Development of Indirect Competitive Enzyme-Linked Immunosorbent Assay to Detect Fusarium verticillioides in Poultry Feed Samples

 ,
,
Abstract

1. Introduction
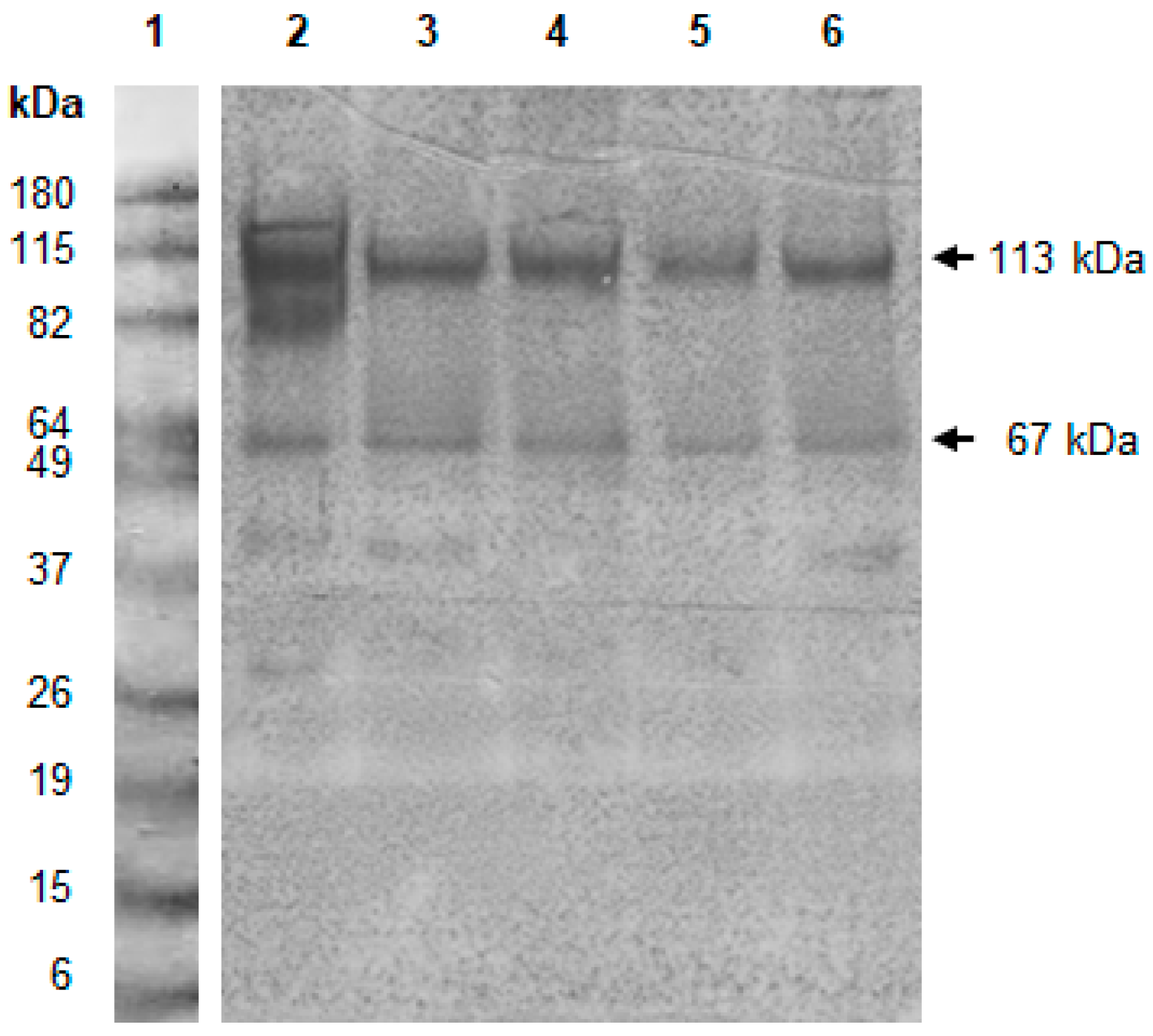
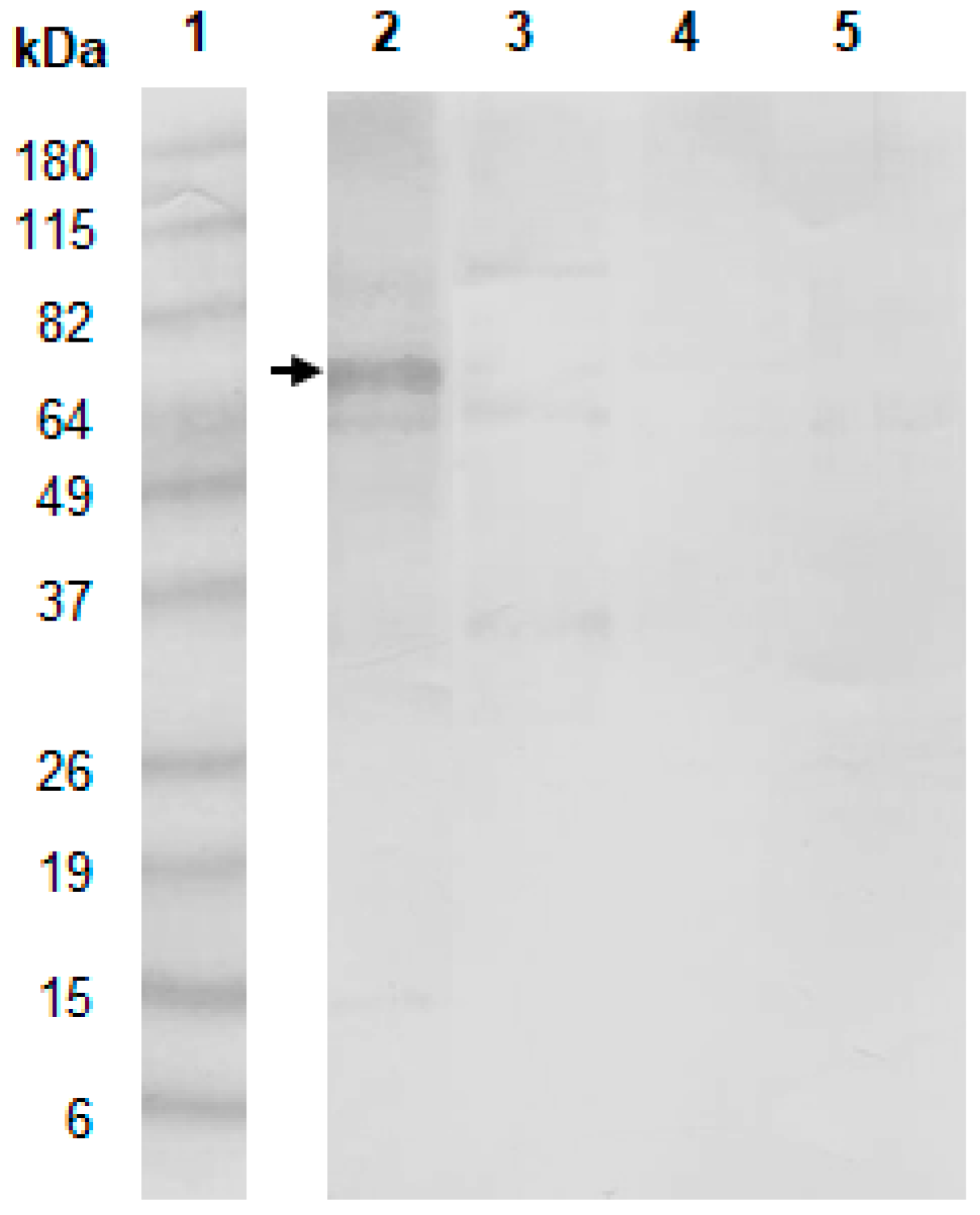
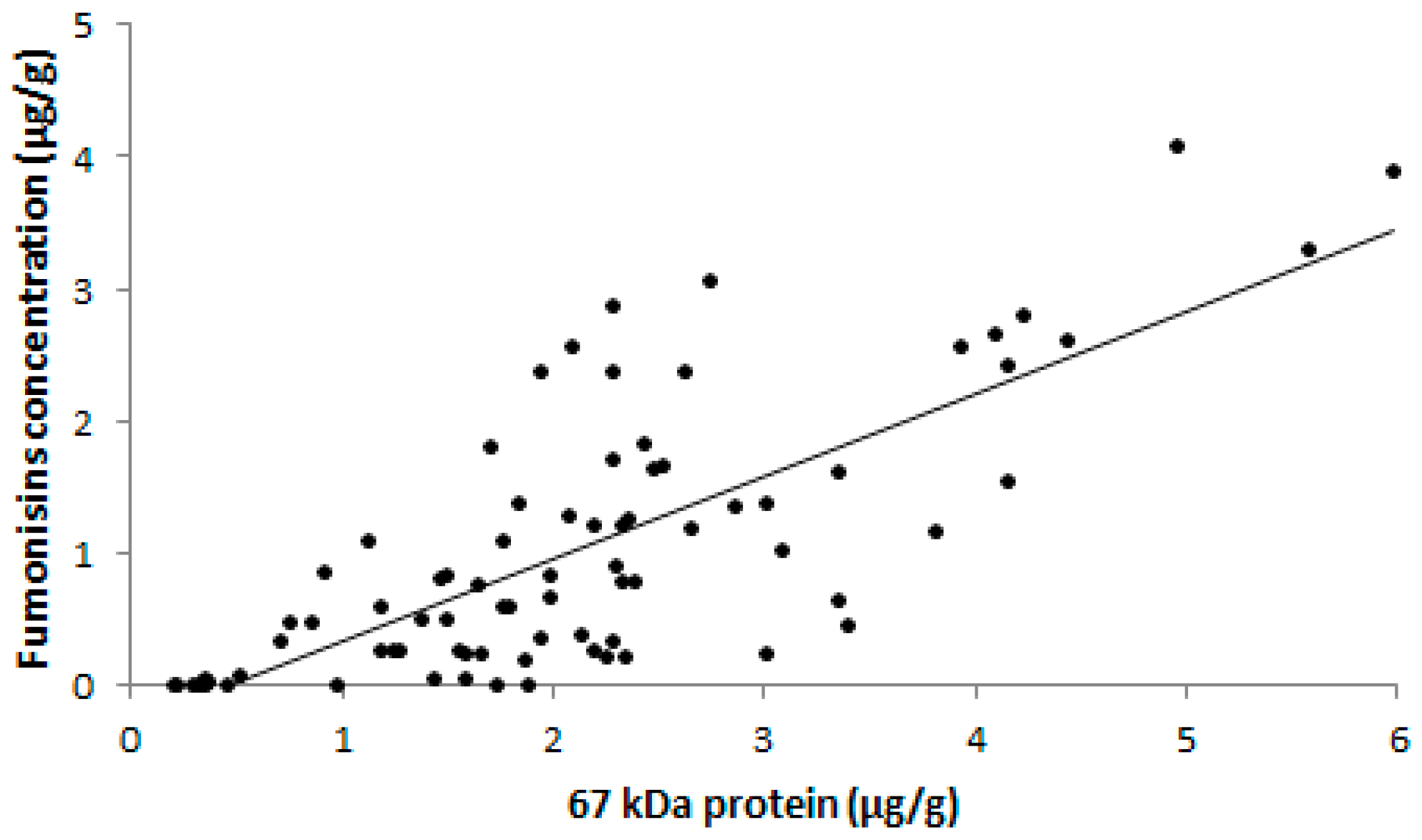
2. Results and discussion
3. Material and Methods
3.1. Fungal Isolates
3.2. Exoantigen Preparation
3.3. Antibodies Production
3.4. Indirect ELISA
3.5. Western Blot
3.6. Purification of 67 kDa Orotein by Affinity Chromatography
3.7. Poultry Feed Samples
3.8. Fusarium sp. Count and Exoantigen Extraction from Feed Samples
3.9. Indirect Competitive ELISA
3.10. Fumonisin Analysis
3.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CONAB—Acompanhamento da Safra Brasileira de Grãos, v. 5—Safra 2017/18, n4—Quarto Levantamento, Janeiro 2018. Available online: http://www.conab.gov.br/OlalaCMS/uploads/arquivos/18_01_11_14_17_49_graos_4o_levantamento.pdf (accessed on 12 January 2018).
- ABIMILHO—Associação Brasileira das Indústrias de Milho. Estatísticas. 2017. Available online: http://www.abimilho.com.br/estatisticas (accessed on 20 December 2017).
- Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Butkeraitis, P.; Oliveira, C.A.F.; Ledoux, D.R.; Ogido, R.; Albuquerque, R.; Rosmaninho, J.F.; Rottinghaus, G.E. Effect of dietary fumonisin B1 on laying Japanese quail. Br. Poult. Sci. 2004, 45, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poultry Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Zhu, W.; Pang, M.; Liu, Y.; Dong, J. Natural occurrence of fumonisins B1 and B2 in corn in four provinces of China. Food Addit. Contam. Part B 2013, 6, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.N.; Takabayashi, C.R.; Ono, M.A.; Bordini, J.G.; Kawamura, O.; Vizoni, E.; Hirooka, E.Y.; Ono, E.Y.S. Assessment of exposure of broiler chicken in Brazil to mycotoxins through naturally contaminated feed. Food Secur. 2013, 5, 541–550. [Google Scholar] [CrossRef]
- Monge, M.P.; Magnoli, C.E.; Chiacchiera, S.M. Survey of Aspergillus and Fusarium species and their mycotoxins in raw materials and poultry feeds from Córdoba, Argentina. Mycotoxin Res. 2012, 28, 111–122. [Google Scholar] [CrossRef]
- Greco, M.V.; Franchi, M.L.; Golba, S.L.R.; Pardo, A.G.; Pose, G.N. Mycotoxins and mycotoxigenic fungi in poultry feed for food-producing animals. Sci. World J. 2014, 9. [Google Scholar] [CrossRef]
- Bordini, J.G.; Ono, M.A.; Garcia, G.T.; Fazani, V.H.F.; Vizoni, K.; Rodrigues, K.C.B.; Hirooka, E.Y.; Ono, E.Y.S. Impact of industrial dry-milling on fumonisin redistribution in non-transgenic corn in Brazil. Food Chem. 2017, 220, 438–443. [Google Scholar] [CrossRef]
- Yeni, F.; Acar, S.; Polat, O.G.; Soyer, Y.; Alpas, H. Rapid and standardized methods for detection of foodborne pathogens and mycotoxins on fresh produce. Food Control 2014, 40, 359–367. [Google Scholar] [CrossRef]
- Bluhm, B.H.; Flaherty, J.E.; Cousin, M.A.; Woloshuk, C.P. Multiplex polymerase chain reaction assay for the differential detection of trichothecene- and fumonisin-producing species of Fusarium in cornmeal. J. Food Prot. 2002, 65, 1955–1961. [Google Scholar] [CrossRef] [PubMed]
- Dawidziuk, A.; Koczyk, G.; Popiel, D.; Kaczmarek, J.; Busko, M. Molecular diagnostics on the toxigenic potential of Fusarium spp. plant pathogens. J. Appl. Microbiol. 2014, 116, 1607–1620. [Google Scholar] [CrossRef] [PubMed]
- Ramana, M.V.; Balakrishna, K.; Murali, H.C.S.; Batra, H.V. Multiplex PCR-based strategy to detect contamination with mycotoxigenic Fusarium species in rice and fingermillet collected from southern India. J. Sci. Food Agric. 2011, 91, 1666–1673. [Google Scholar] [CrossRef] [PubMed]
- Rashmi, R.; Ramana, M.V.; Shylaja, R.; Uppalapati, S.R.; Murali, H.S.; Batra, H.V. Evaluation of multiplex PCR assay for concurrent detection of four major mycotoxigenic fungi from foods. J. Appl. Microbiol. 2012, 144, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasa, M.Y.; Dass, R.S.; Raj, A.P.C.; Janardhana, G.R. Molecular detection of fumonisin producing Fusarium species of freshly harvested maize kernels using Polymerase Chain Reaction (PCR). Taiwania 2006, 51, 251–257. [Google Scholar]
- Omori, A.M.; Ono, E.Y.S.; Bordini, J.G.; Hirozawa, M.T.; Fungaro, M.H.P.; Ono, M.A. Detection of Fusarium verticillioides by PCR-ELISA based on FUM21 gene. Food Microbiol. 2018, 73, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, L.; Standard, P.G. Specific and rapid identification of medically important fungi by exoantigen detection. Ann. Rev. Microbiol. 1987, 41, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Iyer, M.S.; Cousin, M.A. Immunological detection of Fusarium species in cornmeal. J. Food Prot. 2003, 66, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Shon, D.; Kim, Y. Application of an enzyme-linked immunosorbent assay for detecting mold contamination in agricultural commodities and comparison with conventional assays. Food Agric. Immunol. 2003, 15, 159–166. [Google Scholar] [CrossRef]
- Meirelles, P.G.; Ono, M.A.; Ohe, M.C.T.; Maroneze, D.M.; Itano, E.N.; Garcia, G.T.; Sugiura, Y.; Ueno, Y.; Hirooka, E.Y.; Ono, E.Y.S. Detection of Fusarium sp. contamination in corn by enzyme-linked immunosorbent assay. Food Agric. Immunol. 2006, 17, 79–89. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A.; Bullock, S. The Fusarium Laboratory Manual; Wiley-Blackwell Publishing: Ames, IA, USA, 2008; p. 388. [Google Scholar]
- Ono, E.Y.S.; Sugiura, Y.; Homechin, M.; Kamogae, M.; Vizzoni, E.; Ueno, Y.; Hirooka, E.Y. Effect of climatic conditions on natural mycoflora and fumonisins in freshly harvested corn of the State of Paraná, Brazil. Mycopathologia 1999, 147, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Rahjoo, V.; Zad, J.; Javan-Nikkhah, M.; Gohari, A.M.; Okhovvat, S.M.; Bihanta, M.R.; Razzaghian, J.; Klemsdal, S.S. Morphological and molecular identification of Fusarium isolated from maize ears in Iran. J. Plant Pathol. 2008, 90, 463–468. [Google Scholar]
- Rocha, L.O.; Reis, G.M.; Silva, V.N.; Braghini, R.; Teixeira, M.M.G.; Corrêa, B. Molecular characterization and fumonisin production by Fusarium verticillioides isolated from corn grains of different geographic origins in Brazil. Int. J. Food Microbiol. 2011, 145, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Madania, A.; Altawil, M.; Naffaa, W.; Volker, P.H.; Hawat, M. Morphological and molecular characterization of Fusarium isolated from maize in Syria. J. Phytopathol. 2013, 161, 452–458. [Google Scholar] [CrossRef]
- Lanza, F.E.; Zambolim, L.; Costa, R.V.; Queiroz, V.A.V.; Cota, L.V.; Silva, D.D.; Souza, A.G.C.; Figueiredo, J.E.F. Prevalence of fumonisin-producing Fusarium species in Brazilian corn grains. Crop Prot. 2014, 65, 232–237. [Google Scholar] [CrossRef]
- Frisvad, J.C. Rationale for a polyphasic approach in the identification of mycotoxigenic fungi. In Determining Mycotoxins and Mycotoxigenic Fungi in Food and Feed; De Saeger, S., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2011; pp. 279–297. [Google Scholar]
- Kvas, M.; Marasas, W.F.O.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Diversity and evolution of Fusarium species in the Gibberella fujikuroi complex. Fungal Divers. 2009, 34, 1–21. [Google Scholar]
- Silva, J.J.; Viaro, H.P.; Ferranti, L.S.; Oliveira, A.L.M.; Ferreira, J.M.; Ruas, C.F.; Ono, E.Y.S.; Fungaro, M.H.P. Genetic structure of Fusarium verticillioides populations and occurrence of fumonisins in maize grown in Southern Brazil. Crop Prot. 2017, 99, 160–167. [Google Scholar] [CrossRef]
- Li, S.; Marquardt, R.R.; Abramson, D. Immunochemical detection of molds: A review. J. Food Prot. 2000, 63, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Yong, R.K.; Cousin, M.A. Detection of moulds producing aflatoxins in maize and peanuts by an immunoassay. Int. J. Food Microbiol. 2001, 65, 27–38. [Google Scholar] [CrossRef]
- Anand, S.; Rati, E.R. An enzyme linked immunosorbent assay for monitoring of Aspergillus ochraceus growth in coffee powder, chilli powder and poultry feed. Lett. Appl. Microbiol. 2006, 42, 59–65. [Google Scholar] [CrossRef]
- Biazon, L.; Meirelles, P.G.; Ono, M.A.; Itano, E.N.; Taniwaki, M.H.; Sugiura, Y.; Ueno, Y.; Hirooka, E.Y.; Ono, E.Y.S. Development of polyclonal antibodies against Fusarium verticillioides exoantigens. Food Agric. Immunol. 2006, 17, 69–77. [Google Scholar] [CrossRef]
- Gan, Z.; Marquardt, R.R.; Abramson, D.; Randy, M.C. The characterization of chicken antibodies raised against Fusarium spp. by enzyme-linked immunosorbent assay and immunoblotting. Int. J. Food Microbiol. 1997, 38, 191–200. [Google Scholar] [CrossRef]
- Divakara, S.T.; Santosh, P.; Aiyaz, M.; Ramana, M.V.; Hariprasad, P.; Nayaka, S.C.; Niranjana, S.R. Molecular identification and characterization of Fusarium spp. associated with sorghum seeds. J. Sci. Food Agric. 2014, 94, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). Background Paper in Support of Fumonisin Levels in Animal Feed: Executive Summary of This Scientific Support Document—Guidance for Industry: Fumonisin Levels in Human Foods and Animal Feeds. 2001. Available online: www.fda.gov/Food/FoodSafety/FoodContaminantsAdulteration/NaturalToxins (accessed on 10 December 2018).
- Labuda, R.; Parich, A.; Vekiru, E.; Tancinová, D. Incidence of fumonisins, moniliformin and Fusarium species in poultry feed mixtures from Slovakia. Ann. Agric. Environ. Med. 2005, 12, 81–86. [Google Scholar] [PubMed]
- Gourama, H.; Bullerman, L.B. Detection of molds in foods and feeds: Potential rapid and selective methods. J. Food Prot. 1995, 58, 1389–1394. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schwarzkopf, C.; Thiele, B. Effectivity of different methods for the extraction and purification of IgY. Altex 1996, 13, 35–39. [Google Scholar]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the Biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Nelson, P.E.; Tousson, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Rossi, C.N.; Takabayashi, C.R.; Ono, M.A.; Saito, G.H.; Itano, E.N.; Kawamura, O.; Hirooka, E.Y.; Ono, E.Y.S. Immunoassay based on monoclonal antibody for aflatoxin detection in poultry feed. Food Chem. 2012, 132, 2211–2216. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Exoantigen | Cross-Reactivity (%) | |
|---|---|---|
| Anti-Exoantigen | Anti-67 kDa Protein | |
| F. subglutinans 332 | 19 | 0 |
| F. subglutinans 852 | 17 | 0 |
| F. proliferatum 559 | 26 | 0 |
| F. sporotrichioides | 81 | 10 |
| F. graminearum FRS26 | 4 | 2 |
| F. graminearum FSP27 | 8 | 0 |
| F. graminearum 17102918 | 27 | 7 |
| A. niger 10A | 0 | 0 |
| A. niger 104CF | 0 | 0 |
| A. niger 4138 | 0 | 0 |
| A. niger 23115 | 0 | 0 |
| A. ochraceus 153 | 6 | 0 |
| A. ochraceus 4363 | 0 | 0 |
| A. ochraceus 4368 | 0 | 0 |
| A. flavus 58A | 0 | 0 |
| A. flavus 89A | 0 | 0 |
| A. carbonarius 178 | 3 | 0 |
| A. carbonarius 180 | 0 | 0 |
| A. carbonarius 222 | 10 | 0 |
| A. welwitschiae 112581 | 0 | 0 |
| A. welwitschiae 115625 | 0 | 0 |
| P. purpurogenum 30 F45 | 2 | 1 |
| P. variabile 30 F 33-4 | 7 | 0 |
| P. funiculosum 30 F 88-2 | 10 | 0 |
| P. brevicompactum | 13 | 0 |
| Parameters | Range | Mean | Median |
|---|---|---|---|
| Fusarium sp. count (CFU/g) | 50–7.5 × 105 | 6.2 × 104 | 6.0 × 103 |
| 67 kDa protein concentration (µg/g) | 2.0–59.8 | 21.0 | 19.8 |
| Fumonisin concentration (µg/g) | |||
| FB1 | 0.03–3.03 | 0.69 | 0.64 |
| FB2 | 0.03–1.27 | 0.33 | 0.30 |
| Total (FB1+ FB2) | 0.03–4.07 | 1.02 | 0.83 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omori, A.M.; Ono, E.Y.S.; Hirozawa, M.T.; de Souza Suguiura, I.M.; Hirooka, E.Y.; Pelegrinelli Fungaro, M.H.; Ono, M.A. Development of Indirect Competitive Enzyme-Linked Immunosorbent Assay to Detect Fusarium verticillioides in Poultry Feed Samples. Toxins 2019, 11, 48. https://doi.org/10.3390/toxins11010048
Omori AM, Ono EYS, Hirozawa MT, de Souza Suguiura IM, Hirooka EY, Pelegrinelli Fungaro MH, Ono MA. Development of Indirect Competitive Enzyme-Linked Immunosorbent Assay to Detect Fusarium verticillioides in Poultry Feed Samples. Toxins. 2019; 11(1):48. https://doi.org/10.3390/toxins11010048
Chicago/Turabian StyleOmori, Aline Myuki, Elisabete Yurie Sataque Ono, Melissa Tiemi Hirozawa, Igor Massahiro de Souza Suguiura, Elisa Yoko Hirooka, Maria Helena Pelegrinelli Fungaro, and Mario Augusto Ono. 2019. "Development of Indirect Competitive Enzyme-Linked Immunosorbent Assay to Detect Fusarium verticillioides in Poultry Feed Samples" Toxins 11, no. 1: 48. https://doi.org/10.3390/toxins11010048
APA StyleOmori, A. M., Ono, E. Y. S., Hirozawa, M. T., de Souza Suguiura, I. M., Hirooka, E. Y., Pelegrinelli Fungaro, M. H., & Ono, M. A. (2019). Development of Indirect Competitive Enzyme-Linked Immunosorbent Assay to Detect Fusarium verticillioides in Poultry Feed Samples. Toxins, 11(1), 48. https://doi.org/10.3390/toxins11010048