Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin

 ,
,  , ,
, , 

Abstract
:1. Introduction
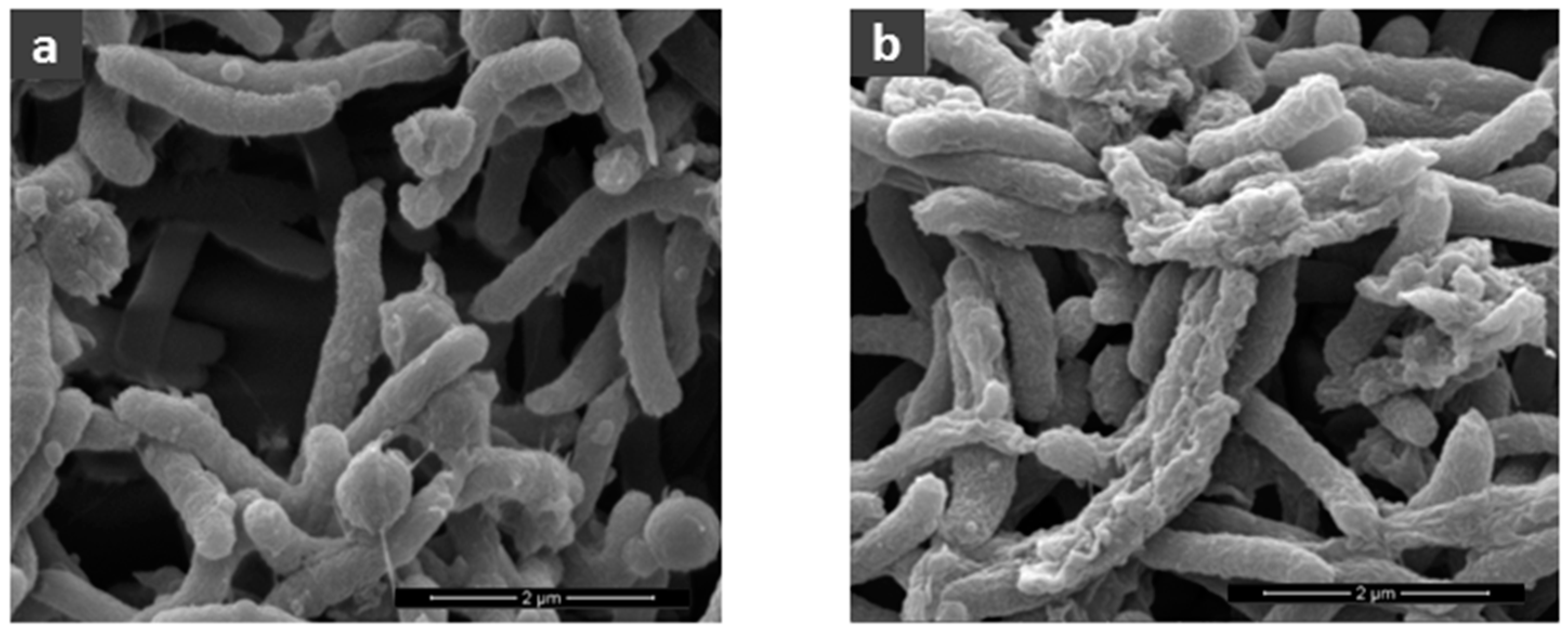
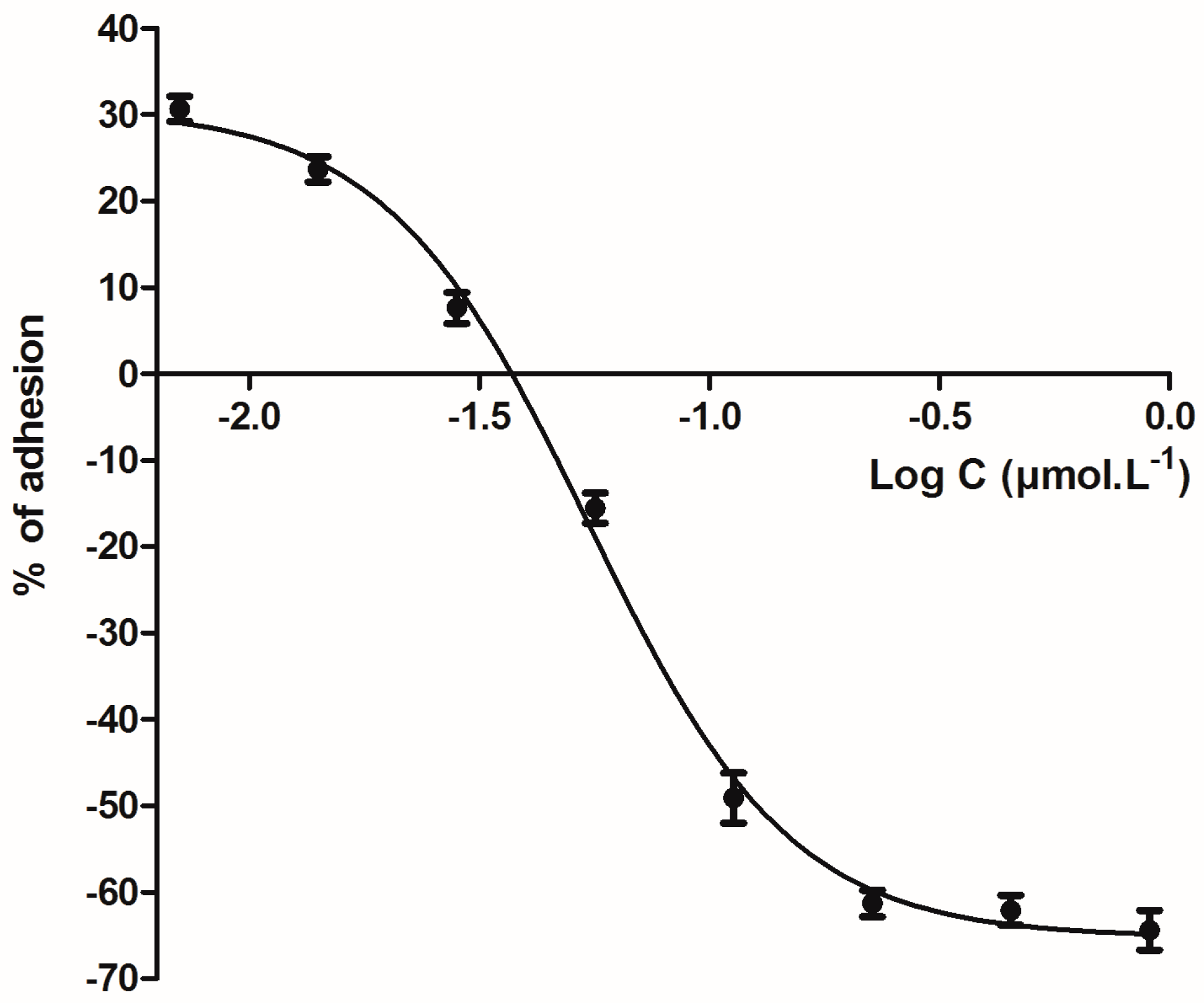
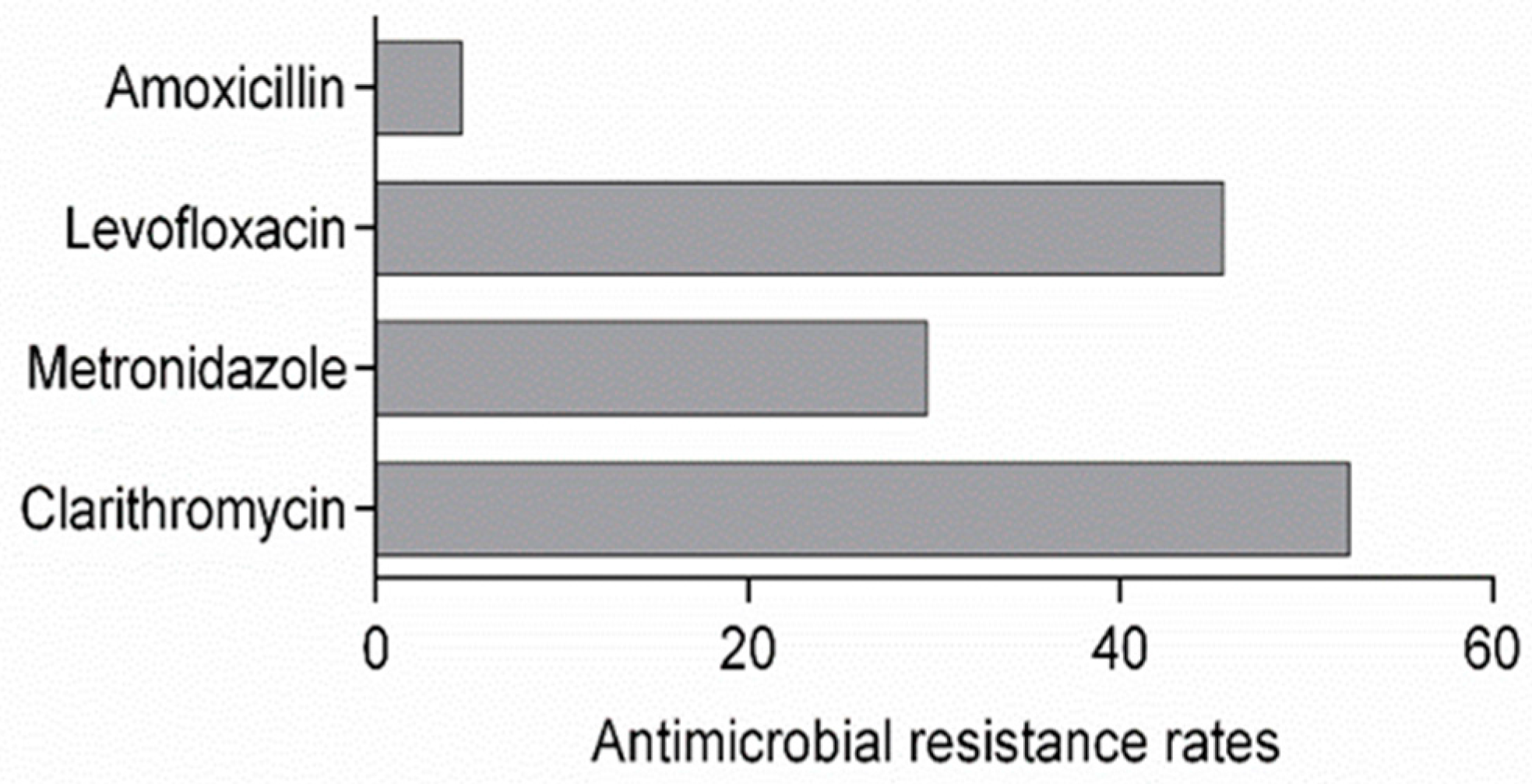
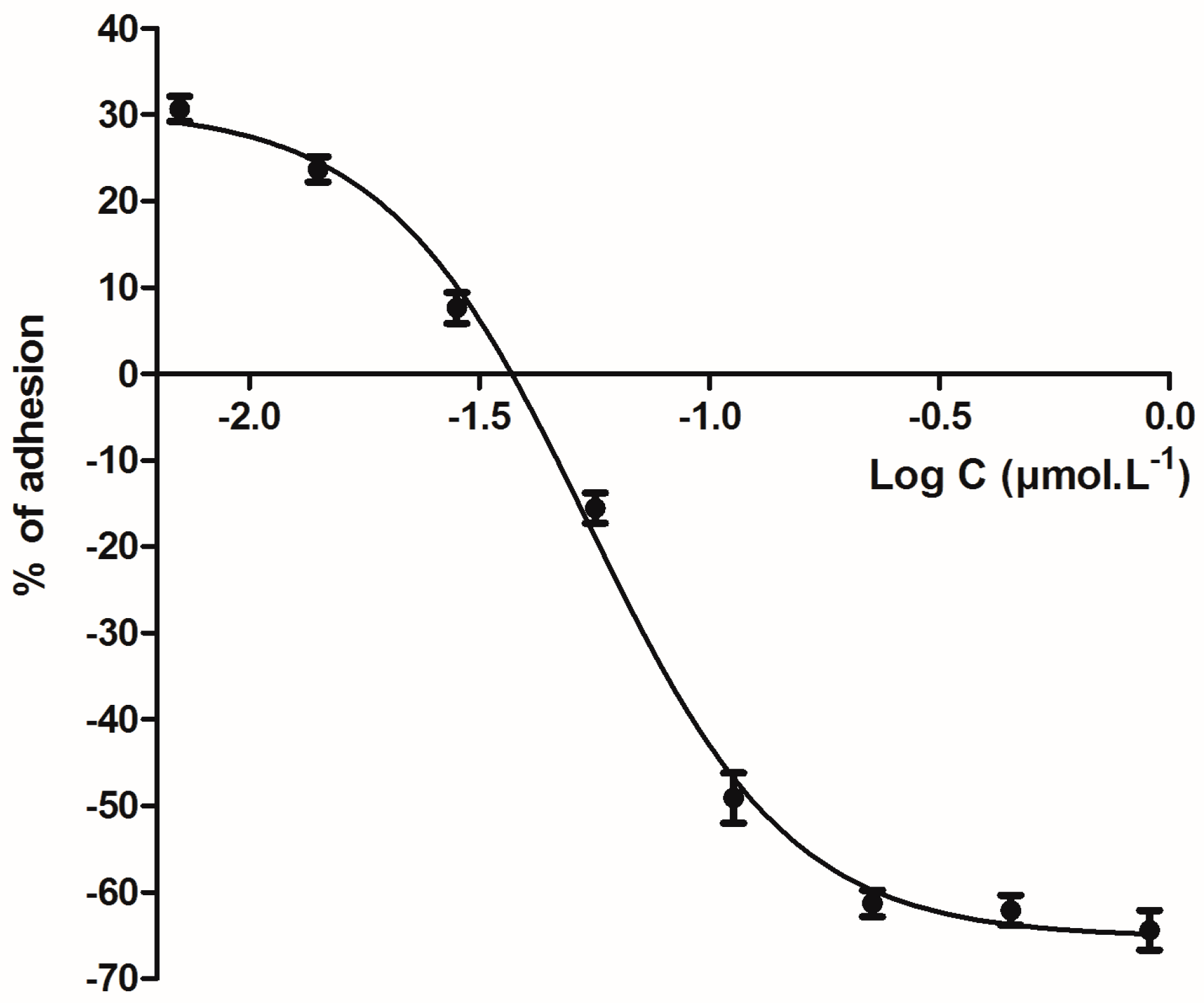
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bicarinalin Synthesis
5.2. Microorganism Strains and Growth Conditions
5.3. Antimicrobial Assays
5.4. Anti-Adherence Effect
5.5. Cytotoxicity of Bicarinalin
5.6. Scanning Electron Microscopy
5.6.1. Helicobacter SEM
5.6.2. Gastric cells SEM
5.6.3. Scanning Electron Microscopy
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Keilberg, D.; Ottemann, K.M. How Helicobacter pylori senses, targets and interacts with the gastric epithelium. Environ. Microbiol. 2016, 18, 791–806. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, M.; Cerda, O. Overview on chemotaxis and acid resistance in Helicobacter pylori. Biol. Res. 2003, 36, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Eusebi, L.H.; Zagari, R.M.; Bazzoli, F. Epidemiology of Helicobacter pylori infection. Helicobacter 2014, 19, 1–5. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Helicobacter pylori and Cancer. 2013. Available online: https://www.cancer.gov/about-cancer/causes-prevention/risk/infectious-agents/h-pylori-fact-sheet#q1 (accessed on 13 September 2017).
- Dubois, A. Spiral bacteria in the human stomach: The gastric helicobacters. Emerg. Infect. Dis. 1995, 1, 79–85. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). Schistosomes, Liver flukes and Helicobacter pylori: Monographs on the evaluation of Carcinogenic Risks to Human. IARC: Lyon, Francia, France; 1994. IARC Sci. Publ. 1994, 6, 177–241. [Google Scholar]
- European Helicobacter Pylori Study Group (EHPSG). Current European concepts in the management of Helicobacter pylori infection. The Maastricht Consensus Report. Gut 1997, 41, 8–16. [Google Scholar]
- Espino, A. Infección por Helicobacter pylori. Gastroenterol. Latinoam. 2010, 21, 323–327. [Google Scholar]
- Cogo, L.L.; Monteiro, C.L.B.; Miguel, M.D.; Miguel, O.G.; Cunico, M.M.; Ribeiro, M.L.; de Carmago, E.R.; Kussen, G.M.B.; da Silva Nogueira, K.; Dalla Costa, L.M. Anti-Helicobacter pylori activity of plant extracts traditionally used for the treatment of gastrointestinal disorders. Brazilian J. Microbiol. 2010, 41, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Aili, S.R.; Fox, E.G.P.; Escoubas, P.; Orivel, J.; Nicholson, G.M.; Dejean, A. The biochemical toxin arsenal from ant venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Vogel, H.J. Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta 1999, 1462, 11–28. [Google Scholar] [CrossRef]
- Li, Y.; Xiang, Q.; Zhang, Q.; Huang, Y.; Su, Z. Overview on the recent study of antimicrobial peptides: Origins, functions, relative mechanisms and application. Peptides 2012, 37, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Téné, N.; Bonnafé, E.; Berger, F.; Rifflet, A.; Guilhaudis, L.; Ségalas-Milazzo, I.; Pipy, B.; Coste, A.; Leprince, J.; Treilhou, M. Biochemical and biophysical combined study of bicarinalin, an ant venom antimicrobial peptide. Peptides 2016, 79, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-J.; Park, S.J.; Mishig-Ochir, T.; Lee, B.-J. Antimicrobial peptides: Therapeutic potentials. Expert Rev. Anti-Infect. Ther. 2014, 12, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Dai, C.; Li, Z.; Fan, Z.; Song, Y.; Wu, Y.; Cao, Z.; Li, W. Antibacterial activity and mechanism of a scorpion venom peptide derivative in vitro and in vivo. PLoS ONE 2012, 7, e40135. [Google Scholar] [CrossRef] [PubMed]
- Abreu, T.F.; Sumitomo, B.N.; Nishiyama, M.Y.; Oliveira, U.C.; Souza, G.H.M.F.; Kitano, E.S.; Zelanis, A.; Serrano, S.M.T.; Junqueira, I.; Azevedo, D.; et al. Peptidomics of Acanthoscurria gomesiana spider venom reveals new toxins with potential antimicrobial activity. J. Proteom. 2017, 151, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Kong, Y.; Zhai, L.; Wu, X.; Jia, P.; Liu, J.; Yu, H. Two novel antimicrobial peptides from centipede venoms. Toxicon 2010, 55, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, A.; Roberto, J.; Musacchio, A.; Sergio, M.; Brochetto-Braga, M.R. Wasp venomic: Unravelling the toxins arsenal of Polybia paulista venom and its potential pharmaceutical applications. J. Proteom. 2017, 161, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Pluzhnikov, K.A.; Kozlov, S.A.; Vassilevski, A.A.; Vorontsova, O.V.; Feofanov, A.V.; Grishin, E.V. Linear antimicrobial peptides from Ectatomma quadridens ant venom. Biochimie 2014, 107, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Wanandy, T.; Gueven, N.; Davies, N.W.; Brown, S.G.A.; Wiese, M.D. Pilosulins: A review of the structure and mode of action of venom peptides from an Australian ant Myrmecia pilosula. Toxicon 2015, 98, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Rifflet, A.; Gavalda, S.; Téné, N.; Orivel, J.; Leprince, J.; Guilhaudis, L.; Génin, E.; Treilhou, M. Identification and characterization of a novel antimicrobial peptide from the venom of the ant Tetramorium bicarinatum. Peptides 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Téné, N.; Roche-Chatain, V.; Rifflet, A.; Bonnafé, E.; Lefranc, B.; Leprince, J.; Treilhou, M. Potent bactericidal effects of bicarinalin against strains of the Enterobacter and Cronobacter genera. Food Control 2014, 42, 202–206. [Google Scholar] [CrossRef]
- Ramos, A.R.; Sánchez, R.S. Helicobacter pylori 25 anos después (1983–2008): Epidemiologia, microbiologia, patogenia, diagnóstico y tratamiento. Rev. Gastroenterol. Perú 2009, 29, 158–170. [Google Scholar]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.; Li, J.; Xu, X.; Lai, R.; Zou, Q. An antimicrobial peptide with antimicrobial activity against Helicobacter pylori. Peptides 2007, 28, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Narayana, J.L.; Huang, H.; Wu, C.; Chen, J. Epinecidin-1 antimicrobial activity: In vitro membrane lysis and In vivo efficacy against Helicobacter pylori infection in a mouse model. Biomaterials 2015, 61, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Ofek, I.; Hasty, D.L.; Sharon, N. Anti-adhesion therapy of bacterial diseases: Prospects and problems. FEMS Immunol. Med. Microbiol. 2003, 38, 181–191. [Google Scholar] [CrossRef]
- Wittschier, N.; Lengsfeld, C.; Vorthems, S.; Stratmann, U.; Ernst, J.F.; Verspohl, E.J.; Hensel, A. Large molecules as anti-adhesive compounds against pathogens. J. Pharm. Pharmacol. 2007, 59, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Wittschier, N.; Faller, G.; Hensel, A. Aqueous extracts and polysaccharides from liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to human gastric mucosa. J. Ethnopharmacol. 2009, 125, 218–223. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, R.; Al-Khtheeri, H.; Weerasekera, D.; Fernando, N.; Vaira, D.; Holton, J.; Basset, C. Bactericidal and anti-adhesive properties of culinary and medicinal plants against Helicobacter pylori. World J. Gastroenterol. 2005, 11, 7499–7507. [Google Scholar] [CrossRef] [PubMed]
- Ndip, R.N.; MacKay, W.G.; Farthing, M.J.G.; Weaver, L.T. Culturing Helicobacter pylori from Clinical Specimens: Review of Microbiologic Methods. J. Pedriatr. Gasteroenterol. Nutr. 2003, 36, 616–622. [Google Scholar] [CrossRef]
- Rimbar, E.; Sasatsu, M.; Graham, D.Y. PCR detection of Helicobacter pylori in clinical samples. Methods Mol. Biol. 2013, 943, 279–287. [Google Scholar] [CrossRef]
- NHS. NHS UK Standards for Microbiology Investigations. In Identification of Helicobacter Species; Service NH: London, UK, 2015. [Google Scholar]
- CLSI. CLSI Methods for antimicrobial dilution and disk susceptibility testing of infrequently isolated or fastidious bacteria. In Approved Guideline, 2nd ed.; PA Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2010. [Google Scholar]
- Piccolomini, R.; Di Bonaventura, G.; Festi, D.; Catamo, G.; Laterza, F.; Neri, M. Optimal combination of media for primary isolation of Helicobacter pylori from gastric biopsy specimens. J. Clin. Microbiol. 1997, 35, 1541–1544. [Google Scholar] [PubMed]
- Hachem, C.Y.; Clarridge, J.E.; Reddy, R.; Flamm, R.; Evans, D.G.; Tanaka, S.K.; Graham, D.Y. Antimicrobial susceptibility testing of Helicobacter pylori: Comparison of E-test, broth microdilution and disk diffusion for ampicillin, clarithromycin and metronidazole. Diagn. Microbiol. Infect. Dis. 1996, 24, 37–41. [Google Scholar] [CrossRef]
- EUCAST. EUCAST Breakpoint Tables for Interpretation of MICs and Zone Diameters; Contract No.: Version 5.0.; European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2015. [Google Scholar]
- Diesing, A.; Nossol, C.; Faber-Zuschratter, H.; Zuschratter, W.; Renner, L.; Sokolova, O.; Naumann, M.; Rothkotter, H.-J. Rapid interaction of Helicobacter pylori with microvilli of the polar human gastric epithelial cell line NCI-N87. Anat. Rec. 2013, 296, 1800–1805. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Lee, K.L.; Kim, B.K.; Kim, J.W.; Jeong, J.B.; Kim, S.G.; Kim, J.S.; Jung, H.C.; Song, I.S. Usefulness of NCI-N87 cell lines in Helicobacter pylori infected gastric mucosa model. Korean J. Gastroenterol. 2006, 47, 357–362. [Google Scholar] [PubMed]
- Horemans, T.; Kerstens, M.; Clais, S.; Struijs, K.; van den Abbeele, P.; Van Assche, T.; Maes, L.; Cos, P. Evaluation of the anti-adhesive effect of milk fat globule membrane glycoproteins on Helicobacter pylori in the human NCI-N87 cell line and C57BL/6 mouse model. Helicobacter 2012, 17, 312–318. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. pylori Strain | MIC50 μmol·L−1 (μg·mL−1) | ||||
|---|---|---|---|---|---|
| Bicarinalin | Clarithromycin | Levofloxacin | Metronidazole | Amoxicillin | |
| ATCC 43504 | 3.9 (8.6) | 0.042 (0.03) | 0.17 (0.06) | 374.4 (64) | 0.035 (0.014) |
| Peruvian patients | 0.99 (2.2) | 0.66 (0.5) | 1.94 (0.7) | 23.4 (4) | <0.082 (0.03) |
| CC50 (μmol·L−1) | SI | |
|---|---|---|
| Peritoneal macrophages (Balb/C) | 39.2 | >39 a |
| Gastric cells (N87) | 1.7 * | >17 b |
| a: SI = CC50 */MIC50; b: SI = CC50 **/IC50 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzman, J.; Téné, N.; Touchard, A.; Castillo, D.; Belkhelfa, H.; Haddioui-Hbabi, L.; Treilhou, M.; Sauvain, M. Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin. Toxins 2018, 10, 21. https://doi.org/10.3390/toxins10010021
Guzman J, Téné N, Touchard A, Castillo D, Belkhelfa H, Haddioui-Hbabi L, Treilhou M, Sauvain M. Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin. Toxins. 2018; 10(1):21. https://doi.org/10.3390/toxins10010021
Chicago/Turabian StyleGuzman, Jesus, Nathan Téné, Axel Touchard, Denis Castillo, Haouaria Belkhelfa, Laila Haddioui-Hbabi, Michel Treilhou, and Michel Sauvain. 2018. "Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin" Toxins 10, no. 1: 21. https://doi.org/10.3390/toxins10010021
APA StyleGuzman, J., Téné, N., Touchard, A., Castillo, D., Belkhelfa, H., Haddioui-Hbabi, L., Treilhou, M., & Sauvain, M. (2018). Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin. Toxins, 10(1), 21. https://doi.org/10.3390/toxins10010021