The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats

Abstract
1. Introduction
2. Results
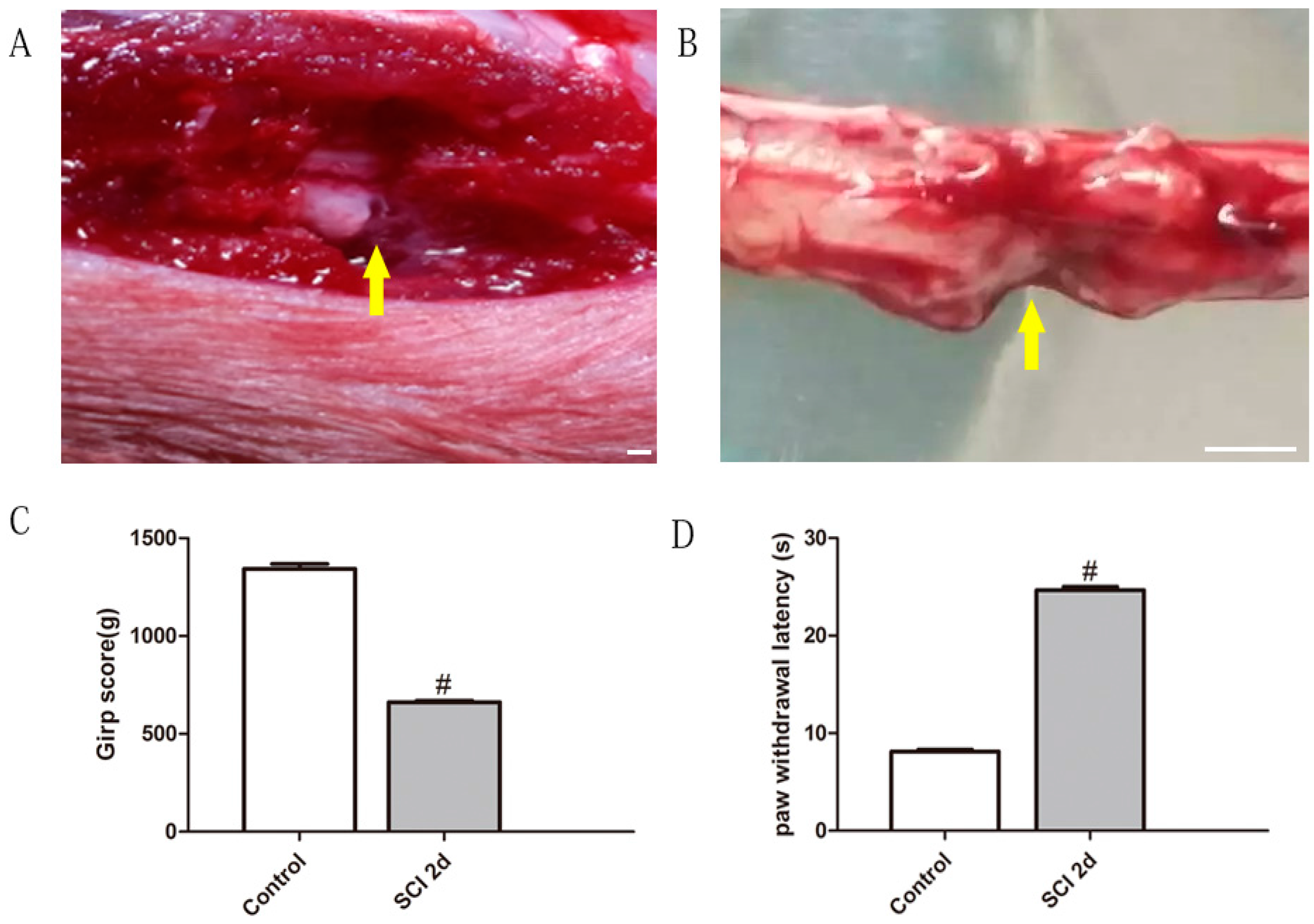
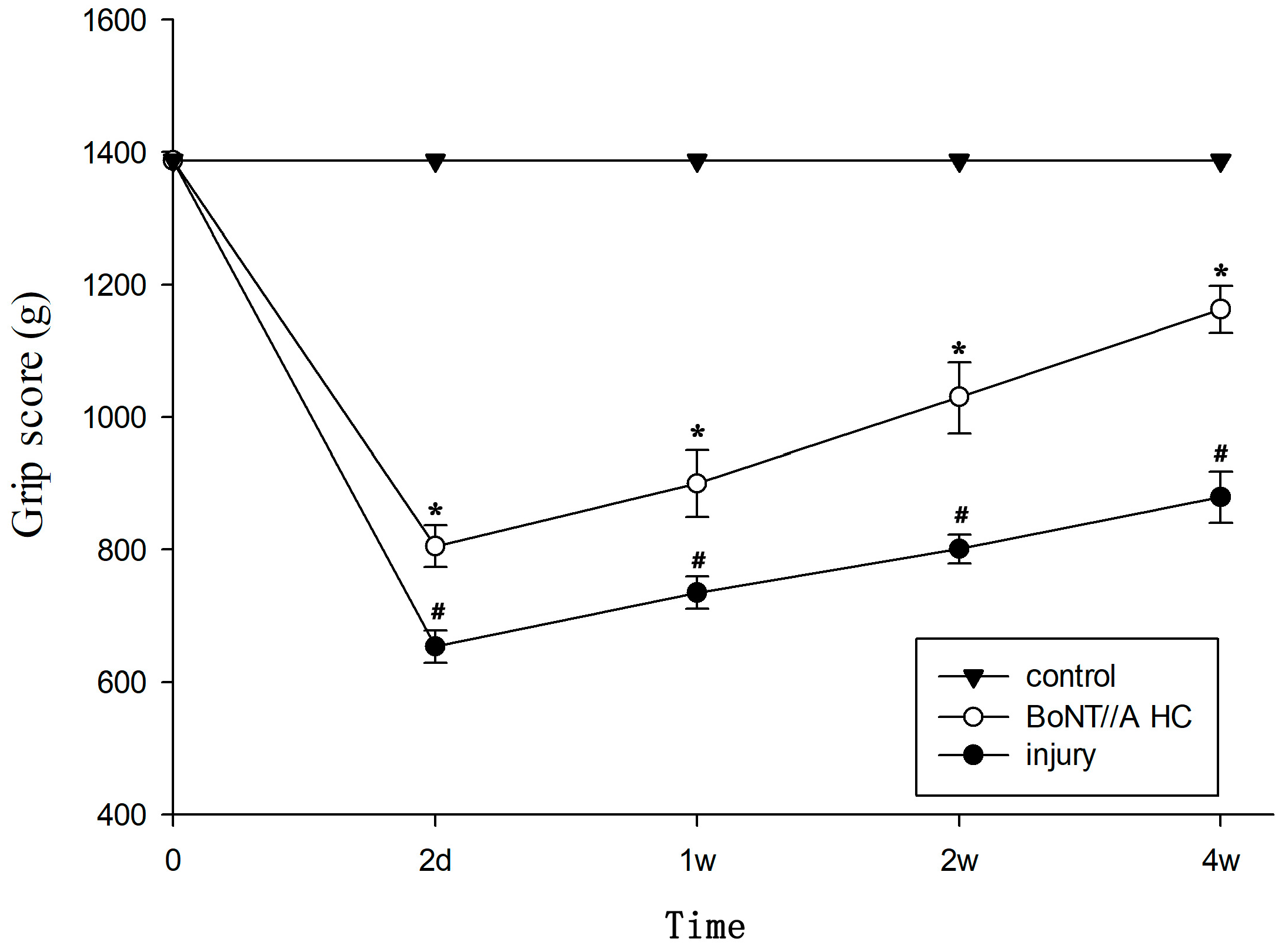
2.1. Unilateral Lumbar SCI Model and Spinal Function Evaluation
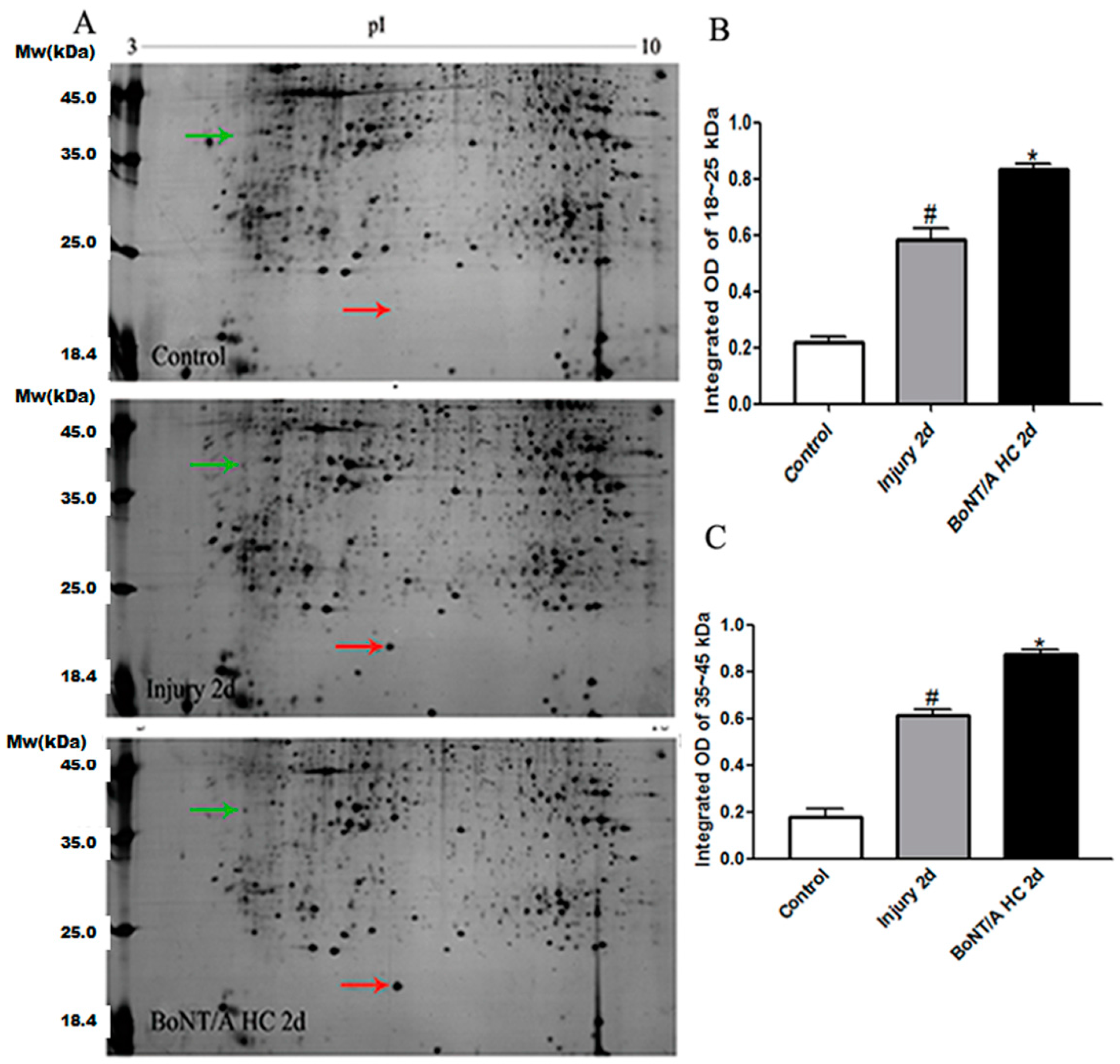
2.2. The Alteration of Local Protein Expression after SCI and the Effect of BoNT/A HC on It
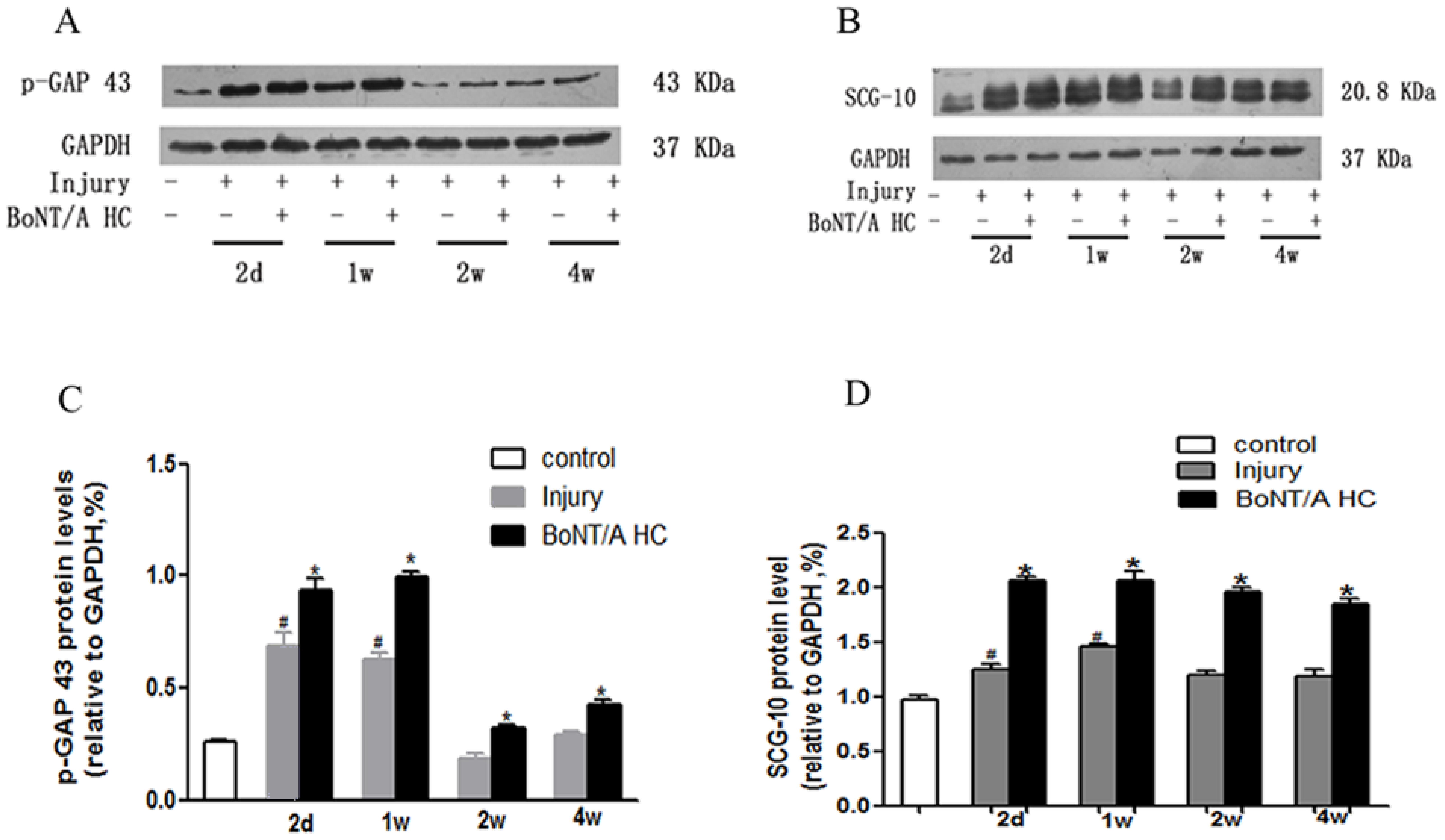
2.3. Effects of BoNT/A Heavy Chain on the Expression of Selective Growth-Associated Proteins
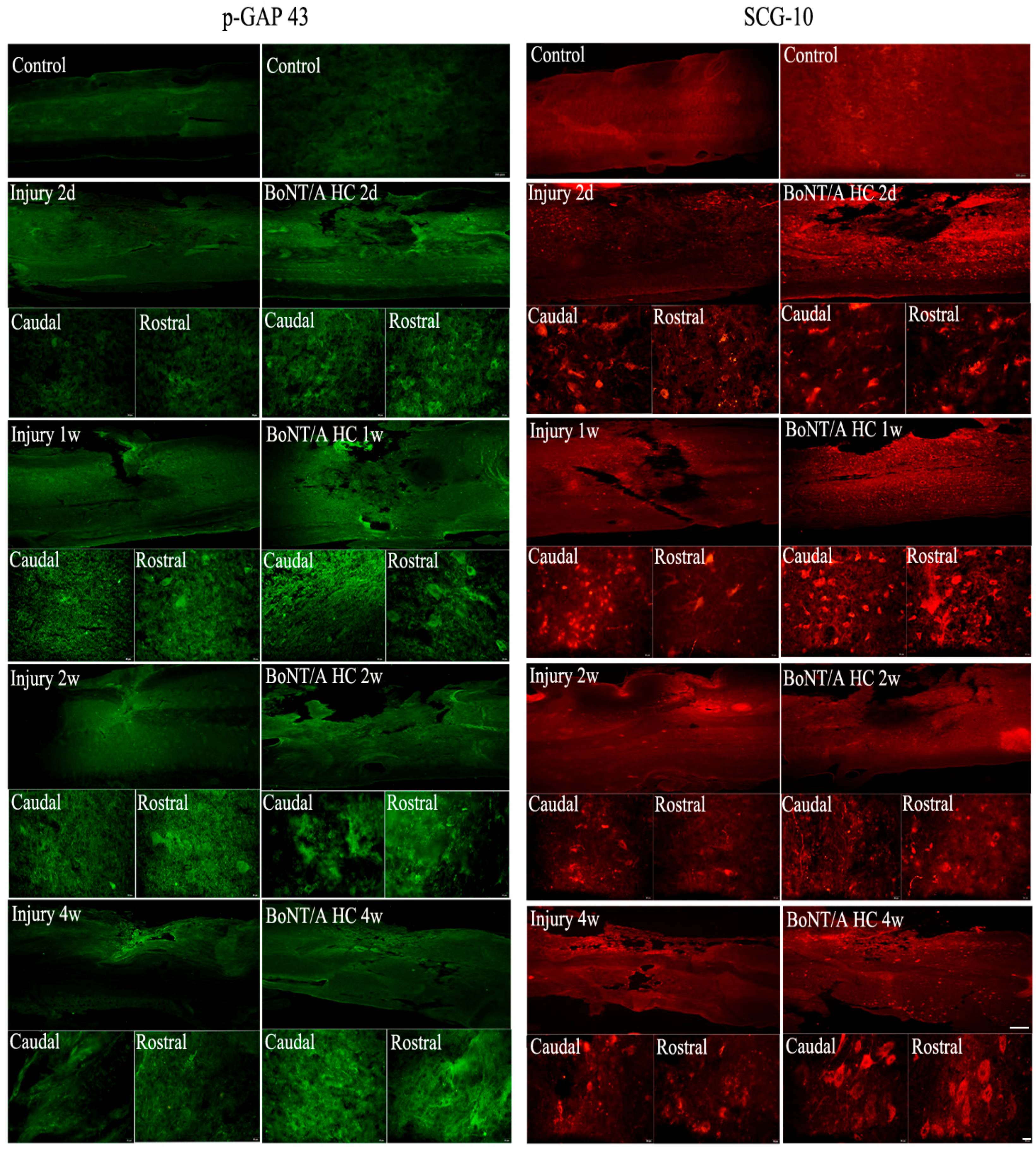
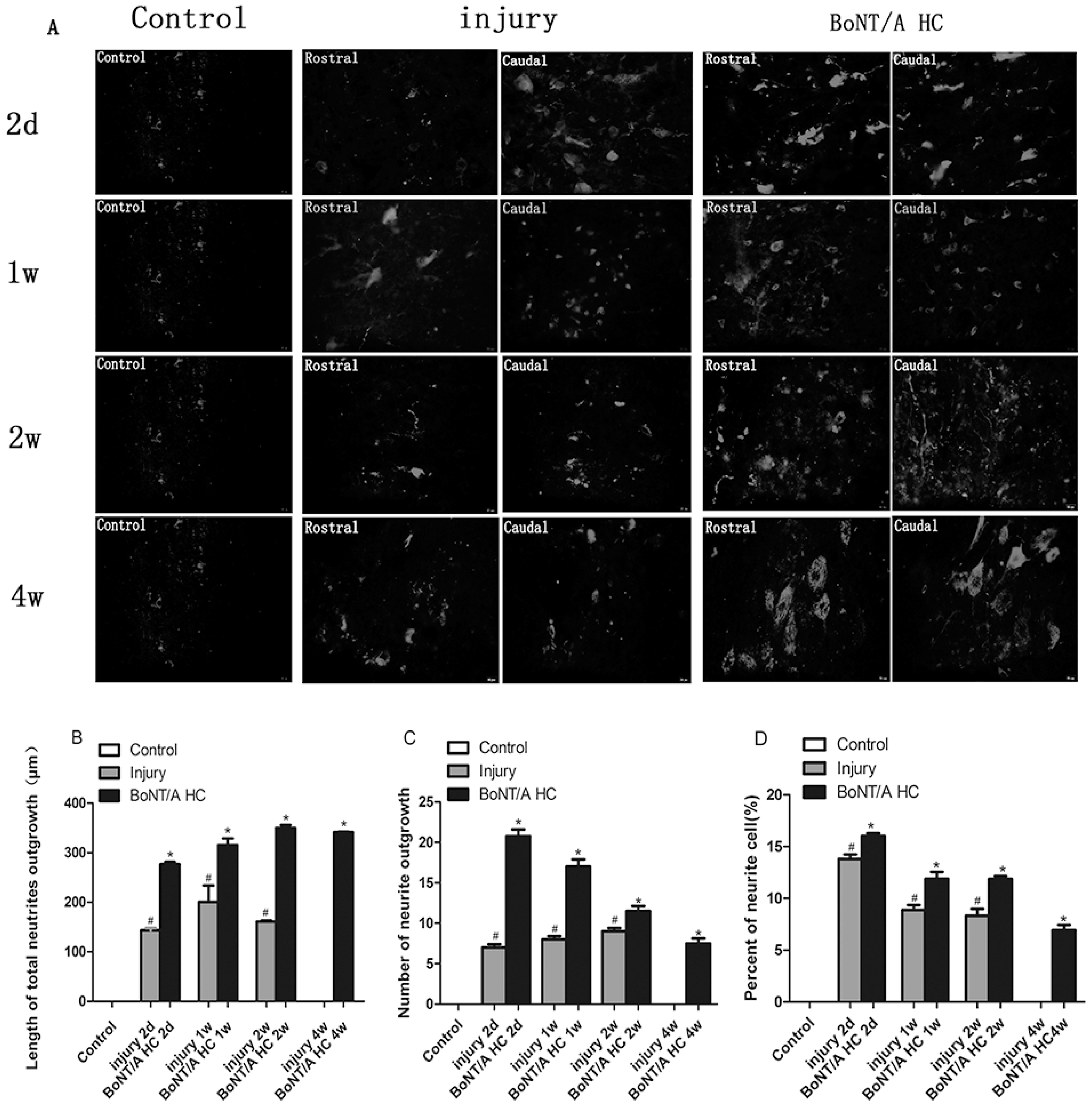
2.4. The Distribution of Positive GAP43 and SCG10 Immunofluorescence at the Injured Area after Administration of the BoNT/A HC
2.5. The Measurement of SCG 10 Positive Neuronal Processes
2.6. The Amelioration of Motor and Sensory Function by BoNT/A HC Treatment Based on SCI
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Establishing the Rat Spinal Cord Hemi-Section Injury Model
5.2. BoNT/A Heavy Chain Administration and Groups
5.3. Two-Dimensional Gel Electrophoresis
5.4. SDS-PAGE and Western Blot
5.5. Immunofluorescence Staining
5.6. Measurement of Neuronal Processes
5.7. Behavioral Evaluation
5.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Giger, R.J.; Hollis, E.R.; Tuszynski, M.H. Guidance molecules in axon regeneration. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.A.; Eng, J.J.; Hsieh, J.T.C.; Wolfe, D.L.; Team, T.S. The health and life priorities of individuals with spinal cord injury: A systematic review. J. Neurotrauma 2012, 29, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, T.A.; Son, Y.J. Extrinsic and intrinsic determinants of nerve regeneration. J. Tissue Eng. 2011, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.T.; Giger, R.J. Insights into the physiological role of CNS regeneration inhibitors. Front. Mol. Neurosci. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Geoffroy, C.G.; Zheng, B. Myelin-associated inhibitors in axonal growth after CNS injury. Curr. Opin. Neurobiol. 2014, 27, 31–38. [Google Scholar] [CrossRef] [PubMed]
- He, Z. Intrinsic control of axon regeneration. J. Biomed. Res. 2010, 24, 2–5. [Google Scholar] [CrossRef]
- Mckerracher, L.; Rosen, K.M. MAG, myelin and overcoming growth inhibition in the CNS. Front. Mol. Neurosci. 2015, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Amemori, T.; Ruzicka, J.; Romanyuk, N.; Jhanwaruniyal, M.; Sykova, E.; Jendelova, P. Comparison of intraspinal and intrathecal implantation of induced pluripotent stem cell-derived neural precursors for the treatment of spinal cord injury in rats. Stem Cell Res. Ther. 2015, 6, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Qin, Y.; Bian, C.; Zhao, Y.; Zhang, W. Intrathecal delivery of il-6 reactivates the intrinsic growth capacity of pyramidal cells in the sensorimotor cortex after spinal cord injury. PLoS ONE 2015, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ray, R.; Singh, B.R.; Ray, P. Mastoparan-7 rescues botulinum toxin-A poisoned neurons in a mouse spinal cord cell culture model. Toxicon 2013, 76, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zichel, R.; Mimran, A.; Keren, A.; Barnea, A.; Steinberger-Levy, I.; Marcus, D.; Turgeman, A.; Reuvenyet, S. Efficacy of a potential trivalent vaccine based on Hc fragments of botulinum toxins A, B, and E produced in a cell-free expression system. Clin. Vaccine Immunol. 2010, 17, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Lisk, G.; Sesardic, D.; Dolly, J.O. Dynamics of motor nerve terminal remodeling unveiled using SNARE-cleaving botulinum toxins: The extent and duration are dictated by the sites of SNAP-25 truncation. Mol. Cell. Neurosci. 2003, 22, 454–466. [Google Scholar] [CrossRef]
- Dolly, O. Synaptic transmission: Inhibition of neurotransmitter release by Botulinum toxins. Headache 2003, 43, 16–24. [Google Scholar] [CrossRef]
- Jankovic, J. Botulinum toxin in clinical practice. J. Neurol. Neurosurg. Psychiatry 2004, 75, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Beske, P.H.; Hoffman, K.M.; Machamer, J.B.; Eisen, M.R.; McNutt, P.M. Use-dependent potentiation of voltage-gated calcium channels rescues neurotransmission in nerve terminals intoxicated by botulinum neurotoxin serotype A. Sci. Rep. 2017, 7, 15862. [Google Scholar] [CrossRef] [PubMed]
- Durham, P.L.; Cady, R.; Cady, R. Regulation of Calcitonin Gene-Related Peptide Secretion From Trigeminal Nerve Cells by Botulinum Toxin Type A: Implications for Migraine Therapy. Headache 2004, 44, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.L.; Zheng, C.X.; Sui, B.D.; Li, Y.Q.; Wang, Y.Y.; Yang, Y.L. A closer look to botulinum neurotoxin type A-induced analgesia. Toxicon 2013, 71, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.R. Review of a Proposed Mechanism for the Antinociceptive Action of Botulinum Toxin Type A. Neurotoxicology 2005, 26, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Jeynes, L.C.; Gauci, C.A. Evidence for the use of botulinum toxin in the chronic pain setting: A review of the literature. Pain Pract. 2008, 8, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.R. Future aspects of botulinum neurotoxins. J. Neural Trans. 2008, 115, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Thenganatt, M.A.; Fahn, S. Botulinum toxin for the treatment of movement disorders. Curr. Neurol. Neurosci. Rep. 2012, 12, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Foster, L.; Clapp, L.; Erickson, M.; Jabbari, B. Botulinum toxin A and chronic low back pain: A randomized, double-blind study. Neurology 2001, 56, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Ayyar, B.V.; Tajhya, R.B.; Beeton, C.; Atassi, M.Z. Antigenic sites on the H N domain of botulinum neurotoxin A stimulate protective antibody responses against active toxin. Sci. Rep. 2015, 5, 15776. [Google Scholar] [CrossRef] [PubMed]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2002, 82, 427–446. [Google Scholar] [CrossRef]
- Juzans, P.; Comella, J.X.; Molgo, J.; Faille, L.; Angaut-Petit, D. Nerve terminal sprouting in botulinum type-A treated mouse levator auris longus muscle. Neuromuscul. Disord. 1996, 6, 177–185. [Google Scholar] [CrossRef]
- Foran, P.G.; Davletov, B.; Meunier, F.A. Getting musclesmoving again after botulinum toxin: Novel therapeuti challenges. Trends Mol. Med. 2003, 9, 291–299. [Google Scholar] [CrossRef]
- Ko, C.P. Do nerve terminal sprouts contribute to functional recovery from botulinum neurotoxin A? J. Physiol. 2008, 586, 3021. [Google Scholar] [CrossRef] [PubMed]
- Coffield, J.A.; Yan, X. Neuritogenic actions of botulinum neurotoxin A on cultured motor neurons. J. Pharmacol. Exp. Ther. 2009, 330, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Morii, H. SCG10-related neuronal growth-associated proteins in neural development, plasticity, degeneration, and aging. J. Neurosci. Res. 2002, 70, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Pellier-Monnin, V.; Astic, L.; Bichet, S.; Riederer, B.M.; Grenningloh, G. Expression of SCG10 and stathmin proteins in the rat olfactory system during development and axonal regeneration. J. Comp. Neurol. 2001, 433, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Holahan, M.R. A Shift from a Pivotal to Supporting Role for the Growth-Associated Protein (GAP-43) in the Coordination of Axonal Structural and Functional Plasticity. Front. Cell. Neurosci. 2017, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.L.; Klassen, M.P.; Hua, Y.; Barres, B.A. Amacrine-signaled loss of intrinsic axon growth ability by retinal ganglion cells. Science 2002, 296, 1860–1864. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.J.; Fan, D.P.; Tsui, B.J.; Cassar, S.L.; Tetzlaff, W. Influence of the axotomy to cell body distance in rat rubrospinal and spinal motoneurons: Differential regulation of gap-43, tubulins, and neurofilamentm. J. Comp. Neurol. 1999, 414, 495–510. [Google Scholar] [CrossRef]
- Fagoe, N.D.; Heest, J.V.; Verhaagen, J. Spinal cord injury and the neuron-intrinsic regeneration-associated gene program. Neuromol. Med. 2014, 16, 799–813. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, A.R. The axon reaction: A review of the principal features of perikaryal responses to axon injury. Int. Rev. Neurobiol. 1971, 14, 49–124. [Google Scholar] [PubMed]
- Grafstein, B. The nerve cell body response to axotomy. Exp. Neurol. 1975, 48, 32–51. [Google Scholar] [CrossRef]
- Skene, J.H.; Willard, M. Axonally Transported Proteins Associated with Axon Growth in Rabbit Central and Peripheral Nervous Systems. J. Cell Biol. 1981, 89, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Skene, J.H. Axonal growth-associated proteins. Annu. Rev. Neurosci. 1989, 12, 127–156. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, W.; Alexander, S.W.; Miller, F.D.; Bisby, M.A. Response of facial and rubrospinal neurons to axotomy: Changes in mrna expression for cytoskeletal proteins and gap-43. J. Neurosci. 1991, 11, 2528–2544. [Google Scholar] [PubMed]
- Shin, J.E.; Geisler, S.; Diantonio, A. Dynamic regulation of SCG10 in regenerating axons after injury. Exp. Neurol. 2014, 252, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ozon, S.; Maucuer, A.; Sobel, A. The stathmin family—Molecular and biological characterization of novel mammalian proteins expressed in the nervous system. Eur. J. Biochem. 2010, 248, 794–806. [Google Scholar] [CrossRef]
- Riederer, B.M.; Pellier, V.; Antonsson, B.; Paolo, G.D.; Stimpson, S.A.; Lütjens, R.; Catsicas, S.; Grenningloh, G. Regulation of microtubule dynamics by the neuronal growth-associated protein SCG 10. Proc. Natl. Acad. Sci. USA 1997, 94, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shan, C.; Cao, W.; Zhang, C.; Teng, J.; Chen, J. SCG10 promotes non-amyloidogenic processing of amyloid precursor protein by facilitating its trafficking to the cell surface. Hum. Mol. Genet. 2013, 22, 4888–4900. [Google Scholar] [CrossRef] [PubMed]
- Prado, W.A. Antinociceptive potency of intrathecal morphine in the rat tail flick test: A comparative study using acute lumbar catheter in rats with or without a chronic atlanto-occipital catheter. J. Neurosci. Methods 2003, 129, 33–39. [Google Scholar] [CrossRef]
- Ulmann, L.; Rodeau, J.L.; Danoux, L.; Contet-Audonneau, J.L.; Pauly, G.; Schlichter, R. Dehydroepiandrosterone and neurotrophins favor axonal growth in a sensory neuron-keratinocyte coculture model. Neuroscience 2009, 159, 514–525. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 0 d | 2 d | 1 w | 2 w | 4 w |
|---|---|---|---|---|---|
| Control | 8.5 ± 0.2 | 8.5 ± 0.2 | 8.3 ± 0.1 | 7.8 ± 0.2 | 8.2 ± 0.2 |
| Injury | 8.5 ± 0.2 | 22.8 ± 1.0 # | 22.1 ± 0.9 # | 22.4 ± 1.0 # | 21.5 ± 0.9 # |
| BoNT/A HC | 8.5 ± 0.2 | 24.4 ± 0.3 | 24.5 ± 0.3 | 24.1 ± 0.4 | 22.5 ± 0.9 |
| Name of Group | Pseudo Surgery | SCI without BoNT/A HC | SCI with BoNT/A HC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Time | 0 | 2 d | 1 wk | 2 wks | 4 wks | 2 d | 1 wk | 2 wks | 4 wks |
| No. of animals | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-F.; Liu, F.; Lan, J.; Bai, J.; Li, X.-Q. The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats. Toxins 2018, 10, 66. https://doi.org/10.3390/toxins10020066
Wang Y-F, Liu F, Lan J, Bai J, Li X-Q. The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats. Toxins. 2018; 10(2):66. https://doi.org/10.3390/toxins10020066
Chicago/Turabian StyleWang, Ya-Fang, Fu Liu, Jing Lan, Juan Bai, and Xia-Qing Li. 2018. "The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats" Toxins 10, no. 2: 66. https://doi.org/10.3390/toxins10020066
APA StyleWang, Y.-F., Liu, F., Lan, J., Bai, J., & Li, X.-Q. (2018). The Effect of Botulinum Neurotoxin Serotype a Heavy Chain on the Growth Related Proteins and Neurite Outgrowth after Spinal Cord Injury in Rats. Toxins, 10(2), 66. https://doi.org/10.3390/toxins10020066