493–1497. [Google Scholar] [CrossRef] [PubMed]
- Difonzo, C.; Porter, P. Handy Bt Trait Table. 2018. Available online: https://lubbock.tamu.edu/files/2018/11/BtTraitTableNov2018.pdf (accessed on 18 January 2019).
- Capinera, J.L. Corn earworm, Helicoverpa (=Heliothis) zea (Boddie) (Lepidoptera: Noctuidae). Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida. 2000, EENY-145 (IN302). Available online: https://www.maine.gov/dacf/php/gotpests/bugs/factsheets/corn-earworm-fla.pdf (accessed on 8 February 2019).
- Siegfried, B.D.; Spencer, T.; Nearman, J. Baseline susceptibility of the corn earworm (Lepidoptera: Noctuidae) to the Cry1Ab toxin from Bacillus thuringiensis. J. Econ. Entomol. 2000, 93, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Burd, A.D.; Bradley, J.R.; Van-Duyn, J.W.; Gould, F. Resistance of bollworm, Helicoverpa zea, to Cry1Ac toxin. In Proceedings of the 2000 Beltwide Cotton Conferences, Memphis, TN, Egypt, 5–8 January 2000. [Google Scholar]
- Sivasupramaniam, S.; Moar, W.J.; Ruschke, L.G.; Osborn, J.A.; Jiang, C.; Sebaugh, J.L.; Brown, G.R.; Shappley, Z.W.; Oppenhuizen, M.E.; Mullins, J.W.; et al. Toxicity and characterization of cotton expressing Bacillus thuringiensis Cry1Ac and Cry2Ab2 proteins for control of lepidopteran pests. J. Econ. Entomol. 2008, 101, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Heuberger, S.; Zhang, M.; Ellers-Kirk, C.; Ni, X.; Masson, L.; Li, X.; Tabashnik, B.E.; Carrière, Y. Potential shortfall of pyramided transgenic cotton for insect resistance management. Proc. Natl. Acad. Sci. USA 2013, 110, 5806–5811. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Venugopal, P.D.; Finkenbinder, C. Field-evolved resistance in corn earworm to Cry proteins expressed by transgenic sweet corn. PLoS ONE 2016, 11, e0169115. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Gore, J.; Catchot, A.; Cook, D.; Musser, F.; Lorenz, G.; Seiter, N.; Stewart, S.; Sebe, B. Continuous monitoring of the susceptibility of Helicoverpa zea in the southern U.S. to different Bt technologies. In Proceedings of the 2018 Beltwide Cotton Conferences, San Antonio, TX, USA, 3–5 January 2018. [Google Scholar]
- Yang, F.; Kerns, D.L.; Gore, J.; Catchot, A.; Lorenz, G.; Stewart, S. Susceptibility of field populations of the cotton bollworm in the southern U.S. to four individual Bt proteins. In Proceedings of the 2017 Beltwide Cotton Conferences, Dallas, TX, USA, 4–6 January 2017. [Google Scholar]
- Yang, F.; Kerns, D.L.; Gore, J.; Cook, D.; Seiter, N. Susceptibility of field populations of the cotton bollworm to purified Cry1Ac protein and cotton leaf tissues containing Cry1Ac protein. In Proceedings of the 2016 Beltwide Cotton Conference, New Orleans, LA, USA, 5–7 January 2016. [Google Scholar]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Aghaee, M.A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long-Term Empirical and Observational Evidence of Practical Helicoverpa zea Resistance to Cotton with Pyramided Bt Toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Mahon, R.J.; Downes, S.J.; James, B. Vip3A resistance alleles exist at high levels in Australian targets before release of cotton expressing this toxin. PLoS ONE 2012, 7, e39192. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wu, S.; Yang, Y.; Wu, Y. Baseline Susceptibility of field populations of Helicoverpa armigera to Bacillus thuringiensis Vip3Aa toxin and lack of cross-resistance between Vip3Aa and Cry toxins. Toxins 2017, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.L.; Unnithan, G.C.; Degain, B.A.; Wei, J.; Zhang, J.; Li, X.; Tabashnik, B.E.; Carrière, Y. Cross-resistance to toxins used in pyramided Bt crops and resistance to Bt sprays in Helicoverpa zea. J. Invert. Pathol. 2015, 132, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Head, G.P.; Price, P.; Huang, F. Cross-resistance to purified Bt proteins, Bt corn and Bt cotton in a Cry2Ab2-corn resistant strain of Spodoptera frugiperda. Pest Manag. Sci. 2017, 73, 2495–2503. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Head, G.P.; Brown, S.; Huang, F. Susceptibility of Cry1F-maize resistant, heterozygous, and susceptible Spodoptera frugiperda to Bt proteins used in the transgenic cotton. Crop Prot. 2017, 98, 128–135. [Google Scholar] [CrossRef]
- Yang, F.; Morsello, S.; Head, G.P.; Sansone, C.; Huang, F.; Gilreath, R.T.; Kerns, D.L. F2 screen, inheritance and cross-resistance of field-derived Vip3A resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae) collected from Louisiana, USA. Pest Manag. Sci. 2018, 74, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defining terms for proactive management of resistance to Bt crops and pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Tabashnik, B.E.; Carrière, Y. A seed mixture increases dominance of resistance to Bt cotton in Helicoverpa zea. Sci. Rep. 2015, 5, 9807. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Brown, S.; Head, G.P.; Huang, F. Pollen contamination in seed mixture increases the dominance of resistance to Bt maize in Spodoptera frugiperda (Lepidoptera: Noctuidae). Pest Manag. Sci. 2017, 73, 2379–2385. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Head, G.P.; Leonard, B.R.; Levy, R.; Niu, Y.; Huang, F. A challenge for the seed mixture refuge strategy in Bt maize: Impact of cross-pollination on an ear-feeding pest, corn earworm. PLoS ONE 2014, 9, e112962. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, D.F. The corn earworm complex. Mem. Entomol. Soc. Can. 1965, 40, 1–246. [Google Scholar] [CrossRef]
- Morey, A.C.; Hutchison, W.D.; Venette, R.C.; Burkness, E.C. Cold hardiness of corn earworm (Lepidoptera: Noctuidae) pupae. Environ. Entomol. 2012, 41, 172–179. [Google Scholar] [CrossRef]
- Sandstrom, M.A.; Changnon, D.; Flood, B.R. Improving Our Understanding of Helicoverpa zea Migration in the Midwest: Assessment of Source Populations. Plant Health Prog, 2007. Available online: http://www.plantmanagementnetwork.org/pub/php/symposium/hzea/migrate/ (accessed on 18 January 2019).
- US Environmental Protection Agency. Bacillus thuringiensis Vip3Aa20 Insecticidal Protein and the Genetic Material Necessary for Its Production (via Elements of Vector pNOV1300) in Event MIR162 Maize. 2009. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/decision_PC-006599_3-Apr-09.pdf (accessed on 18 January 2019).
- US Department of Agriculture, Agricultural Marketing Service. Cotton Varieties Planted 2014 Crop. Available online: https://search.ams.usda.gov/mndms/2014/09/CN20140912AVAR.PDF (accessed on 4 February 2019).
- Yang, F.; Kerns, D.L.; Leonard, B.R.; Oyediran, I.; Burd, T.; Niu, Y.; Huang, F. Performance of Agrisure® Viptera™ 3111 corn against Helicoverpa zea (Lepidoptera: Noctuidae) in seed mixed plantings. Crop Protect. 2015, 69, 77–82. [Google Scholar] [CrossRef]
- Reisig, D.D.; Kurtz, R. Bt resistance implications for Helicoverpa zea (Lepidoptera: Noctuidae) insecticide resistance management in the United States. Environ. Entomol. 2018, 47, 1357–1364. [Google Scholar] [CrossRef]
- Burkness, E.C.; Dively, G.; Patton, T.; Morey, A.C.; Hutchison, W.D. Novel Vip3A Bacillus thuringiensis (Bt) maize approaches high-dose efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under field conditions: Implications for resistance management. GM Crops 2010, 1, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Little, N.S.; Catchot, A.L.; Allen, K.C.; Gore, J.; Musser, F.R.; Cook, D.R.; Luttrell, R.G. Supplemental control with diamides for Heliothines in Bt cotton. Southwest Entomol. 2017, 42, 15–26. [Google Scholar] [CrossRef]
- Reay-Jones, F.P.F.; Bessin, R.T.; Brewer, M.J.; Buntin, D.G.; Catchot, A.L.; Cook, D.R.; Flanders, K.L.; Kerns, D.L.; Porter, R.P.; Reisig, D.D.; Stewart, S.D. Impact of lepidoptera (crambidae, noctuidae, and pyralidae) pests on corn containing pyramided bt traits and a blended refuge in the Southern United States. J. Econ. Entomol. 2016, 109, 1859–1871. [Google Scholar] [CrossRef] [PubMed]
- Bilbo, T.R.; Reay-Jones, F.P.; Reisig, D.D.; Musser, F.R.; Greene, J.K. Effects of Bt corn on the development and fecundity of corn earworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 2018, 111, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Buntin, G.D. Corn expressing Cry1Ab or Cry1F endotoxin for fall armyworm and corn earworm (Lepidoptera: Noctuidae) management in field corn for grain production. Fla. Entomol. 2008, 91, 523–530. [Google Scholar]
- Anilkumar, K.J.; Adams, N.; Clark, T.L.; Dennehy, T.J.; Head, G.P.; Price, P.A. Diet overlay bioassays with Cry2Ab reduce experiment-wise variation and enhance resistance discrimination in Helicoverpa zea, relative to diet-incorporation bioassays: Conclusions from monitoring of resistance in the US in 2008. In Proceedings of the Beltwide Cotton Conferences, National Cotton Council, San Antonio, TX, USA, 5–8 January 2009. [Google Scholar]
- SAS Institute. SAS/STAT, 3rd ed.; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
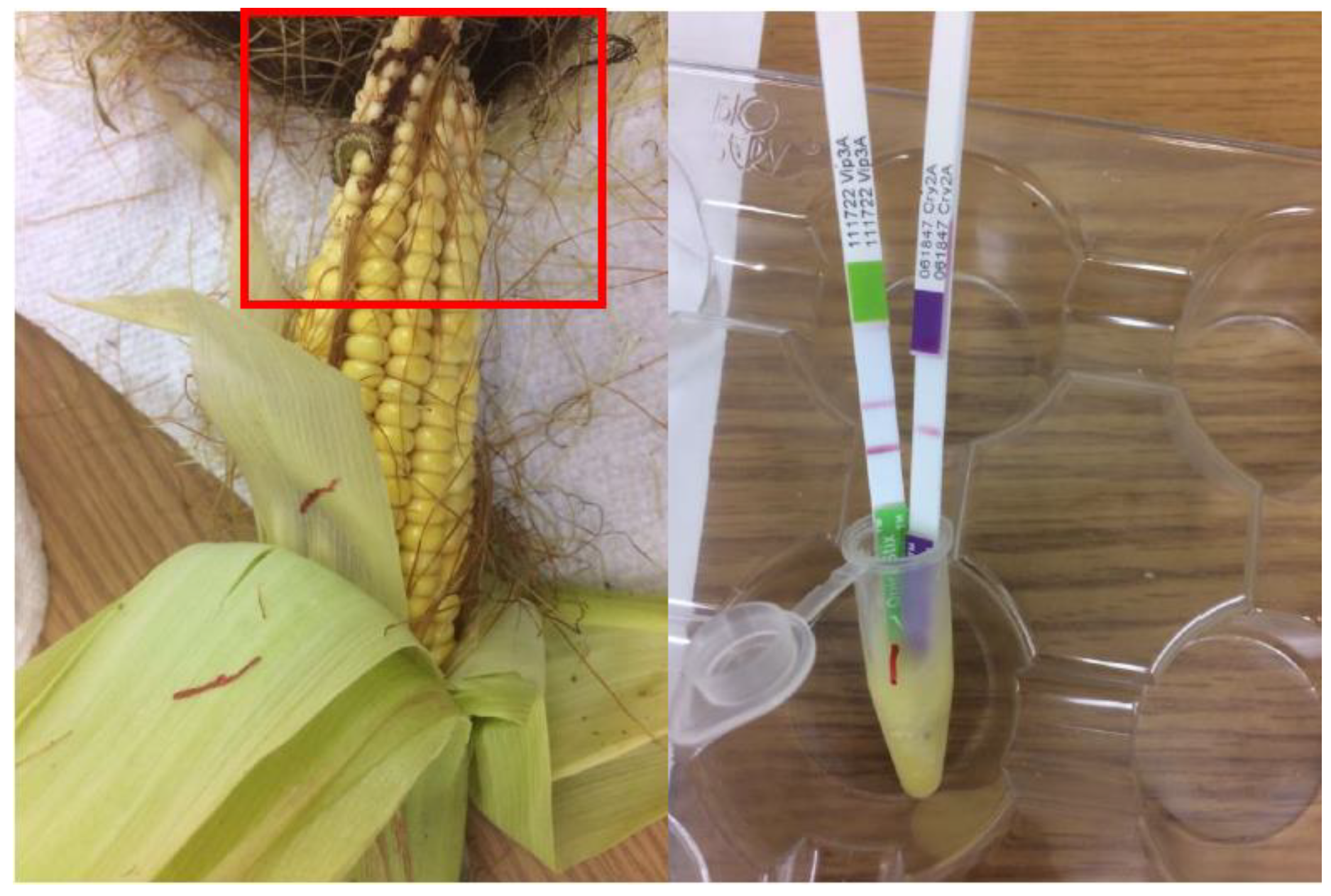


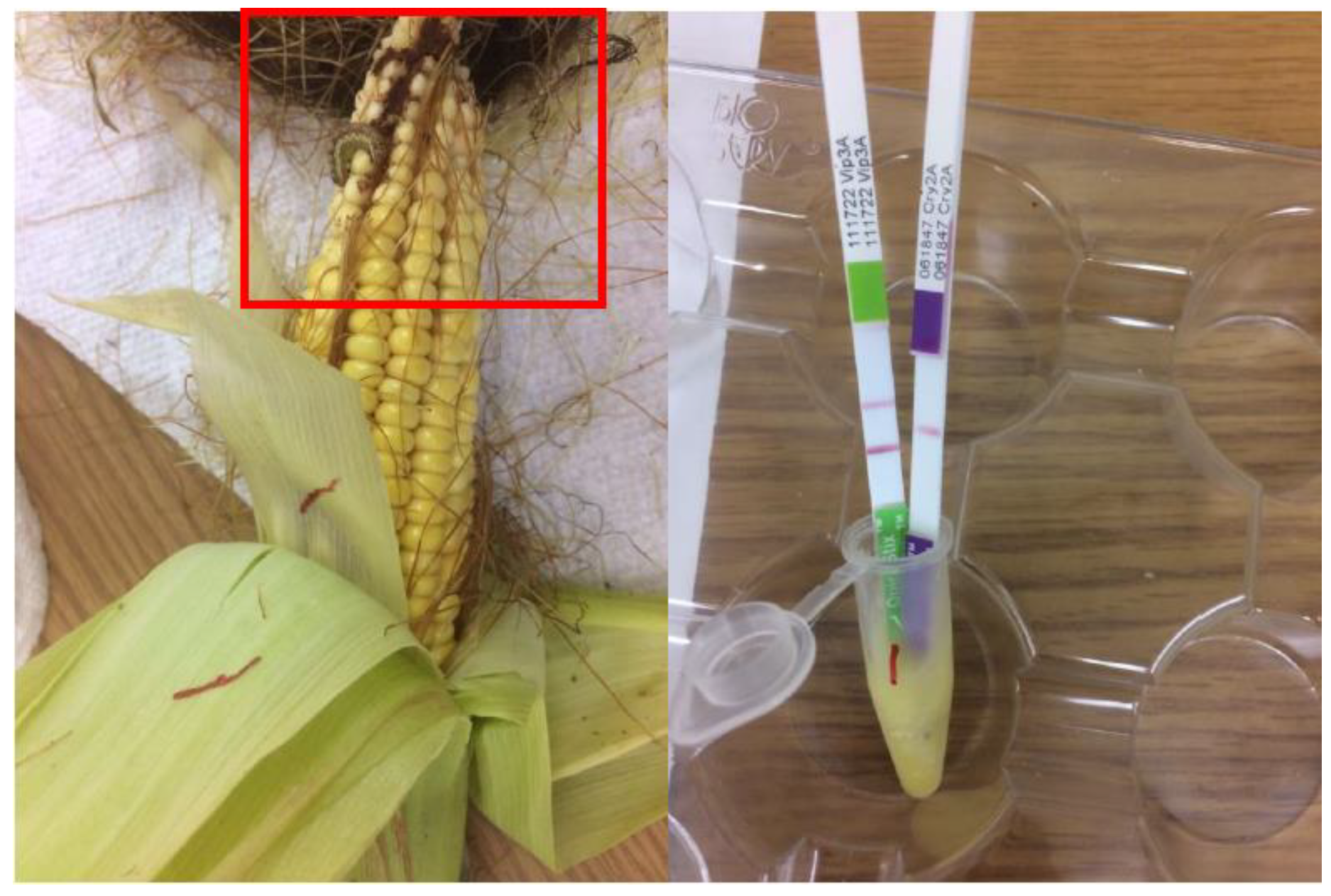{kind=link}
{kind=link}
{kind=link}
| Variety | Bt Proteins | % Plant with Larvae # | Number of Larvae/Ear | Average Instar | % Plant with Damaged Ear § | Damaged Area per Ear (cm2) |
|---|---|---|---|---|---|---|
| DKC-NBt | / | 77.5 ± 6.0 ab | 0.96 ± 0.11 ab | 4.86 ± 0.13 bc | 100.0 ± 0.0 b | 18.1 ± 0.9 b |
| P-NBt | / | 75.0 ± 2.0 ab | 0.93 ± 0.05 ab | 4.94 ± 0.11 c | 100.0 ± 0.0 b | 19.1 ± 1.7 b |
| Intrasect | Cry1Ab+Cry1F | 91.3 ± 4.3 b | 1.85 ± 0.13 b | 4.31 ± 0.06 b | 100.0 ± 0.0 b | 15.7 ± 1.5 b |
| VT2P | Cry1A.105+Cry2Ab2 | 83.8 ± 3.8 ab | 1.30 ± 0.18 b | 4.71 ± 0.11 bc | 100.0 ± 0.0 b | 15.0 ± 1.2 b |
| Leptra | Cry1Ab+Cry1F+Vip3A | 61.3 ± 3.1 a | 0.79 ± 0.04 a | 3.19 ± 0.19 a | 67.5 ± 1.4 a | 1.3 ± 0.2 a |
| F-test | F-value | F4,12 = 6.16 | F4,12 = 13.30 | F4,12 = 40.36 | F4,12 = 1546.47 | F4,12 = 224.56 |
| P-value | 0.0062 | 0.0002 | <0.0001 | <0.0001 | <0.0001 |
| Insect Population * | N # | LC50 (95% CI) (μg/cm2) § | Slope ± SE | X2 | df | Resistance Ratio £ |
|---|---|---|---|---|---|---|
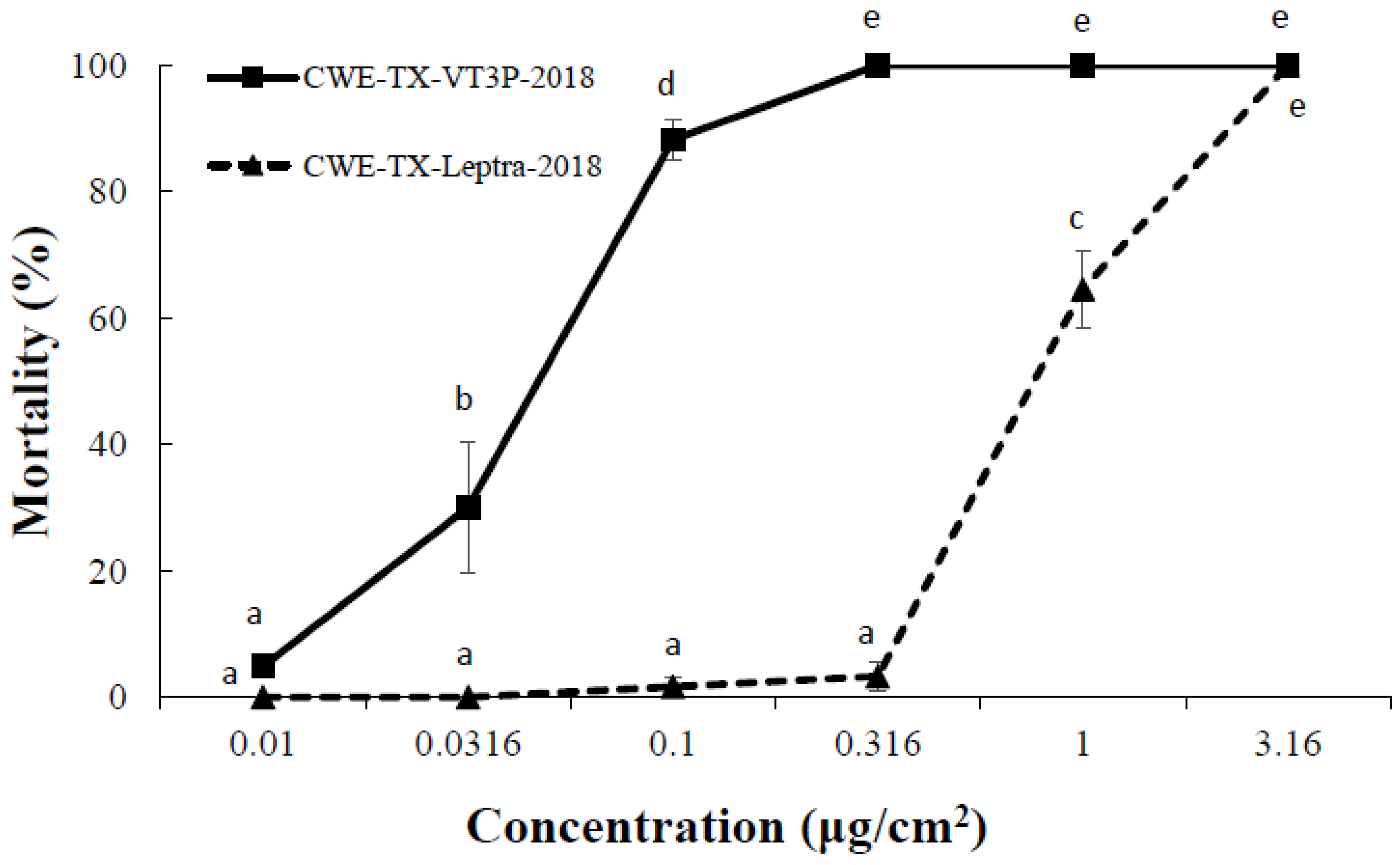
| CEW-TX-VT3P-2018 | 448 | 0.041 (0.035, 0.050) | 2.87 ± 0.30 | 18.9 | 22 | 1.0 |
| CEW-TX-Leptra-2018 | 448 | 0.838 (0.686, 0.966) | 4.93 ± 1.02 | 19.0 | 22 | 20.4 |
| Cotton Variety | Insect § | Survivorship (%) £ | Average Instar | |
|---|---|---|---|---|
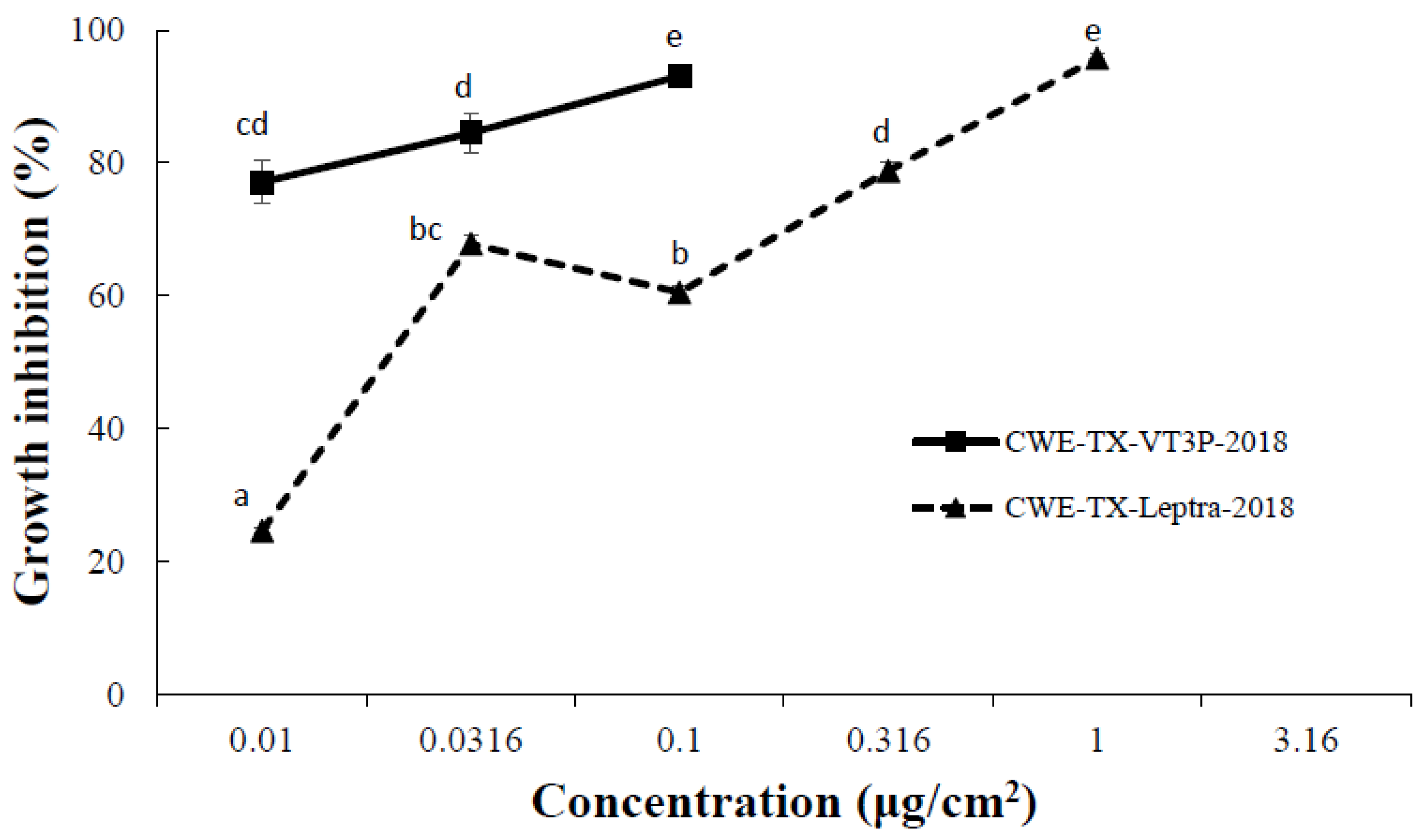
| Non-Bt | CEW-TX-Leptra-2018 | 78.3 ± 2.9 c | 3.39 ± 0.01 c | |
| CEW-TX-SS | 82.5 ± 3.2 c | 3.45 ± 0.05 c | ||
| WideStrike 3 | CEW-TX-Leptra-2018 | 41.7 ± 7.5 b | 2.68 ± 0.03 b | |
| CEW-TX-SS | 3.3 ± 1.4 a | 2.00 ± 0.00 a | ||
| F-test | Insect | F-value | F1, 12 = 19.83 | F1, 11 = 161.46 |
| p-value | 0.0008 | <0.0001 | ||
| Cotton variety | F-value | F1, 12 = 158.41 | F1, 11 = 1483.01 | |
| p-value | <0.0001 | < 0.0001 | ||
| Insect * Cotton variety | F-value | F1, 12 = 29.64 | F1, 11 = 215.14 | |
| p-value | <0.0001 | <0.0001 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; González, J.C.S.; Williams, J.; Cook, D.C.; Gilreath, R.T.; Kerns, D.L. Occurrence and Ear Damage of Helicoverpa zea on Transgenic Bacillus thuringiensis Maize in the Field in Texas, U.S. and Its Susceptibility to Vip3A Protein. Toxins 2019, 11, 102. https://doi.org/10.3390/toxins11020102
Yang F, González JCS, Williams J, Cook DC, Gilreath RT, Kerns DL. Occurrence and Ear Damage of Helicoverpa zea on Transgenic Bacillus thuringiensis Maize in the Field in Texas, U.S. and Its Susceptibility to Vip3A Protein. Toxins. 2019; 11(2):102. https://doi.org/10.3390/toxins11020102
Chicago/Turabian StyleYang, Fei, José C. Santiago González, Jayme Williams, Donald C. Cook, Ryan T. Gilreath, and David L. Kerns. 2019. "Occurrence and Ear Damage of Helicoverpa zea on Transgenic Bacillus thuringiensis Maize in the Field in Texas, U.S. and Its Susceptibility to Vip3A Protein" Toxins 11, no. 2: 102. https://doi.org/10.3390/toxins11020102
APA StyleYang, F., González, J. C. S., Williams, J., Cook, D. C., Gilreath, R. T., & Kerns, D. L. (2019). Occurrence and Ear Damage of Helicoverpa zea on Transgenic Bacillus thuringiensis Maize in the Field in Texas, U.S. and Its Susceptibility to Vip3A Protein. Toxins, 11(2), 102. https://doi.org/10.3390/toxins11020102