Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists

 ,
, 
 and
and 
Abstract

1. Introduction
2. Results
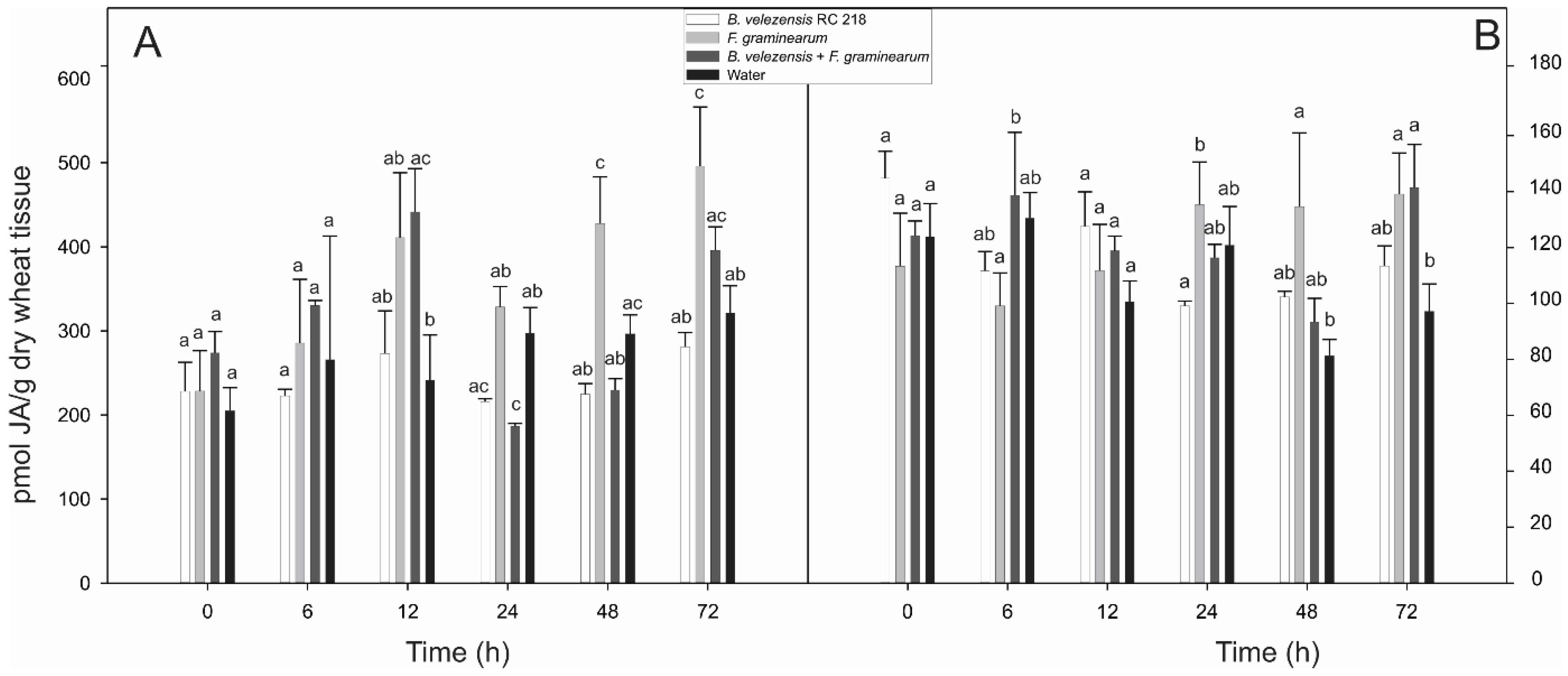
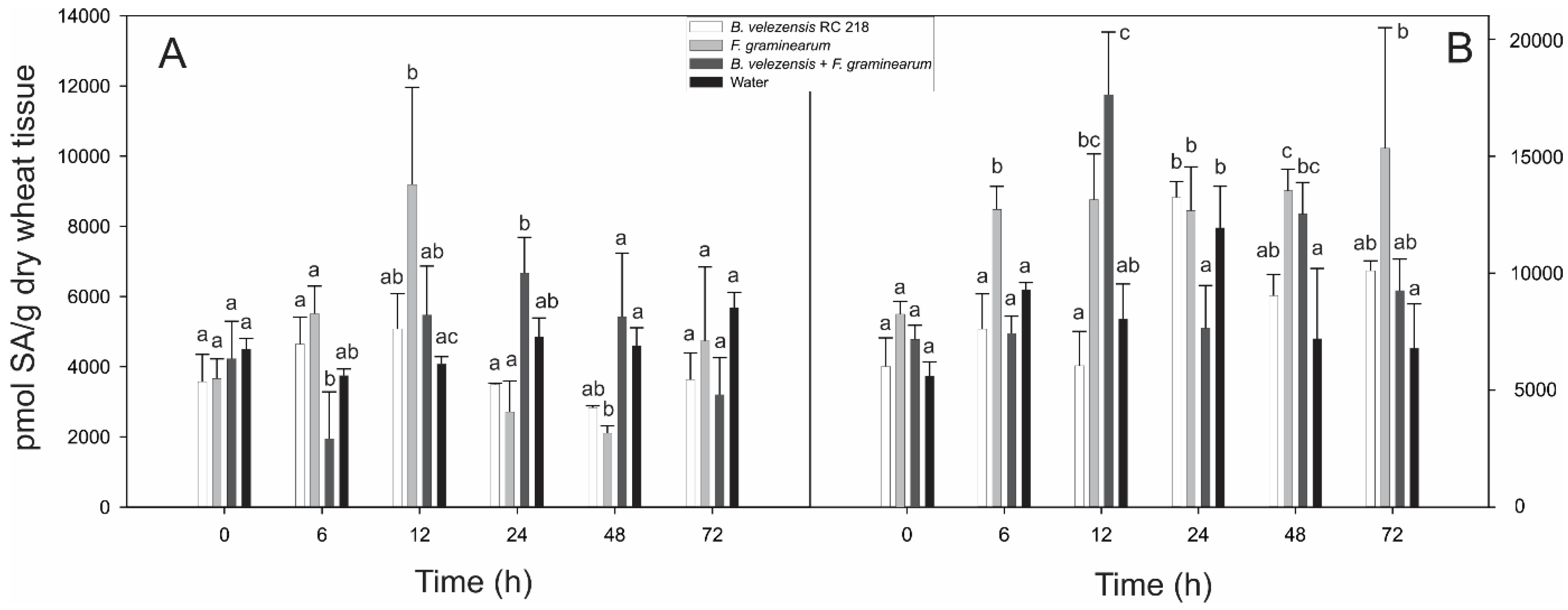
2.1. Jasmonic and Salicylic Acid Levels in Wheat Spikes Inoculated with Bacillus Velezensis RC 218 and Fusarium graminearum RC 276
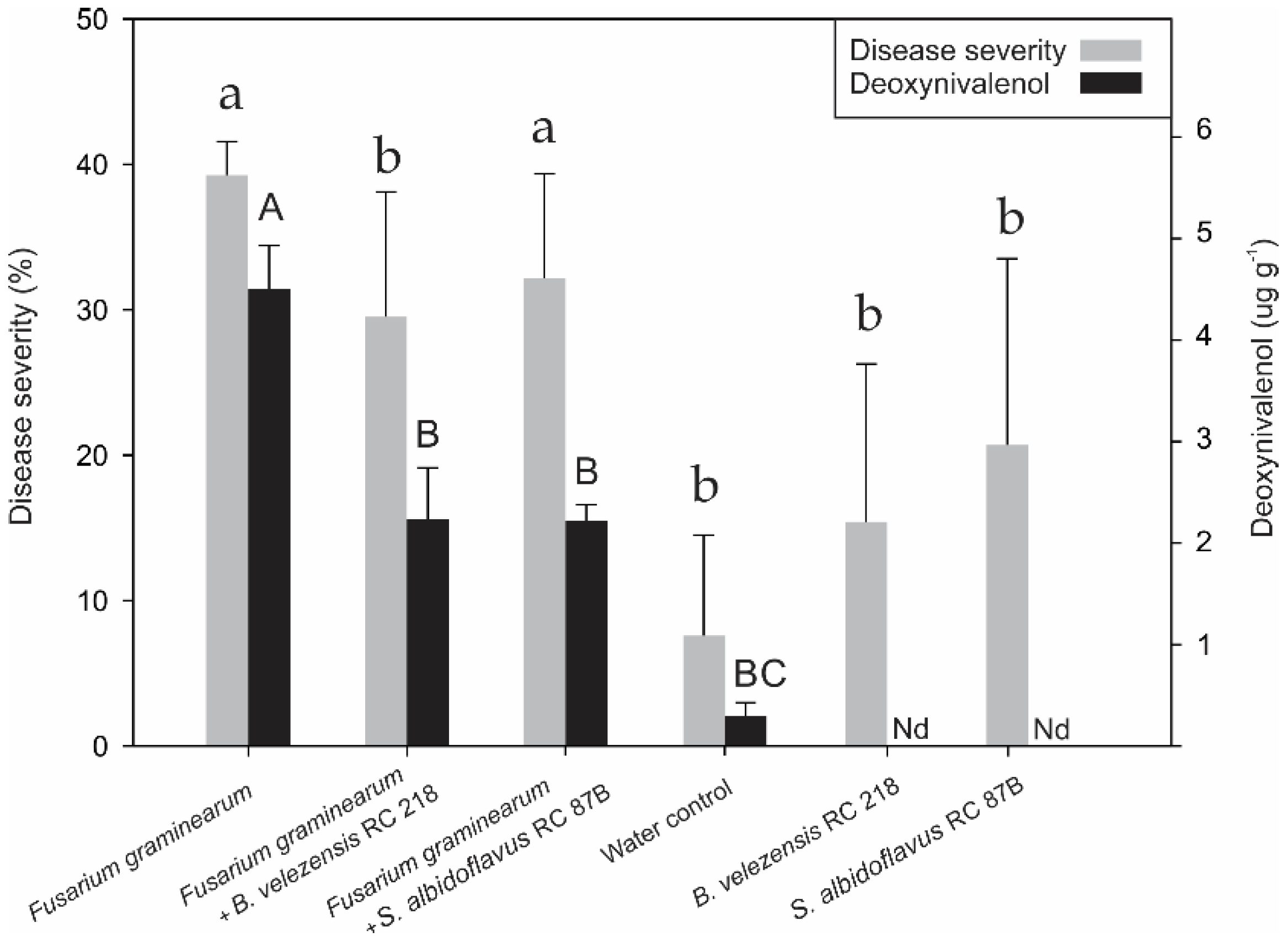
2.2. Biocontrol of Fusarium graminearum by B. Velezensis RC 218 and S. Albidoflavus RC 87B on Durum Wheat under Field Trial
3. Discussion
4. Materials and Methods
4.1. Salicylic and Jasmonic Acid in Wheat Spikes Inoculated with B. Velezensis RC 218 and F. Graminearum under Greenhouse Conditions
4.1.1. Greenhouse Conditions and Treatments Application
4.1.2. Phytohormone Extraction and Quantification by High Performance Liquid Chromatography-Tandem Mass Spectrometry
4.2. Evaluation of Bacillus Velezensis RC 218 and Streptomyces Albidoflavus RC 87B as Biocontrol Agents against F. Graminearum in Durum Wheat under Field Conditions
4.2.1. Field Trial on Durum Wheat
4.2.2. Culture Conditions of BCAs and Pathogen Production
4.2.3. Biocontrol Agents and Pathogen Applications
4.2.4. Fusarium Head Blight Incidence and Severity, DON Content, F. graminearum DNA Levels by qPCR and Fusarium-Damaged Kernels on Harvested Grains
4.2.5. Statistical Analyses of Phytohormone Values and FHB Parameters Evaluated
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and deoxynivalenol during the 2012/13 wheat Fusarium head blight outbreak in Argentina. Cereal Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef]
- Palacios, S.; Erazo, J.; Ciasca, B.; Lattanzio, V.; Reynoso, M.; Farnochi, C.; Torres, A. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat from Argentina. Food Chem. 2017, 230, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Beccari, G.; Colasante, V.; Tini, F.; Senatore, M.; Prodi, A.; Sulyok, M.; Covarelli, L. Causal agents of Fusarium Head Blight of durum wheat (Triticum durum Desf.) in central Italy and their in vitro biosynthesis of secondary metabolites. Food Microbiol. 2018, 70, 17–27. [Google Scholar] [CrossRef] [PubMed]
- UIFRA. Unión de Industriales Fideeros de la República Argentina. Available online: http://copal.org.ar/wp-content/uploads/2017/06/pastas_secas_argentinas.pdf (accessed on 11 June 2017).
- Commission Regulation, EC N° 1881/2006. Commission regulation no 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union L 2006, 364, 5–24. [Google Scholar]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Edwards, S.; Godley, N. Reduction of Fusarium head blight and deoxynivalenol in wheat with early fungicide applications of prothioconazole. Food Addit. Contam. Part A 2010, 27, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Tóth, B.; Varga, M.; Bartók, T.; Szabó-Hevér, Á.; Farády, L.; Lehoczki-Krsjak, S. Role of fungicides, application of nozzle types, and the resistance level of wheat varieties in the control of Fusarium head blight and deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.; Figlan, S.; Shimelis, H.; Motaung, T.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef]
- Crane, J.; Gibson, D.; Vaughan, R.; Bergstrom, G. Iturin Levels on Wheat Spikes Linked to Biological Control of Fusarium Head Blight by Bacillus amyloliquefaciens. Biol. Control 2013, 103, 146–155. [Google Scholar]
- Dunlap, C.; Bowman, M.; Schisler, D. Genomic analysis and secondary metabolite production in Bacillus amyloliquefaciens AS 43.3: A biocontrol antagonist of Fusarium head blight. Biol. Control. 2013, 64, 166–175. [Google Scholar] [CrossRef]
- Palazzini, J.; Ramirez, M.; Torres, A.; Chulze, S. Potential biocontrol agents for Fusarium Head Blight and deoxynivalenol production in wheat. Crop Prot. 2007, 26, 1702–1710. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Alberione, E.; Torres, A.; Donat, C.; Köhl, J.; Chulze, S. Biological control of Fusarium graminearum sensu stricto, causal agent of Fusarium head blight of wheat, using formulated antagonists under field conditions in Argentina. Biol. Control 2016, 94, 56–61. [Google Scholar] [CrossRef]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J.; Lipps, P.E.; Zhang, S. Selection and evaluation of the potential of choline-metabolizing microbial strains to reduce Fusarium head blight. Biol. Control 2006, 39, 497–506. [Google Scholar] [CrossRef]
- Zhao, Y.; Selvaraj, J.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.; et al. Antagonistic action of Bacillus subtilis strain sg6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Doohan, F. Bacterium-mediated control of Fusarium head blight disease of wheat and barley and associated mycotoxin contamination of grain. Biol. Control 2009, 48, 42–47. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Yerkovich, N.; Alberione, E.; Chiotta, M.; Chulze, S. An integrated dual strategy to control Fusarium graminearum sensu stricto by the biocontrol agent Streptomyces sp. RC 87B under field conditions. Plant. Genet. 2017, 9, 13–18. [Google Scholar] [CrossRef]
- Jochum, C.C.; Osborne, L.E.; Yuen, G.Y. Fusarium head blight biological control with Lysobacter enzymogenes strain C3. Biol. Control 2006, 39, 336–344. [Google Scholar] [CrossRef]
- Schisler, D.A.; Core, A.B.; Boehm, M.J.; Horst, L.; Krause, C.; Dunlap, C.A.; Rooney, A.P. Population dynamics of the Fusarium head blight biocontrol agent Cryptococcus flavescens OH 182.9 on wheat anthers and heads. Biol. Control 2014, 70, 17–27. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.; Voldeng, H.D.; Fedak, G.; Savard, M.E.; Längle, T.; Zhang, J.; Harman, G.E. Concentration and cultivar effects on efficacy of CLO-1 biofungicide in controlling Fusarium head blight of wheat. Biol. Control 2014, 73, 2–7. [Google Scholar] [CrossRef]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.; Chen, W.; Le Floch, G. Challenges facing the biological control strategies for the management of Fusarium Head Blight of cereals caused by F. graminearum. Biol. Control 2017, 113, 26–38. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Palazzini, J.M.; Dunlap, C.A.; Bowman, M.J.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent to reduce Fusarium head blight and deoxynivalenol accumulation: Genome sequencing and secondary metabolite cluster profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Denancé, N.; Sánchez-Vallet, A.; Goffner, D.; Molina, A. Disease resistance on growth: The role of plant hormones in balancing immune response s and fitness costs. Front. Plant Sci. 2013, 4, 155. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Xue, S.; Jia, H.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef] [PubMed]
- Makandar, R.; Nalam, V.; Lee, H.; Trick, H.; Dong, Y.; Shah, J. Salicylic Acid Regulates Basal Resistance to Fusarium Head Blight in Wheat. Mol. Plant-Microbe Interact. 2012, 25, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Henkes, G.; Jousset, A.; Bonkowski, M.; Thorpe, M.; Scheu, S.; Lanoue, A.; Schurr, U.; Ro, U. Pseudomonas fluorescens CHA0 maintains carbon delivery to Fusarium graminearum-infected roots and prevents reduction in biomass of barley shoots through systemic interactions. J. Exp. Bot. 2011, 62, 4337–4344. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Manstretta, V.; Ruggeri, M. A multicomponent decision system to manage Fusarium head blight and mycotoxin in durum wheat. World Mycotoxin J. 2015, 8, 629–640. [Google Scholar] [CrossRef]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J.; Slininger, P. Greenhouse and field evaluation of biological control of Fusarium head blight on durum wheat. Plant Dis. 2002, 86, 1350–1356. [Google Scholar] [CrossRef]
- Palazzini, J.; Ramirez, M.L.; Alberione, E.J.; Torres, A.M.; Chulze, S. Osmotic stress adaptation, compatible solutes accumulation and biocontrol efficacy of two potential biocontrol agents on Fusarium head blight in wheat. Biol. Control 2009, 51, 370–376. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggia, F.; Dalanaj, N.; Prodi, A.; Nipoti, P.; Pisi, A.; Biavati, B.; Di Gioia, D. Microbial inoculants for the biocontrol of Fusarium spp. in durum wheat. BMC Microbiol. 2015, 15, 242. [Google Scholar] [CrossRef] [PubMed]
- Cendoya, E.; Monge, M.; Palacios, S.; Chiacchiera, S.; Torrea, A.; Farnochi, M.; Ramirez, M. Fumonisin occurrence in naturally contaminated wheat grain harvested in Argentina. Food Control 2014, 37, 56–61. [Google Scholar] [CrossRef]
- Horevaj, P.; Milus, E.; Bluhn, B. A real-time qPCR assay to quantify Fusarium graminearum biomass in wheat kernels. J. App. Microbiol. 2011, 111, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Demeke, T.; Grafehan, T.; Clear, R.; Phan, A.; Ratnayaka, I.; Chapados, J.; Patrick, S.; Gaba, D.; Lévesque, A.; Seifert, K. Development of a specific TaqMan® real-time PCR assay for quantification of Fusarium graminearum clade 7 and comparison of fungal biomass determined by PCR with deoxynivalenol content in wheat and barley. Int. J. Food Microbiol. 2010, 141, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Durgbanshi, A.; Arbona, V.; Pozo, O.; Miersch, O.; Sancho, J.; Gomez-Cadenas, A. Simultaneous determination of multiple phytohormones in plants extracts by liquid chromatography electrospray tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 8437–8442. [Google Scholar] [CrossRef] [PubMed]
- Balcke, G.; Handrick, V.; Bergau, N.; Fichtner, M.; Henning, A.; Stellmach, H.; Tissier, A.; Hause, B.; Frolov, A. An UPLC-MS/MS method by highly sensitive high-throughput analysis of phytohromones in plant tissues. Plant Methods 2012, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Iven, T.; König, S.; Singh, S.; Braus-Stromeyer, S.; Bischoff, M.; Tietze, L.; Braus, G.; Lipka, V.; Feussner, I.; Dröge-Laser, W. Transcriptional Activation and Production of Tryptophan-Derived Secondary Metabolites in Arabidopsis Roots Contributes to the Defense against the Fungal Vascular Pathogen Verticillium longisporum. Mol. Plant 2012, 5, 1389–1402. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.; Usall, J.; Teixidó, N.; Delgado, J.; Viñas, I. Water activity, temperature and pH effects on growth of the biocontrol agent Pantoea agglomerans CPA-2. Can. J. Microbiol. 2002, 48, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Palacios, S.; Giaj Merlera, G.; Erazo, J.; Reynoso, M.; Farnochi, C.; Torres, A. Trichothecene genotype and genetic variability of Fusarium graminearum and F. cerealis isolated from durum wheat in Argentina. Eur. J. Plant Pathol. 2017, 149, 969–981. [Google Scholar] [CrossRef]
- Rosewich Gale, L.; Chen, L.; Hernick, C.; Takamura, K.; Kistler, H. Population analysis of Fusarium graminearum from wheat fields in eastern China. Phytopathology 2002, 92, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Wiese, N. Compendium of Wheat Diseases; American Phytopathology Society Press: St. Paul, MN, USA, 1987. [Google Scholar]
- Stack, R.; McMullen, M. A Visual Scale to Estimate Severity of Fusarium Head Blight of Wheat. NDSU Extension Service, North Dakota. 1995, p. 1095. Available online: https://books.google.com.ar/books?id=s4vRGwAACAAJ (accessed on 15 October 2016).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fusarium Head Blight Parameters | ||
|---|---|---|---|
| F. graminearum DNA (pg DNA mg grain−1) * | Fusarium-Damaged Kernels | Deoxynivalenol (µg g−1) | |
| F. graminearum control | 60.7 ± 24.7a | 10.88 ± 2.2a | 4.43 ± 1.7a |
| B. velezensis RC 218 + F. graminearum | 23.8 ± 11.1b | 6 ± 0.58b | 2.2 ± 0.5b |
| S. albidoflavus RC 87B + F. graminearum | 37.1 ± 22.7ab | 7 ± 2.36b | 2.18 ± 0.16b |
| B. velezensis RC 218 | 2.6 ± 1.04b | 5.22 ± 2.5b | Nd |
| S. albidoflavus RC 87B | 7.6 ± 0.4b | 3.44 ± 0.38b | Nd |
| Water Control | 10.1 ± 0.03b | 5.5 ± 3.47b | 0.3 ± 0.13bc |
| Treatments | Inoculum Concentration |
|---|---|
| F. graminearum control | 1 × 105 conidia mL−1 |
| B. velezensis RC 218 + F. graminearum | 1 × 106 cfu mL−1 + 1 × 105 conidia mL−1 |
| B. velezensis RC 218 control | 1 × 106 cfu mL−1 |
| Water Control | -* |
| Treatments | Inoculum Concentration |
|---|---|
| F. graminearum control | 1 × 105 conidia mL−1 |
| B. velezensis RC 218 + F. graminearum | 1 × 106 cfu mL−1 + 1 × 105 conidia mL−1 |
| S. albidoflavus RC 87B + F. graminearum | 1 × 106 cfu mL−1 + 1 × 105 conidia mL−1 |
| B. velezensis RC 218 control | 1 × 106 cfu mL−1 |
| S. albidoflavus RC 87B control | 1 × 106 cfu mL−1 |
| Water Control | -* |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palazzini, J.; Roncallo, P.; Cantoro, R.; Chiotta, M.; Yerkovich, N.; Palacios, S.; Echenique, V.; Torres, A.; Ramirez, M.; Karlovsky, P.; et al. Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists. Toxins 2018, 10, 88. https://doi.org/10.3390/toxins10020088
Palazzini J, Roncallo P, Cantoro R, Chiotta M, Yerkovich N, Palacios S, Echenique V, Torres A, Ramirez M, Karlovsky P, et al. Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists. Toxins. 2018; 10(2):88. https://doi.org/10.3390/toxins10020088
Chicago/Turabian StylePalazzini, Juan, Pablo Roncallo, Renata Cantoro, María Chiotta, Nadia Yerkovich, Sofía Palacios, Viviana Echenique, Adriana Torres, María Ramirez, Petr Karlovsky, and et al. 2018. "Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists" Toxins 10, no. 2: 88. https://doi.org/10.3390/toxins10020088
APA StylePalazzini, J., Roncallo, P., Cantoro, R., Chiotta, M., Yerkovich, N., Palacios, S., Echenique, V., Torres, A., Ramirez, M., Karlovsky, P., & Chulze, S. (2018). Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists. Toxins, 10(2), 88. https://doi.org/10.3390/toxins10020088