RTX Toxins of Animal Pathogens and Their Role as Antigens in Vaccines and Diagnostics

Abstract
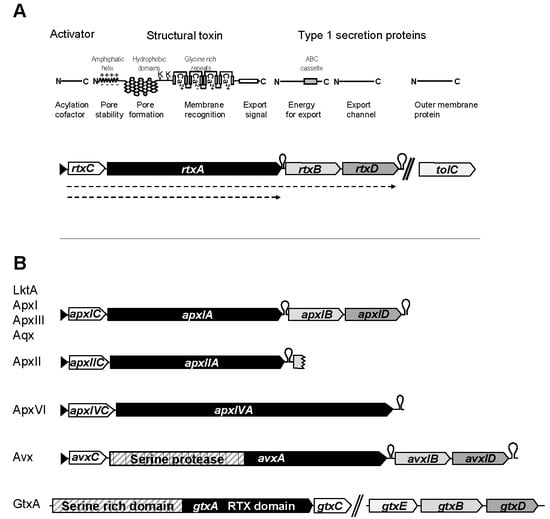
1. Introduction
2. RTX Toxins of Animal Pathogens
2.1. LktA Leukotoxin of Mannheimia (Pasteurella) Haemolytica
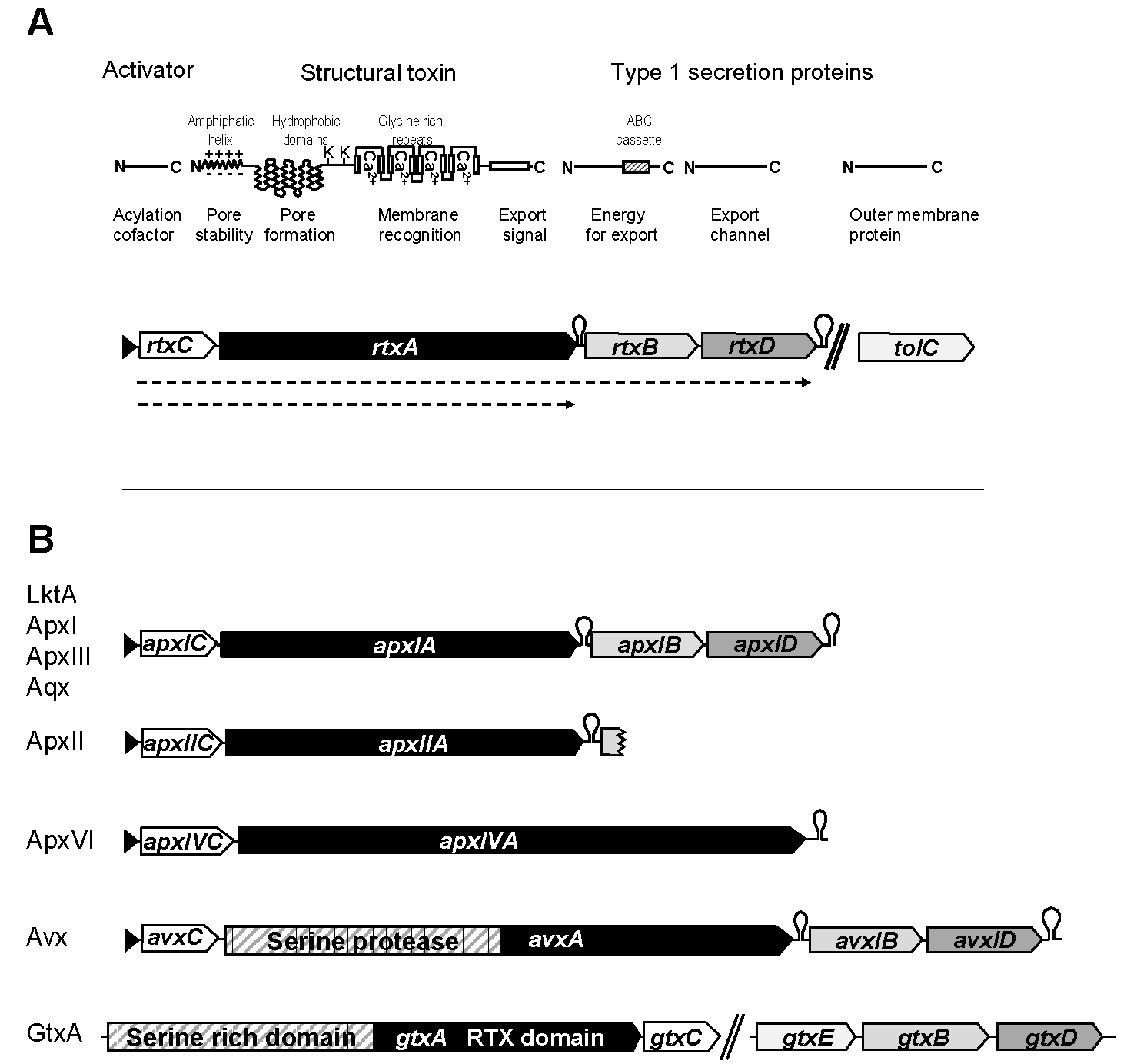
2.2. ApxI, ApxII, ApxIII, and ApxIV from Actinobacillus Pleuropneumoniae, a Multi-RTX Toxin-Producing Pathogen Causing Porcine Pleuropneumonia
2.3. ApxI and ApxII Toxins in Actinobacillus Suis and Actinobacillus Porcitonsillarum
2.4. Aqx Toxin from Equine Actinobacillus Equuli
2.5. GtxA MARTX from the Poultry Pathogen Gallibacterium Anatis
2.6. AvxA MARTX from Poultry Pathogen Avibacterium Paragallinarum
3. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Welch, R.A. Pore-forming cytolysins of gram-negative bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Baumann, U.; Wu, S.; Flaherty, K.M.; McKay, D.B. Three-dimensional structure of the alkaline protease of Pseudomonas aeruginosa: A two-domain protein with a calcium binding parallel beta roll motif. EMBO J. 1993, 12, 3357–3364. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Vance, T.D.R.; Stevens, C.A.; Voets, I.K.; Davies, P.L. RTX Adhesins are Key Bacterial Surface Megaproteins in the Formation of Biofilms. Trends Microbiol. 2019, 27, 470. [Google Scholar] [CrossRef] [PubMed]
- Ostolaza, H.; Gonzalez-Bullon, D.; Uribe, K.B.; Martin, C.; Amuategi, J.; Fernandez-Martinez, X. Membrane Permeabilization by Pore-Forming RTX Toxins: What Kind of Lesions Do These Toxins Form? Toxins 2019, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Goebel, W.; Hedgpeth, J. Cloning and functional characterization of the plasmid-encoded hemolysin determinant of Escherichia coli. J. Bacteriol. 1982, 151, 1290–1298. [Google Scholar] [PubMed]
- Hacker, J.; Hughes, C.; Hof, H.; Goebel, W. Cloned hemolysin genes from Escherichia coli that cause urinary tract infection determine different levels of toxicity in mice. Infect. Immun. 1983, 42, 57–63. [Google Scholar]
- Hughes, C.; Muller, D.; Hacker, J.; Goebel, W. Genetics and pathogenic role of Escherichia coli haemolysin. Toxicon 1982, 20, 247–252. [Google Scholar] [CrossRef]
- Koronakis, V.; Hughes, C. Synthesis, maturation and export of the E. coli hemolysin. Med. Microbiol. Immunol. 1996, 185, 65–71. [Google Scholar] [CrossRef]
- Stanley, P.; Koronakis, V.; Hughes, C. Acylation of Escherichia coli hemolysin: A unique protein lipidation mechanism underlying toxin function. Microbiol. Mol. Biol. Rev. 1998, 62, 309–333. [Google Scholar]
- Fullner, K.J.; Mekalanos, J.J. In vivo covalent cross-linking of cellular actin by the Vibrio cholerae RTX toxin. EMBO J. 2000, 19, 5315–5323. [Google Scholar] [CrossRef]
- Gygi, D.; Nicolet, J.; Frey, J.; Cross, M.; Koronakis, V.; Hughes, C. Isolation of the Actinobacillus pleuropneumoniae haemolysin gene and the activation and secretion of the prohaemolysin by the HlyC, HlyB and HlyD proteins of Escherichia coli. Mol. Microbiol. 1990, 4, 123–128. [Google Scholar] [CrossRef]
- Koronakis, V.; Cross, M.; Hughes, C. Expression of the E. coli hemolysin secretion gene hlyB involves transcript anti-termination within the hly operon. Nucleic Acids Res. 1988, 16, 4789–4800. [Google Scholar] [PubMed]
- Koronakis, V.; Eswaran, J.; Hughes, C. Structure and function of TolC: The bacterial exit duct for proteins and drugs. Annu. Rev. Biochem. 2004, 73, 467–489. [Google Scholar] [CrossRef] [PubMed]
- Koronakis, V.; Stanley, P.; Koronakis, E.; Hughes, C. The HlyB/HlyD-dependent secretion of toxins by gram-negative bacteria. FEMS Microbiol. Immunol. 1992, 5, 45–53. [Google Scholar] [CrossRef]
- Koronakis, V.; Hughes, C. Bacterial signal peptide-independent protein export: HlyB-directed secretion of hemolysin. Semin. Cell Biol. 1993, 4, 7–15. [Google Scholar] [CrossRef]
- Delepelaire, P. Type I secretion in gram-negative bacteria. Biochim. Biophys. Acta 2004, 1694, 149–161. [Google Scholar] [CrossRef]
- Benabdelhak, H.; Kiontke, S.; Horn, C.; Ernst, R.; Blight, M.A.; Holland, I.B.; Schmitt, L. A specific interaction between the NBD of the ABC-transporter HlyB and a C-terminal fragment of its transport substrate haemolysin A. J. Mol. Biol. 2003, 327, 1169–1179. [Google Scholar] [CrossRef]
- Holland, I.B.; Blight, M.A.; Kenny, B. The mechanism of secretion of hemolysin and other polypeptides from gram-negative bacteria. J. Bioenerg. Biomembr. 1990, 22, 473–491. [Google Scholar] [CrossRef]
- Koronakis, E.; Hughes, C.; Milisav, I.; Koronakis, V. Protein exporter function and in vitro ATPase activity are correlated in ABC-domain mutants of HlyB. Mol. Microbiol. 1995, 16, 87–96. [Google Scholar] [CrossRef]
- Frey, J.; Haldimann, A.; Nicolet, J.; Boffini, A.; Prentki, P. Sequence analysis and transcription of the apxI operon (hemolysin I) from Actinobacillus pleuropneumoniae. Gene 1994, 142, 97–102. [Google Scholar] [CrossRef]
- Welch, R.A. RTX toxin structure and function: A story of numerous anomalies and few analogies in toxin biology. Curr. Top. Microbiol. Immunol. 2001, 257, 85–111. [Google Scholar] [PubMed]
- Aulik, N.A.; Hellenbrand, K.M.; Czuprynski, C.J. Mannheimia haemolytica and its leukotoxin cause macrophage extracellular trap formation by bovine macrophages. Infect. Immun. 2012, 80, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Balashova, N.; Dhingra, A.; Boesze-Battaglia, K.; Lally, E.T. Aggregatibacter actinomycetemcomitans leukotoxin induces cytosol acidification in LFA-1 expressing immune cells. Mol. Oral Microbiol. 2016, 31, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Lally, E.T.; Hill, R.B.; Kieba, I.R.; Korostoff, J. The interaction between RTX toxins and target cells. Trends Microbiol. 1999, 7, 356–361. [Google Scholar] [CrossRef]
- Zecchinon, L.; Fett, T.; Desmecht, D. How Mannheimia haemolytica defeats host defence through a kiss of death mechanism. Vet. Res. 2005, 36, 133–156. [Google Scholar] [CrossRef]
- Confer, A.W.; Panciera, R.J.; Clinkenbeard, K.D.; Mosier, D.A. Molecular aspects of virulence of Pasteurella haemolytica. Can. J. Vet. Res. 1990, 54, S48–S52. [Google Scholar]
- Atapattu, D.N.; Czuprynski, C.J. Mannheimia haemolytica leukotoxin binds to lipid rafts in bovine lymphoblastoid cells and is internalized in a dynamin-2- and clathrin-dependent manner. Infect. Immun. 2007, 75, 4719–4727. [Google Scholar] [CrossRef]
- Raksanoh, V.; Prangkio, P.; Imtong, C.; Thamwiriyasati, N.; Suvarnapunya, K.; Shank, L.; Angsuthanasombat, C. Structural requirement of the hydrophobic region of the Bordetella pertussis CyaA-hemolysin for functional association with CyaC-acyltransferase in toxin acylation. Biochem. Biophys. Res. Commun. 2018, 499, 862–867. [Google Scholar] [CrossRef]
- Osickova, A.; Balashova, N.; Masin, J.; Sulc, M.; Roderova, J.; Wald, T.; Brown, A.C.; Koufos, E.; Chang, E.H.; Giannakakis, A.; et al. Cytotoxic activity of Kingella kingae RtxA toxin depends on post-translational acylation of lysine residues and cholesterol binding. Emerg. Microbes Infect. 2018, 7, 178. [Google Scholar] [CrossRef]
- O’Brien, D.P.; Cannella, S.E.; Voegele, A.; Raoux-Barbot, D.; Davi, M.; Douche, T.; Matondo, M.; Brier, S.; Ladant, D.; Chenal, A. Post-translational acylation controls the folding and functions of the CyaA RTX toxin. FASEB J. 2019, 33, 10065–10076. [Google Scholar] [CrossRef]
- Meetum, K.; Imtong, C.; Katzenmeier, G.; Angsuthanasombat, C. Acylation of the Bordetella pertussis CyaA-hemolysin: Functional implications for efficient membrane insertion and pore formation. Biochim. Biophys. Acta Biomembr. 2017, 1859, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Bellalou, J.; Sakamoto, H.; Ladant, D.; Geoffroy, C.; Ullmann, A. Deletions affecting hemolytic and toxin activities of Bordetella pertussis adenylate cyclase. Infect. Immun. 1990, 58, 3242–3247. [Google Scholar] [PubMed]
- Basler, M.; Knapp, O.; Masin, J.; Fiser, R.; Maier, E.; Benz, R.; Sebo, P.; Osicka, R. Segments crucial for membrane translocation and pore-forming activity of Bordetella adenylate cyclase toxin. J. Biol. Chem. 2007, 282, 12419–12429. [Google Scholar] [CrossRef]
- Ludwig, A.; Volkerink, G.; von Rhein, C.; Bauer, S.; Maier, E.; Bergmann, B.; Goebel, W.; Benz, R. Mutations affecting export and activity of cytolysin A from Escherichia coli. J. Bacteriol. 2010, 192, 4001–4011. [Google Scholar] [CrossRef]
- Strathdee, C.A.; Lo, R.Y. Cloning, nucleotide sequence, and characterization of genes encoding the secretion function of the Pasteurella haemolytica leukotoxin determinant. J. Bacteriol. 1989, 171, 916–928. [Google Scholar] [CrossRef]
- Strathdee, C.A.; Lo, R.Y. Regulation of expression of the Pasteurella haemolytica leukotoxin determinant. J. Bacteriol. 1989, 171, 5955–5962. [Google Scholar] [CrossRef]
- Burrows, L.L.; Olah-Winfield, E.; Lo, R.Y. Molecular analysis of the leukotoxin determinants from Pasteurella haemolytica serotypes 1 to 16. Infect. Immun. 1993, 61, 5001–5007. [Google Scholar]
- Miller, M.W.; Hause, B.M.; Killion, H.J.; Fox, K.A.; Edwards, W.H.; Wolfe, L.L. Phylogenetic and epidemiologic relationships among Pasteurellaceae from Colorado bighorn sheep herds. J. Wildl. Dis. 2013, 49, 653–660. [Google Scholar] [CrossRef][Green Version]
- Tatum, F.M.; Briggs, R.E.; Sreevatsan, S.S.; Zehr, E.S.; Ling Hsuan, S.; Whiteley, L.O.; Ames, T.R.; Maheswaran, S.K. Construction of an isogenic leukotoxin deletion mutant of Pasteurella haemolytica serotype 1: Characterization and virulence. Microb. Pathog. 1998, 24, 37–46. [Google Scholar] [CrossRef]
- Czuprynski, C.J.; Noel, E.J.; Ortiz-Carranza, O.; Srikumaran, S. Activation of bovine neutrophils by partially purified Pasteurella haemolytica leukotoxin. Infect. Immun. 1991, 59, 3126–3133. [Google Scholar]
- Cudd, L.A.; Ownby, C.L.; Clarke, C.R.; Sun, Y.; Clinkenbeard, K.D. Effects of Mannheimia haemolytica leukotoxin on apoptosis and oncosis of bovine neutrophils. Am. J. Vet. Res. 2001, 62, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Highlander, S.K. Molecular genetic analysis of virulence in Mannheimia (pasteurella) haemolytica. Front. Biosci. 2001, 6, D1128–D1150. [Google Scholar] [PubMed]
- Gioia, J.; Qin, X.; Jiang, H.; Clinkenbeard, K.; Lo, R.; Liu, Y.; Fox, G.E.; Yerrapragada, S.; McLeod, M.P.; McNeill, T.Z.; et al. The genome sequence of Mannheimia haemolytica A1: Insights into virulence, natural competence, and Pasteurellaceae phylogeny. J. Bacteriol. 2006, 188, 7257–7266. [Google Scholar] [CrossRef]
- Shewen, P.E.; Wilkie, B.N. Cytotoxin of Pasteurella haemolytica acting on bovine leukocytes. Infect. Immun. 1982, 35, 91–94. [Google Scholar] [PubMed]
- Davies, R.L.; Campbell, S.; Whittam, T.S. Mosaic structure and molecular evolution of the leukotoxin operon (lktCABD) in Mannheimia (Pasteurella) haemolytica, Mannheimia glucosida, and Pasteurella trehalosi. J. Bacteriol. 2002, 184, 266–277. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Davies, R.L.; Baillie, S. Cytotoxic activity of Mannheimia haemolytica strains in relation to diversity of the leukotoxin structural gene lktA. Vet. Microbiol. 2003, 92, 263–279. [Google Scholar] [CrossRef]
- Larsen, J.; Kuhnert, P.; Frey, J.; Christensen, H.; Bisgaard, M.; Olsen, J.E. Analysis of gene order data supports vertical inheritance of the leukotoxin operon and genome rearrangements in the 5′ flanking region in genus Mannheimia. BMC Evol. Biol. 2007, 7, 184. [Google Scholar] [CrossRef]
- Larsen, J.; Pedersen, A.G.; Davies, R.L.; Kuhnert, P.; Frey, J.; Christensen, H.; Bisgaard, M.; Olsen, J.E. Evolution of the leukotoxin promoter in genus Mannheimia. BMC Evol. Biol. 2009, 9, 121. [Google Scholar] [CrossRef]
- Larsen, J.; Pedersen, A.G.; Christensen, H.; Bisgaard, M.; Angen, O.; Ahrens, P.; Olsen, J.E. Evidence for vertical inheritance and loss of the leukotoxin operon in genus Mannheimia. J. Mol. Evol. 2007, 64, 423–437. [Google Scholar] [CrossRef]
- Dassanayake, R.P.; Maheswaran, S.K.; Srikumaran, S. Monomeric expression of bovine beta2-integrin subunits reveals their role in Mannheimia haemolytica leukotoxin-induced biological effects. Infect. Immun. 2007, 75, 5004–5010. [Google Scholar] [CrossRef]
- Dassanayake, R.P.; Shanthalingam, S.; Davis, W.C.; Srikumaran, S. Mannheimia haemolytica leukotoxin-induced cytolysis of ovine (Ovis aries) leukocytes is mediated by CD18, the beta subunit of beta2-integrins. Microb. Pathog. 2007, 42, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, M.S.; Ambagala, T.C.; Ambagala, A.P.N.; Kehrli, M.E.J.; Srikumaran, S. Bovine CD18 is necessary and sufficient to mediate Mannheimia (Pasteurella) haemolytica leukotoxin-induced cytolysis. Infect. Immun. 2002, 70, 5058–5064. [Google Scholar] [CrossRef] [PubMed]
- Zecchinon, L.; Fett, T.; Baise, E.; Desmecht, D. Molecular cloning and characterisation of the CD18 partner in ovine (Ovis aries) beta2-integrins. Gene 2004, 334, 47–52. [Google Scholar] [CrossRef]
- Zecchinon, L.; Fett, T.; Baise, E.; Desmecht, D. Characterization of the caprine (Capra hircus) beta-2 integrin CD18-encoding cDNA and identification of mutations potentially responsible for the ruminant-specific virulence of Mannheimia haemolytica. Mol. Membr. Biol. 2004, 21, 289–295. [Google Scholar] [CrossRef]
- Dileepan, T.; Kannan, M.S.; Walcheck, B.; Maheswaran, S.K. Integrin-EGF-3 domain of bovine CD18 is critical for Mannheimia haemolytica leukotoxin species-specific susceptibility. FEMS Microbiol. Lett. 2007, 274, 67–72. [Google Scholar] [CrossRef][Green Version]
- Dileepan, T.; Kannan, M.S.; Walcheck, B.; Thumbikat, P.; Maheswaran, S.K. Mapping of the binding site for Mannheimia haemolytica leukotoxin within bovine CD18. Infect. Immun. 2005, 73, 5233–5237. [Google Scholar] [CrossRef]
- Workman, A.M.; Chitko-McKown, C.G.; Smith, T.P.L.; Bennett, G.L.; Kalbfleisch, T.S.; Basnayake, V.; Heaton, M.P. A bovine CD18 signal peptide variant with increased binding activity to Mannheimia hemolytica leukotoxin. F1000Research 2018, 7, 1985. [Google Scholar] [CrossRef]
- Atapattu, D.N.; Albrecht, R.M.; McClenahan, D.J.; Czuprynski, C.J. Dynamin-2-dependent targeting of mannheimia haemolytica leukotoxin to mitochondrial cyclophilin D in bovine lymphoblastoid cells. Infect. Immun. 2008, 76, 5357–5365. [Google Scholar] [CrossRef]
- Atapattu, D.N.; Czuprynski, C.J. Mannheimia haemolytica leukotoxin induces apoptosis of bovine lymphoblastoid cells (BL-3) via a caspase-9-dependent mitochondrial pathway. Infect. Immun. 2005, 73, 5504–5513. [Google Scholar] [CrossRef]
- Kisiela, D.I.; Aulik, N.A.; Atapattu, D.N.; Czuprynski, C.J. N-terminal region of Mannheimia haemolytica leukotoxin serves as a mitochondrial targeting signal in mammalian cells. Cell. Microbiol. 2010, 12, 976–987. [Google Scholar] [CrossRef]
- Confer, A.W.; Ayalew, S. Mannheimia haemolytica in bovine respiratory disease: Immunogens, potential immunogens, and vaccines. Anim. Heal. Res. Rev. 2018, 19, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, B.; Bavananthasivam, J.; Kugadas, A.; Haldorson, G.J.; Srikumaran, S. Effect of vaccination against pneumonia on the survival of bighorn sheep (Ovis canadensis) commingled with carrier animals. Vet. Microbiol. 2017, 203, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bosch, H.; Frey, J. Interference of outer membrane protein PalA with protective immunity against Actinobacillus pleuropneumoniae infections in vaccinated pigs. Vaccine 2003, 21, 3601–3607. [Google Scholar] [CrossRef]
- Dreyfus, A.; Schaller, A.; Nivollet, S.; Segers, R.P.A.M.; Kobisch, M.; Mieli, L.; Soerensen, V.; Hüssy, D.; Miserez, R.; Zimmermann, W.; et al. Use of recombinant ApxIV in serodiagnosis of Actinobacillus pleuropneumoniae infections, development and prevalidation of the ApxIV ELISA. Vet. Microbiol. 2004, 99, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Frey, J.; Beck, M.; van den Bosch, J.F.; Segers, R.P.A.M.; Nicolet, J. Development of an efficient PCR method for toxin typing of Actinobacillus pleuropneumoniae strains. Mol. Cell. Probes 1995, 9, 277–282. [Google Scholar] [CrossRef]
- Schaller, A.; Djordjevic, S.P.; Eamens, G.J.; Forbes, W.A.; Kuhn, R.; Kuhnert, P.; Gottschalk, M.; Nicolet, J.; Frey, J. Identification and detection of Actinobacillus pleuropneumoniae by PCR based on the gene apxIVA. Vet. Microbiol. 2001, 79, 47–62. [Google Scholar] [CrossRef]
- Berthoud, H.; Frey, J.; Kuhnert, P. Characterization of Aqx and its operon: The hemolytic RTX determinant of Actinobacillus equuli. Vet. Microbiol. 2002, 87, 159–174. [Google Scholar] [CrossRef]
- Kung, E.; Frey, J. AvxA, a composite serine-protease-RTX toxin of Avibacterium paragallinarum. Vet. Microbiol. 2013, 163, 290–298. [Google Scholar] [CrossRef]
- Persson, G.; Pors, S.E.; Thofner, I.C.N.; Bojesen, A.M. Vaccination with outer membrane vesicles and the fimbrial protein FlfA offers improved protection against lesions following challenge with Gallibacterium anatis. Vet. Microbiol. 2018, 217, 104–111. [Google Scholar] [CrossRef]
- Loy, J.D.; Leger, L.; Workman, A.M.; Clawson, M.L.; Bulut, E.; Wang, B. Development of a multiplex real-time PCR assay using two thermocycling platforms for detection of major bacterial pathogens associated with bovine respiratory disease complex from clinical samples. J. Vet. Diagn. Invest. 2018, 30, 837–847. [Google Scholar] [CrossRef]
- Thanthrige-Don, N.; Lung, O.; Furukawa-Stoffer, T.; Buchanan, C.; Joseph, T.; Godson, D.L.; Gilleard, J.; Alexander, T.; Ambagala, A. A novel multiplex PCR-electronic microarray assay for rapid and simultaneous detection of bovine respiratory and enteric pathogens. J. Virol. Methods 2018, 261, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Bosse, J.T.; Li, Y.; Fernandez Crespo, R.; Lacouture, S.; Gottschalk, M.; Sarkozi, R.; Fodor, L.; Casas Amoribieta, M.; Angen, O.; Nedbalcova, K.; et al. Comparative sequence analysis of the capsular polysaccharide loci of Actinobacillus pleuropneumoniae serovars 1–18, and development of two multiplex PCRs for comprehensive capsule typing. Vet. Microbiol. 2018, 220, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Bosse, J.T.; Li, Y.; Sarkozi, R.; Fodor, L.; Lacouture, S.; Gottschalk, M.; Casas Amoribieta, M.; Angen, O.; Nedbalcova, K.; Holden, M.T.G.; et al. Proposal of serovars 17 and 18 of Actinobacillus pleuropneumoniae based on serological and genotypic analysis. Vet. Microbiol. 2018, 217, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Van den Bosch, J.F.; Jongenelen, I.M.C.A.; Loeffen, P.L.W.; Nielsen, R.; Nicolet, J.; Frey, J. RTX toxin genotypes and phenotypes in Actinobacillus pleuropneumoniae field strains. J. Clin. Microbiol. 1994, 32. [Google Scholar]
- Reimer, D.; Frey, J.; Jansen, R.; Veit, H.P.; Inzana, T.J. Molecular investigation of the role of Apxl and Apxll in the virulence of Actinobacillus pleuropneumoniae serotype 5. Microb. Pathog. 1995, 18, 197–209. [Google Scholar] [CrossRef]
- Frey, J. Virulence in Actinobacillus pleuropneumoniae and RTX toxins. Trends Microbiol. 1995, 3, 257–261. [Google Scholar] [CrossRef]
- Kuhnert, P.; Schlatter, Y.; Frey, J. Characterization of the type I secretion system of the RTX toxin ApxII in “Actinobacillus porcitonsillarum”. Vet. Microbiol. 2005, 107, 225–232. [Google Scholar] [CrossRef]
- Dona, V.; Perreten, V. Comparative Genomics of the First and Complete Genome of Actinobacillus porcitonsillarum Supports the Novel Species Hypothesis. Int. J. Genom. 2018, 2018, 5261719. [Google Scholar] [CrossRef]
- Tascon, R.I.; Vazquez-Boland, J.A.; Gutierrez-Martin, C.B.; Rodriguez-Barbosa, I.; Rodriguez-Ferri, E.F. The RTX haemolysins ApxI and ApxII are major virulence factors of the swine pathogen Actinobacillus pleuropneumoniae: Evidence from mutational analysis. Mol. Microbiol. 1994, 14, 207–216. [Google Scholar] [CrossRef]
- Boekema, B.K.H.L.; Kamp, E.M.; Smits, M.A.; Smith, H.E.; Stockhofe-Zurwieden, N. Both ApxI and ApxII of Actinobacillus pleuropneumoniae serotype 1 are necessary for full virulence. Vet. Microbiol. 2004, 100, 17–23. [Google Scholar] [CrossRef]
- Kamp, E.M.; Popma, J.K.; Anakotta, J.; Smits, M.A. Identification of hemolytic and cytotoxic proteins of Actinobacillus pleuropneumoniae by use of monoclonal antibodies. Infect. Immun. 1991, 59, 3079–3085. [Google Scholar] [PubMed]
- Dom, P.; Haesebrouck, F.; De Baetselier, P. Stimulation and suppression of the oxygenation activity of porcine pulmonary alveolar macrophages by Actinobacillus pleuropneumoniae and its metabolites. Am. J. Vet. Res. 1992, 53, 1113–1118. [Google Scholar] [PubMed]
- Bosse, J.T.; Janson, H.; Sheehan, B.J.; Beddek, A.J.; Rycroft, A.N.; Kroll, J.S.; Langford, P.R. Actinobacillus pleuropneumoniae: Pathobiology and pathogenesis of infection. Microbes Infect. 2002, 4, 225–235. [Google Scholar] [CrossRef]
- Vanden Bergh, P.G.; Zecchinon, L.L.; Fett, T.; Desmecht, D. Probing of Actinobacillus pleuropneumoniae ApxIIIA toxin-dependent cytotoxicity towards mammalian peripheral blood mononucleated cells. BMC Res. Notes 2008, 1, 121. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bergh, P.G.A.C.; Zecchinon, L.L.M.; Fett, T.; Desmecht, D. Porcine CD18 mediates Actinobacillus pleuropneumoniae ApxIII species-specific toxicity. Vet. Res. 2009, 40, 33. [Google Scholar] [CrossRef]
- Maier, E.; Reinhard, N.; Benz, R.; Frey, J. Channel-forming activity and channel size of the RTX toxins ApxI, ApxII, and ApxIII of Actinobacillus pleuropneumoniae. Infect. Immun. 1996, 64, 4415–4423. [Google Scholar]
- Osicka, R.; Prochazkova, K.; Sulc, M.; Linhartova, I.; Havlicek, V.; Sebo, P. A novel clip-and-link activity of repeat in toxin (RTX) proteins from gram-negative pathogens. Covalent protein cross-linking by an Asp-Lys isopeptide bond upon calcium-dependent processing at an Asp-Pro bond. J. Biol. Chem. 2004, 279, 24944–24956. [Google Scholar] [CrossRef]
- Schaller, A.; Kuhn, R.; Kuhnert, P.; Nicolet, J.; Anderson, T.J.; MacInnes, J.I.; Segers, R.P.A.M.; Frey, J. Characterization of apxIVA, a new RTX determinant of Actinobacillus pleuropneumoniae. Microbiology 1999, 145, 2105–2116. [Google Scholar] [CrossRef]
- Deslandes, V.; Denicourt, M.; Girard, C.; Harel, J.; Nash, J.H.E.; Jacques, M. Transcriptional profiling of Actinobacillus pleuropneumoniae during the acute phase of a natural infection in pigs. BMC Genom. 2010, 11, 98. [Google Scholar] [CrossRef]
- MacInnes, J.I.; Gottschalk, M.; Lone, A.G.; Metcalf, D.S.; Ojha, S.; Rosendal, T.; Watson, S.B.; Friendship, R.M. Prevalence of Actinobacillus pleuropneumoniae, Actinobacillus suis, Haemophilus parasuis, Pasteurella multocida, and Streptococcus suis in representative Ontario swine herds. Can. J. Vet. Res. 2008, 72, 242–248. [Google Scholar]
- Loera-Muro, V.M.; Jacques, M.; Tremblay, Y.D.N.; Avelar-Gonzalez, F.J.; Loera Muro, A.; Ramirez-Lopez, E.M.; Medina-Figueroa, A.; Gonzalez-Reynaga, H.M.; Guerrero-Barrera, A.L. Detection of Actinobacillus pleuropneumoniae in drinking water from pig farms. Microbiology 2013, 159, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jones, S.C.P.; Angen, Ø.; Bossé, J.T.; Nash, J.H.E.; Frey, J.; Zhou, R.; Chen, H.C.; Kroll, J.S.; Rycroft, A.N.; et al. Multiplex PCR that can distinguish between immunologically cross-reactive serovar 3, 6, and 8 Actinobacillus pleuropneumoniae strains. J. Clin. Microbiol. 2008, 46, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Bosse, J.T.; Li, Y.; Angen, O.; Weinert, L.A.; Chaudhuri, R.R.; Holden, M.T.; Williamson, S.M.; Maskell, D.J.; Tucker, A.W.; Wren, B.W.; et al. Multiplex PCR assay for unequivocal differentiation of Actinobacillus pleuropneumoniae serovars 1 to 3, 5 to 8, 10, and 12. J. Clin. Microbiol. 2014, 52, 2380–2385. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dousse, F.; Thomann, A.; Brodard, I.; Korczak, B.M.; Schlatter, Y.; Kuhnert, P.; Miserez, R.; Frey, J. Routine phenotypic identification of bacterial species of the family Pasteurellaceae isolated from animals. J. Vet. Diagn. Investig. 2008, 20, 716–724. [Google Scholar] [CrossRef]
- Gonzalez, W.; Gimenez-Lirola, L.G.; Holmes, A.; Lizano, S.; Goodell, C.; Poonsuk, K.; Sitthicharoenchai, P.; Sun, Y.; Zimmerman, J. Detection of Actinobacillus Pleuropneumoniae ApxIV Toxin Antibody in Serum and Oral Fluid Specimens from Pigs Inoculated Under Experimental Conditions. J. Vet. Res. 2017, 61, 163–171. [Google Scholar] [CrossRef]
- Capua, I.; Cattoli, G.; Marangon, S. DIVA–A vaccination strategy enabling the detection of field exposure to avian influenza. Dev. Biol. 2004, 119, 229–233. [Google Scholar]
- Gottschalk, M.; Broes, A.; Mittal, K.R.; Kobisch, M.; Kuhnert, P.; Lebrun, A.; Frey, J. Non-pathogenic Actinobacillus isolates antigenically and biochemically similar to Actinobacillus pleuropneumoniae: A novel species? Vet. Microbiol. 2003, 92, 87–101. [Google Scholar] [CrossRef]
- Holyoake, P.K.; Thompson, A. Isolation of [Actinobacillus] rossii from an aborted piglet. Aust. Vet. J. 2017, 95, 483–485. [Google Scholar] [CrossRef]
- Mayor, D.; Korczak, B.M.; Christensen, H.; Bisgaard, M.; Frey, J.; Kuhnert, P. Distribution of RTX toxin genes in strains of [Actinobacillus] rossii and [Pasteurella] mairii. Vet. Microbiol. 2006, 116, 194–201. [Google Scholar] [CrossRef]
- Burrows, L.L.; Lo, R.Y. Molecular characterization of an RTX toxin determinant from Actinobacillus suis. Infect. Immun. 1992, 60, 2166–2173. [Google Scholar]
- Van Ostaaijen, J.; Frey, J.; Rosendal, S.; MacInnes, J.I. Actinobacillus suis strains isolated from healthy and diseased swine are clonal and carry apxICABD(var. suis) and apxIICA(var. suis) toxin genes. J. Clin. Microbiol. 1997, 35, 1131–1137. [Google Scholar]
- Christensen, H.; Bisgaard, M.; Olsen, J.E. Reclassification of equine isolates previously reported as Actinobacillus equuli, variants of A. equuli, Actinobacillus suis or Bisgaard taxon 11 and proposal of A. equuli subsp. equuli subsp. nov. and A. equuli subsp. haemolyticus subsp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1569–1576. [Google Scholar]
- Rycroft, A.N.; Garside, L.H. Actinobacillus species and their role in animal disease. Vet. J. 2000, 159, 18–36. [Google Scholar] [CrossRef]
- Huang, B.F.; Kropinski, A.M.; Bujold, A.R.; MacInnes, J.I. Complete genome sequence of Actinobacillus equuli subspecies equuli ATCC 19392(T). Stand. Genomic Sci. 2015, 10, 32. [Google Scholar] [CrossRef]
- Kuhnert, P.; Berthoud, H.; Christensen, H.; Bisgaard, M.; Frey, J. Phylogenetic relationship of equine Actinobacillus species and distribution of RTX toxin genes among clusters. Vet. Res. 2003, 34, 353–359. [Google Scholar] [CrossRef][Green Version]
- Pusterla, N.; Jones, M.E.B.; Mohr, F.C.; Higgins, J.K.; Mapes, S.; Jang, S.S.; Samitz, E.M.; Byrne, B.A. Fatal pulmonary hemorrhage associated with RTX toxin producing Actinobacillus equuli subspecies haemolyticus infection in an adult horse. J. Vet. Diagn. Invest. 2008, 20, 118–121. [Google Scholar] [CrossRef]
- Kuhnert, P.; Berthoud, H.; Straub, R.; Frey, J. Host cell specific activity of RTX toxins from haemolytic Actinobacillus equuli and Actinobacillus suis. Vet. Microbiol. 2003, 92, 161–167. [Google Scholar] [CrossRef]
- Skaggs, J.S.; Reed Holyoak, G.; Montelongo, M.; Ayalew, S.; Confer, A.W. Serum antibody responses in horses and mice following immunization with Actinobacillus equuli outer membrane proteins and recombinant Aqx toxin. Vet. J. 2011, 190, e91–e94. [Google Scholar] [CrossRef]
- Berthoud, H.; Frey, J.; Sternberg, S.; Straub, R.; Kuhnert, P. Antibodies to Aqx toxin of Actinobacillus equuli in horses and foals. Vet. Rec. 2004, 155, 231–233. [Google Scholar] [CrossRef]
- Rycroft, A.N.; Woldeselassie, A.; Gordon, P.J.; Bjornson, A. Serum antibody in equine neonatal septicaemia due to Actinobacillus equuli. Vet. Rec. 1998, 143, 254–255. [Google Scholar] [CrossRef]
- Wang, C.; Pors, S.E.; Olsen, R.H.; Bojesen, A.M. Transmission and pathogenicity of Gallibacterium anatis and Escherichia coli in embryonated eggs. Vet. Microbiol. 2018, 217, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Ruhnau, D.; Wernsdorf, P.; Liebhart, D.; Hess, M.; Hess, C. Presence of Avibacterium paragallinarum and Histopathologic Lesions Corresponds with Clinical Signs in a Co-infection Model with Gallibacterium anatis. Avian Dis. 2017, 61, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial Resistance in Bacterial Poultry Pathogens: A Review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar] [CrossRef] [PubMed]
- Bojesen, A.M.; Vazquez, M.E.; Bager, R.J.; Ifrah, D.; Gonzalez, C.; Aarestrup, F.M. Antimicrobial susceptibility and tetracycline resistance determinant genotyping of Gallibacterium anatis. Vet. Microbiol. 2011, 148, 105–110. [Google Scholar] [CrossRef]
- Kristensen, B.M.; Frees, D.; Bojesen, A.M. Expression and secretion of the RTX-toxin GtxA among members of the genus Gallibacterium. Vet. Microbiol. 2011, 153, 116–123. [Google Scholar] [CrossRef]
- Vojtova, J.; Kamanova, J.; Sebo, P. Bordetella adenylate cyclase toxin: A swift saboteur of host defense. Curr. Opin. Microbiol. 2006, 9, 69–75. [Google Scholar] [CrossRef]
- Kristensen, B.M.; Frees, D.; Bojesen, A.M. GtxA from Gallibacterium anatis, a cytolytic RTX-toxin with a novel domain organisation. Vet. Res. 2010, 41, 25. [Google Scholar] [CrossRef]
- Merbach, S.; Peters, M.; Kilwinski, J.R.D. Acute necrotising pneumonitis associated with Suttonella ornithocola infection in tits (Paridae). Berl. Münch. Tierärztl. Wochenschc. 2019, 132, 459–463. [Google Scholar]
- Lawson, B.; Malnick, H.; Pennycott, T.W.; Macgregor, S.K.; John, S.K.; Duncan, G.; Hughes, L.A.; Chantrey, J.; Cunningham, A.A. Acute necrotising pneumonitis associated with Suttonella ornithocola infection in tits (Paridae). Vet. J. 2011, 188, 96–100. [Google Scholar] [CrossRef]
- Pors, S.E.; Skjerning, R.B.; Flachs, E.M.; Bojesen, A.M. Recombinant proteins from Gallibacterium anatis induces partial protection against heterologous challenge in egg-laying hens. Vet. Res. 2016, 47, 36. [Google Scholar] [CrossRef]
- Pors, S.E.; Pedersen, I.J.; Skjerning, R.B.; Thofner, I.C.N.; Persson, G.; Bojesen, A.M. Outer membrane vesicles of Gallibacterium anatis induce protective immunity in egg-laying hens. Vet. Microbiol. 2016, 195, 123–127. [Google Scholar] [CrossRef]
- Pedersen, I.J.; Pors, S.E.; Bager Skjerning, R.J.; Nielsen, S.S.; Bojesen, A.M. Immunogenic and protective efficacy of recombinant protein GtxA-N against Gallibacterium anatis challenge in chickens. Avian Pathol. 2015, 44, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Blackall, P.J. Infectious coryza: Overview of the disease and new diagnostic options. Clin. Microbiol. Rev. 1999, 12, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Blackall, P.J.; Christensen, H.; Beckenham, T.; Blackall, L.L.; Bisgaard, M. Reclassification of Pasteurella gallinarum, [Haemophilus] paragallinarum, Pasteurella avium and Pasteurella volantium as Avibacterium gallinarum gen. nov., comb. nov., Avibacterium paragallinarum comb. nov., Avibacterium avium comb. nov. and Avibacterium. Int. J. Syst. Evol. Microbiol. 2005, 55, 353–362. [Google Scholar] [CrossRef]
- Pan, Y.-C.; Tan, D.-H.; Shien, J.-H.; Liu, C.-C.; He, Y.-S.; Shen, P.-C.; Chang, P.-C. Identification and characterization of an RTX toxin-like gene and its operon from Avibacterium paragallinarum. Avian Dis. 2012, 56, 537–544. [Google Scholar] [CrossRef]
- Frey, J. Genetics and Phylogeny of RTX Cytolysins; Elsevier: Amsterdam, The Netherlands, 2006; ISBN 9780120884452. [Google Scholar]


{kind=link}
{kind=link}
| RTX Toxin rtx Gene | Species | Use in Veterinary Medicine |
|---|---|---|
| ApxI | A. pleuropneumoniae | Antigen in commercial, universal serovar vaccines against porcine pleuropneumonia [63] |
| ApxII | A. pleuropneumoniae | Antigen in commercial, universal serovar vaccines against porcine pleuropneumonia [63] |
| ApxIII | A. pleuropneumoniae | Antigen in commercial, universal serovar vaccines against porcine pleuropneumonia [63] |
| ApxIV | A. pleuropneumoniae | Recombinant ApxIV ELISA for sero-detection of A. pleuropneumoniae infected pigs [64] |
| apxI | A. pleuropneumoniae | Diagnostic PCR for toxin typing of A. pleuropneumoniae strains [65] |
| apxII | A. pleuropneumoniae | Diagnostic PCR for toxin typing of A. pleuropneumoniae strains [65] |
| apxIII | A. pleuropneumoniae | Diagnostic PCR for toxin typing of A. pleuropneumoniae strains [65] |
| apxIV | A. pleuropneumoniae | Diagnostic PCR detection of A.pleuropneumoniae [66] |
| aqxA | A. equli | Diagnostic PCR for identification of A. equuli subsp. haemolyticus [67] |
| AvxA | A. paragallinarium | Recombinant AvxA-RTX for development of vaccines against infectious coryza [68] |
| avxA | A. paragallinarium | Diagnostic PCR for A. paragallinarium species confirmation [68] |
| GtxA | G. anatis | Antigen in experimental vaccines against G. anatis infections in layer hens [69] |
| LktA | M. haemolytica | Antigen in commercial vaccines against mannheimiosis and B. threalosi infections [61,62] |
| lktD | M. haemolytica | Diagnostic multiple PCR for bovine respiratory disease complex (BRDC) [70] |
| lktA | M. haemolytica | Diagnostic multiple PCR for bovine respiratory disease complex (BRDC) [71] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frey, J. RTX Toxins of Animal Pathogens and Their Role as Antigens in Vaccines and Diagnostics. Toxins 2019, 11, 719. https://doi.org/10.3390/toxins11120719
Frey J. RTX Toxins of Animal Pathogens and Their Role as Antigens in Vaccines and Diagnostics. Toxins. 2019; 11(12):719. https://doi.org/10.3390/toxins11120719
Chicago/Turabian StyleFrey, Joachim. 2019. "RTX Toxins of Animal Pathogens and Their Role as Antigens in Vaccines and Diagnostics" Toxins 11, no. 12: 719. https://doi.org/10.3390/toxins11120719
APA StyleFrey, J. (2019). RTX Toxins of Animal Pathogens and Their Role as Antigens in Vaccines and Diagnostics. Toxins, 11(12), 719. https://doi.org/10.3390/toxins11120719