Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates Pro-Inflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin


Abstract
1. Introduction
2. Results
2.1. Monocultures and Co-Cultures Exhibit Differential Secretion of Interleukin (IL)-6, Interleukin (IL-8) and Tumor Necrosis Factor (TNF)-α under Basal Conditions
2.2. Stx2 and SubAB Induce Changes in the Release of Pro-Inflammatory Mediators by HGEC and HK-2 Monocultures
2.3. Stx2 and SubAB Exhibit Differential Effects on IL-6, IL-8 and TNF-α Secretion by Monocultures and Co-Cultures
2.4. Soluble Mediators Released by HGEC Increase the Secretion of IL-8 by HK-2
2.5. Toxins Were Incorporated by HGEC and HK-2 Cells Derived from Co-Cultures
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents
5.2. Antibodies
5.3. Human Primary Glomerular Microvascular Endothelial Cell Culture
5.4. Tubular Human Epithelial Cell Line Culture
5.5. Monocultures and Co-Cultures of Renal Endothelial and Epithelial Cells
5.6. Integrity of Endothelial and Epithelial Monolayers and Bilayers
5.7. Cytokine Detection by ELISA
5.8. Cell Viability Evaluation
5.9. Intracellular Detection of Stx2 and SubAB by Flow Cytometry in HGEC/HK-2 Co-Cultures
5.10. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karpman, D. Haemolytic uremic syndrome and thrombotic thrombocytopenic purpura. Curr. Paediatr. 2002, 12, 569–574. [Google Scholar] [CrossRef]
- Karmali, M.A.; Petric, M.; Lim, C.; Fleming, P.C.; Arbus, G.S.; Lior, H. The association between idiopathic hemolytic uremic syndrome and infection by verotoxin-producing escherichia coli. J. Infect. Dis. 1985, 151, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J.M.; Stapleton, J.; Angulo, F.J.; Yeung, D.H.; Kirk, M.D. Global incidence of human shiga toxin-producing escherichia coli infections and deaths: A systematic review and knowledge synthesis. Foodborne Pathog. Dis. 2014, 11, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Pianciola, L.; Rivas, M. Genotypic features of clinical and bovine escherichia coli o157 strains isolated in countries with different associated-disease incidences. Microorganisms 2018, 6, 36. [Google Scholar] [CrossRef]
- Torres, A.G.; Amaral, M.M.; Bentancor, L.; Galli, L.; Goldstein, J.; Kruger, A.; Rojas-Lopez, M. Recent advances in shiga toxin-producing escherichia coli research in latin america. Microorganisms 2018, 6, 100. [Google Scholar] [CrossRef]
- Rivas, M.; Chinen, I.; Miliwebsky, E.; Masana, M. Risk factors for shiga toxin-producing escherichia coli-associated human diseases. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- Argentina, SdPs MsdN. Boletín Integrado de Vigilancia; Secretaría de promoción y programas sanitarios. Ministerio de Salud de la Nación Argentina: Buenos Aires, Argentina, 2017; p. 352-SE. [Google Scholar]
- Repetto, H.A. Epidemic hemolytic-uremic syndrome in children. Kidney Int. 1997, 52, 1708–1719. [Google Scholar] [CrossRef]
- Repetto, H.A. Microangiopatía trombótica y sindrome hemolítico urémico. Nefrol. Clínica 3ra Edición 2009, 6, 286–297. [Google Scholar]
- Kruger, A.; Lucchesi, P.M. Shiga toxins and stx phages: Highly diverse entities. Microbiology 2015, 161, 451–462. [Google Scholar] [CrossRef]
- Ibarra, C.; Amaral, M.M.; Palermo, M.S. Advances in pathogenesis and therapy of hemolytic uremic syndrome caused by shiga toxin-2. Iubmb. Life 2013, 65, 827–835. [Google Scholar] [CrossRef]
- Golshani, M.; Oloomi, M.; Bouzari, S. In silico analysis of shiga toxins (stxs) to identify new potential vaccine targets for shiga toxin-producing escherichia coli. In Silico Pharmacol. 2016, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an escherichia coli strain producing a novel shiga toxin 2 subtype. Sci. Rep. 2018, 8, 6756. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.C.; Svanevik, C.S.; Lunestad, B.T.; Sekse, C.; Johannessen, G.S. Isolation and characterisation of shiga toxin-producing escherichia coli from norwegian bivalves. Food Microbiol. 2019, 84, 103268. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.; Miliwebsky, E.; Chinen, I.; Deza, N.; Leotta, G.A. The epidemiology of hemolytic uremic syndrome in argentina. Diagnosis of the etiologic agent, reservoirs and routes of transmission. Med. (B Aires) 2006, 66 (Suppl. 3), 27–32. [Google Scholar]
- Fitzgerald, S.F.; Beckett, A.E.; Palarea-Albaladejo, J.; McAteer, S.; Shaaban, S.; Morgan, J.; Ahmad, N.I.; Young, R.; Mabbott, N.A.; Morrison, L.; et al. Shiga toxin sub-type 2a increases the efficiency of escherichia coli o157 transmission between animals and restricts epithelial regeneration in bovine enteroids. PLoS Pathog. 2019, 15, e1008003. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S. Prevalence and pathogenicity of shiga toxin-producing escherichia coli in beef cattle and their products. J. Anim. Sci. 2007, 85, E63–E72. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.; Miliwebsky, E. Diagnóstico etiológico para establecer la asociación entre enfermedad humana e infección por Escherichia coli productor de toxina Shiga, cap 4. En Síndrome Urémico Hemolítico post-entérico. Actualización en patogénesis, diagnóstico y tratamiento del Síndrome Urémico Hemolítico asociado a la toxina Shiga. Editor. Acad. Esp. 2015, 4, 28–36. [Google Scholar]
- Lingwood, C.A. Verotoxin/globotriaosyl ceramide recognition: Angiopathy, angiogenesis and antineoplasia. Biosci. Rep. 1999, 19, 345–354. [Google Scholar] [CrossRef]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Tesh, V.L. Activation of cell stress response pathways by shiga toxins. Cell Microbiol. 2012, 14, 1–9. [Google Scholar] [CrossRef]
- Paton, A.W.; Srimanote, P.; Talbot, U.M.; Wang, H.; Paton, J.C. A new family of potent ab(5) cytotoxins produced by shiga toxigenic escherichia coli. J. Exp. Med. 2004, 200, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Galli, L.; Miliwebsky, E.; Irino, K.; Leotta, G.; Rivas, M. Virulence profile comparison between lee-negative shiga toxin-producing escherichia coli (stec) strains isolated from cattle and humans. Vet. Microbiol. 2010, 143, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Paton, A.W. Shiga toxin ‘goes retro’ in human primary kidney cells. Kidney Int. 2006, 70, 2049–2051. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, G.; Morinaga, N.; Yahiro, K.; Komine, R.; Moss, J.; Yoshida, H.; Noda, M. Novel subtilase cytotoxin produced by shiga-toxigenic escherichia coli induces apoptosis in vero cells via mitochondrial membrane damage. Infect. Immun. 2009, 77, 2919–2924. [Google Scholar] [CrossRef] [PubMed]
- May, K.L.; Paton, J.C.; Paton, A.W. Escherichia coli subtilase cytotoxin induces apoptosis regulated by host bcl-2 family proteins bax/bak. Infect. Immun. 2010, 78, 4691–4696. [Google Scholar] [CrossRef]
- Wolfson, J.J.; May, K.L.; Thorpe, C.M.; Jandhyala, D.M.; Paton, J.C.; Paton, A.W. Subtilase cytotoxin activates perk, ire1 and atf6 endoplasmic reticulum stress-signalling pathways. Cell. Microbiol. 2008, 10, 1775–1786. [Google Scholar] [CrossRef]
- Wang, H.; Paton, J.C.; Paton, A.W. Pathologic changes in mice induced by subtilase cytotoxin, a potent new escherichia coli ab5 toxin that targets the endoplasmic reticulum. J. Infect. Dis. 2007, 196, 1093–1101. [Google Scholar] [CrossRef]
- Byres, E.; Paton, A.W.; Paton, J.C.; Lofling, J.C.; Smith, D.F.; Wilce, M.C.; Talbot, U.M.; Chong, D.C.; Yu, H.; Huang, S.; et al. Incorporation of a non-human glycan mediates human susceptibility to a bacterial toxin. Nature 2008, 456, 648–652. [Google Scholar] [CrossRef]
- Inoue, S.; Sato, C.; Kitajima, K. Extensive enrichment of n-glycolylneuraminic acid in extracellular sialoglycoproteins abundantly synthesized and secreted by human cancer cells. Glycobiology 2010, 20, 752–762. [Google Scholar] [CrossRef]
- Lofling, J.C.; Paton, A.W.; Varki, N.M.; Paton, J.C.; Varki, A. A dietary non-human sialic acid may facilitate hemolytic-uremic syndrome. Kidney Int. 2009, 76, 140–144. [Google Scholar] [CrossRef]
- Velandia, C.V.; Mariel Sanso, A.; Kruger, A.; Suarez, L.V.; Lucchesi, P.M.; Parma, A.E. Occurrence of subtilase cytotoxin and relation with other virulence factors in verocytotoxigenic escherichia coli isolated from food and cattle in argentina. Braz. J. Microbiol. 2011, 42, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Obrig, T.G. Escherichia coli shiga toxin mechanisms of action in renal disease. Toxins 2010, 2, 2769–2794. [Google Scholar] [CrossRef] [PubMed]
- Exeni, R.A.; Fernandez-Brando, R.J.; Santiago, A.P.; Fiorentino, G.A.; Exeni, A.M.; Ramos, M.V.; Palermo, M.S. Pathogenic role of inflammatory response during Shiga toxin-associated Hemolytic Uremic Syndrome (HUS). Pediatr. Nephrol. 2018, 33, 2057–2071. [Google Scholar] [CrossRef] [PubMed]
- Repetto, H.A.; Rodríguez de Córdoba, S.; Arrizurieta, E.; Rivas, M.; Ibarra, C. Microangiopatía Trombótica y Síndrome Urémico Hemolítico. In Nefrología Clínica; Editorial médica panamericana; Ministerio de Salud de la Nación Argentina: Buenos Aires, Argentina, 2014; Cap 25 (351-363). [Google Scholar]
- Richardson, S.E.; Karmali, M.A.; Becker, L.E.; Smith, C.R. The histopathology of the hemolytic uremic syndrome associated with verocytotoxin-producing escherichia coli infections. Hum. Pathol. 1988, 19, 1102–1108. [Google Scholar] [CrossRef]
- Kaplan, B.S. Shiga toxin-induced tubular injury in hemolytic uremic syndrome. Kidney Int. 1998, 54, 648–649. [Google Scholar] [CrossRef]
- Amaral, M.M.; Sacerdoti, F.; Jancic, C.; Repetto, H.A.; Paton, A.W.; Paton, J.C.; Ibarra, C. Action of shiga toxin type-2 and subtilase cytotoxin on human microvascular endothelial cells. PLoS ONE 2013, 8, e70431. [Google Scholar] [CrossRef]
- Alvarez, R.S.; Sacerdoti, F.; Jancic, C.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Amaral, M.M. Comparative characterization of shiga toxin type 2 and subtilase cytotoxin effects on human renal epithelial and endothelial cells grown in monolayer and bilayer conditions. PLoS ONE 2016, 11, e0158180. [Google Scholar] [CrossRef]
- Valtin, H. Renal function: Mechanisms preserving fluid and solute balance in health and disease. In Renal Hemodynamics and Oxigen Consumption; Little, Brown and Company: Boston, MA, USA, 1973; Chapter 6. [Google Scholar]
- Amigo, N.; Mercado, E.; Bentancor, A.; Singh, P.; Vilte, D.; Gerhardt, E.; Zotta, E.; Ibarra, C.; Manning, S.D.; Larzabal, M.; et al. Clade 8 and clade 6 strains of escherichia coli o157:H7 from cattle in argentina have hypervirulent-like phenotypes. PLoS ONE 2015, 10, e0127710. [Google Scholar] [CrossRef]
- Pianciola, L.; D’Astek, B.A.; Mazzeo, M.; Chinen, I.; Masana, M.; Rivas, M. Genetic features of human and bovine escherichia coli o157:H7 strains isolated in argentina. Int. J. Med. Microbiol. 2016, 306, 123–130. [Google Scholar] [CrossRef]
- Aydin, S.; Signorelli, S.; Lechleitner, T.; Joannidis, M.; Pleban, C.; Perco, P.; Pfaller, W.; Jennings, P. Influence of microvascular endothelial cells on transcriptional regulation of proximal tubular epithelial cells. Am. J. Physiol. Cell Physiol. 2008, 294, C543–C554. [Google Scholar] [CrossRef]
- Bijuklic, K.; Jennings, P.; Kountchev, J.; Hasslacher, J.; Aydin, S.; Sturn, D.; Pfaller, W.; Patsch, J.R.; Joannidis, M. Migration of leukocytes across an endothelium-epithelium bilayer as a model of renal interstitial inflammation. Am. J. Physiol. Cell Physiol. 2007, 293, C486–C492. [Google Scholar] [CrossRef] [PubMed]
- Guessous, F.; Marcinkiewicz, M.; Polanowska-Grabowska, R.; Kongkhum, S.; Heatherly, D.; Obrig, T.; Gear, A.R. Shiga toxin 2 and lipopolysaccharide induce human microvascular endothelial cells to release chemokines and factors that stimulate platelet function. Infect. Immun. 2005, 73, 8306–8316. [Google Scholar] [CrossRef] [PubMed]
- Zoja, C.; Angioletti, S.; Donadelli, R.; Zanchi, C.; Tomasoni, S.; Binda, E.; Imberti, B.; te Loo, M.; Monnens, L.; Remuzzi, G.; et al. Shiga toxin-2 triggers endothelial leukocyte adhesion and transmigration via nf-kappab dependent up-regulation of il-8 and mcp-1. Kidney Int. 2002, 62, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Matussek, A.; Lauber, J.; Bergau, A.; Hansen, W.; Rohde, M.; Dittmar, K.E.; Gunzer, M.; Mengel, M.; Gatzlaff, P.; Hartmann, M.; et al. Molecular and functional analysis of shiga toxin-induced response patterns in human vascular endothelial cells. Blood 2003, 102, 1323–1332. [Google Scholar] [CrossRef]
- Brigotti, M.; Carnicelli, D.; Ravanelli, E.; Vara, A.G.; Martinelli, C.; Alfieri, R.R.; Petronini, P.G.; Sestili, P. Molecular damage and induction of proinflammatory cytokines in human endothelial cells exposed to shiga toxin 1, shiga toxin 2, and alpha-sarcin. Infect. Immun. 2007, 75, 2201–2207. [Google Scholar] [CrossRef]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin-associated hemolytic uremic syndrome: Pathophysiology of endothelial dysfunction. Pediatr. Nephrol. 2010, 25, 2231–2240. [Google Scholar] [CrossRef]
- Wang, H.; Rogers, T.J.; Paton, J.C.; Paton, A.W. Differential effects of escherichia coli subtilase cytotoxin and shiga toxin 2 on chemokine and proinflammatory cytokine expression in human macrophage, colonic epithelial, and brain microvascular endothelial cell lines. Infect. Immun. 2014, 82, 3567–3579. [Google Scholar] [CrossRef]
- Lentz, E.K.; Leyva-Illades, D.; Lee, M.S.; Cherla, R.P.; Tesh, V.L. Differential response of the human renal proximal tubular epithelial cell line hk-2 to shiga toxin types 1 and 2. Infect. Immun. 2011, 79, 3527–3540. [Google Scholar] [CrossRef]
- Galley, H.F.; Webster, N.R. Physiology of the endothelium. Br. J. Anaesth. 2004, 93, 105–113. [Google Scholar] [CrossRef]
- Tasnim, F.; Zink, D. Cross talk between primary human renal tubular cells and endothelial cells in cocultures. Am. J. Physiol. Ren. Physiol. 2012, 302, F1055–F1062. [Google Scholar] [CrossRef]
- Nee, L.E.; McMorrow, T.; Campbell, E.; Slattery, C.; Ryan, M.P. Tnf-alpha and il-1beta-mediated regulation of mmp-9 and timp-1 in renal proximal tubular cells. Kidney Int. 2004, 66, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Angelini, D.J.; Hyun, S.W.; Grigoryev, D.N.; Garg, P.; Gong, P.; Singh, I.S.; Passaniti, A.; Hasday, J.D.; Goldblum, S.E. Tnf-alpha increases tyrosine phosphorylation of vascular endothelial cadherin and opens the paracellular pathway through fyn activation in human lung endothelia. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L1232–L1245. [Google Scholar] [CrossRef] [PubMed]
- Goldblum, S.E.; Ding, X.; Campbell-Washington, J. Tnf-alpha induces endothelial cell f-actin depolymerization, new actin synthesis, and barrier dysfunction. Am. J. Physiol. 1993, 264, C894–C905. [Google Scholar] [CrossRef] [PubMed]
- Beddoe, T.; Paton, A.W.; Le Nours, J.; Rossjohn, J.; Paton, J.C. Structure, biological functions and applications of the ab5 toxins. Trends Biochem. Sci. 2010, 35, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Luz, D.E.; Moraes, C.T.; Caravelli, A.; Fernandes, I.; Guth, B.E.; Horton, D.S.; Piazza, R.M. Interaction between shiga toxin and monoclonal antibodies: Binding characteristics and in vitro neutralizing abilities. Toxins 2012, 4, 729–747. [Google Scholar] [CrossRef]
- Wang, H.; Paton, A.W.; McColl, S.R.; Paton, J.C. In vivo leukocyte changes induced by escherichia coli subtilase cytotoxin. Infect. Immun. 2011, 79, 1671–1679. [Google Scholar] [CrossRef]
- Creydt, V.P.; Silberstein, C.; Zotta, E.; Ibarra, C. Cytotoxic effect of shiga toxin-2 holotoxin and its b subunit on human renal tubular epithelial cells. Microbes Infect. 2006, 8, 410–419. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
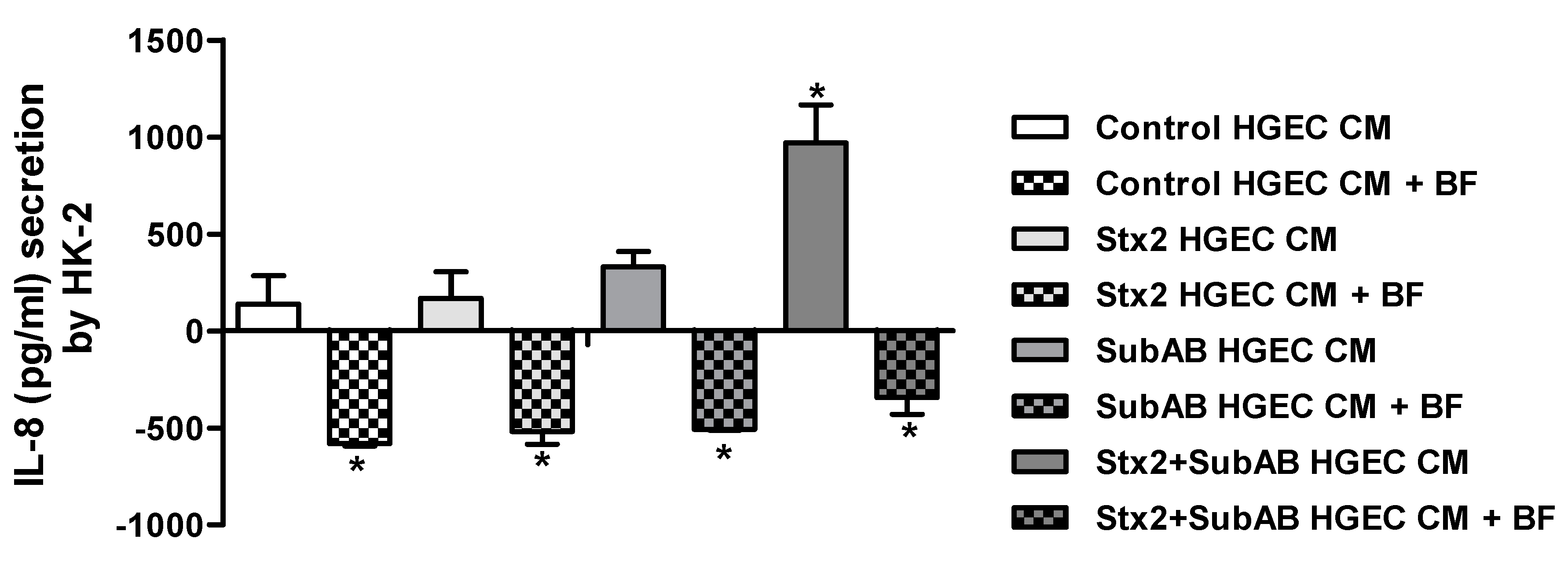{kind=link}
{kind=link}
| HGEC | Control | Stx2 | SubAB | Stx2+SubAB | ||||
|---|---|---|---|---|---|---|---|---|
| % | SEM | % | SEM | % | SEM | % | SEM | |
| Stx2 + | 1.5 | 1.2 | 48.4 * | 8.5 | 0.7 | 0.4 | 0.9 | 0.4 |
| SubAB + | 0.2 | 0.1 | 0.9 | 1.3 | 62.6 * | 7.5 | 20.3 * | 4.1 |
| Stx2 – SubAB + | 0.3 | 0.1 | 0.3 | 0.2 | 0.7 | 0.6 | 68.3 * | 4.2 |
| HK-2 | Control | Stx2 | SubAB | Stx2+SubAB | ||||
|---|---|---|---|---|---|---|---|---|
| % | SEM | % | SEM | % | SEM | % | SEM | |
| Stx2 + | 1.8 | 1.1 | 35.0 * | 4.1 | 1.9 | 0.9 | 4.9 | 6.6 |
| SubAB + | 0.3 | 0.1 | 3.8 | 4.4 | 31.8 * | 5.9 | 12.5 * | 2.0 |
| Stx2 – SubAB + | 0.4 | 0.1 | 0.8 | 0.4 | 0.9 | 0.4 | 40.6 * | 15.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez, R.S.; Jancic, C.; Garimano, N.; Sacerdoti, F.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Amaral, M.M. Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates Pro-Inflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin. Toxins 2019, 11, 648. https://doi.org/10.3390/toxins11110648
Álvarez RS, Jancic C, Garimano N, Sacerdoti F, Paton AW, Paton JC, Ibarra C, Amaral MM. Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates Pro-Inflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin. Toxins. 2019; 11(11):648. https://doi.org/10.3390/toxins11110648
Chicago/Turabian StyleÁlvarez, Romina S., Carolina Jancic, Nicolás Garimano, Flavia Sacerdoti, Adrienne W. Paton, James C. Paton, Cristina Ibarra, and María M. Amaral. 2019. "Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates Pro-Inflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin" Toxins 11, no. 11: 648. https://doi.org/10.3390/toxins11110648
APA StyleÁlvarez, R. S., Jancic, C., Garimano, N., Sacerdoti, F., Paton, A. W., Paton, J. C., Ibarra, C., & Amaral, M. M. (2019). Crosstalk between Human Microvascular Endothelial Cells and Tubular Epithelial Cells Modulates Pro-Inflammatory Responses Induced by Shiga Toxin Type 2 and Subtilase Cytotoxin. Toxins, 11(11), 648. https://doi.org/10.3390/toxins11110648