Characterization and Interrelations of One-Carbon Metabolites in Tissues, Erythrocytes, and Plasma in Mice with Dietary Induced Folate Deficiency


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
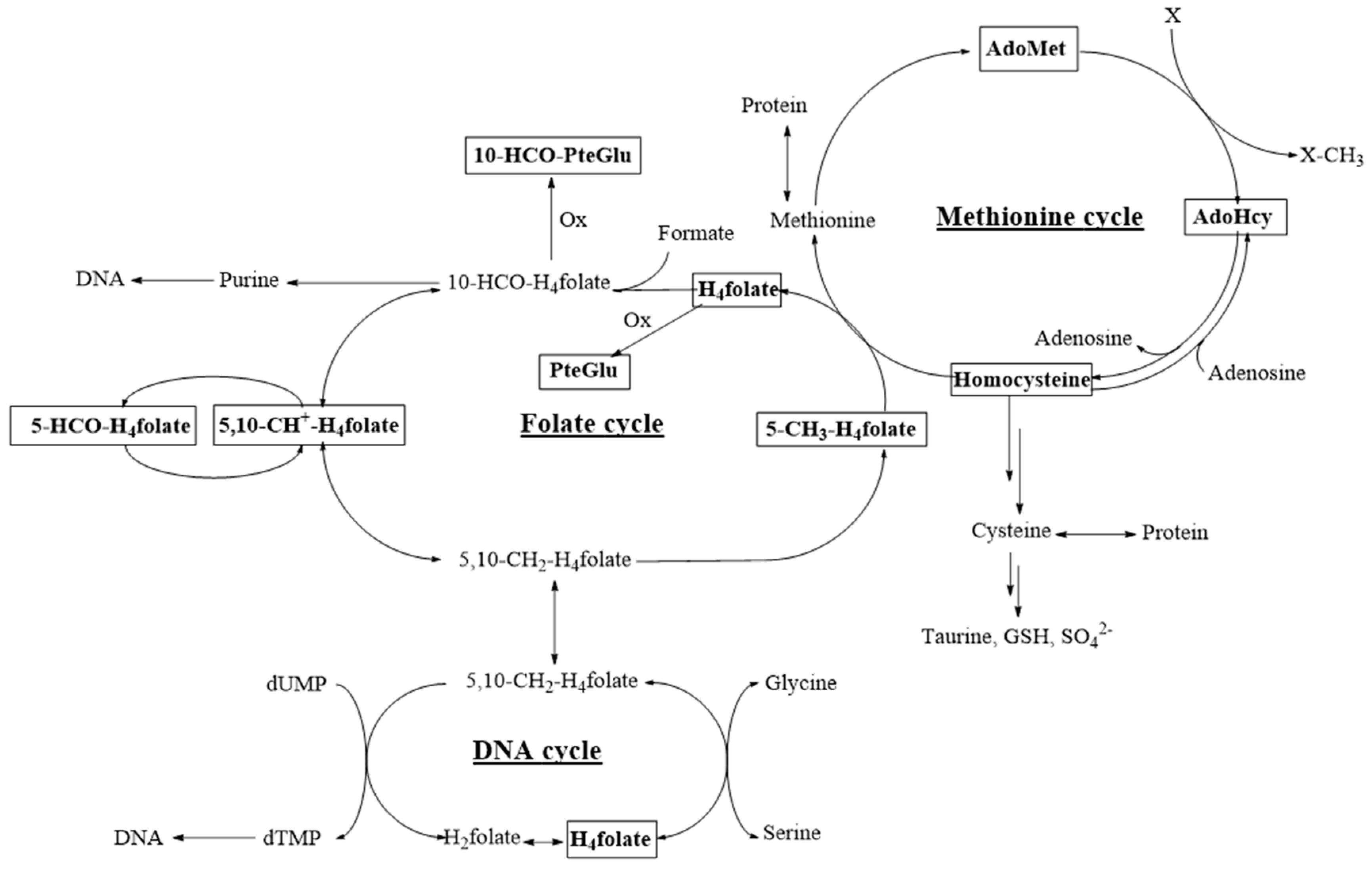
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Solutions
2.2. Animals and Ethics Approval
2.3. Study Design
2.4. Sampling Procedure
2.5. Folate in Liver, Brain, Heart, Kidney, and Erythrocytes
2.6. Solid-Phase Extraction (SPE)
2.7. AdoMet and AdoHcy in Liver, Brain, Heart, and Kidney
2.8. tHcy in Plasma
2.9. LC-MS/MS
2.10. Data Analysis
3. Results
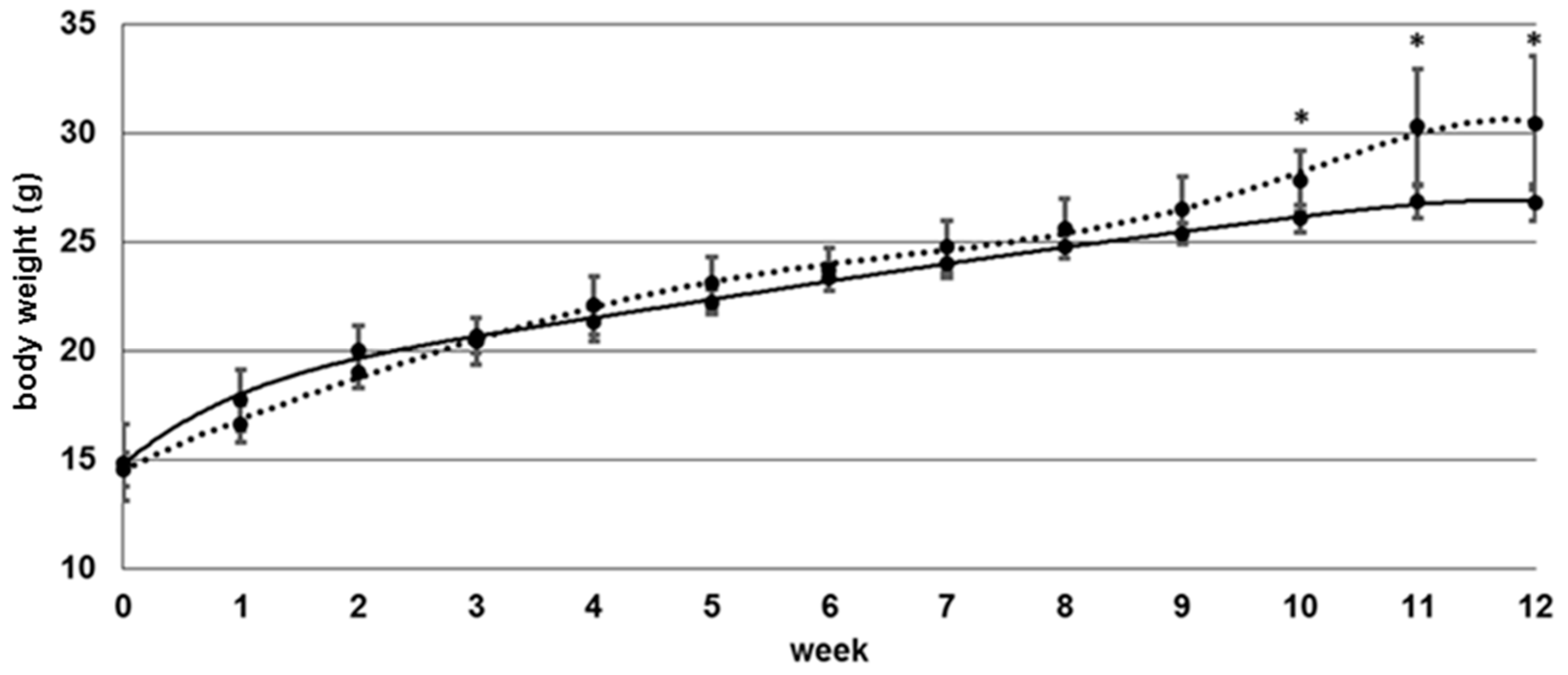
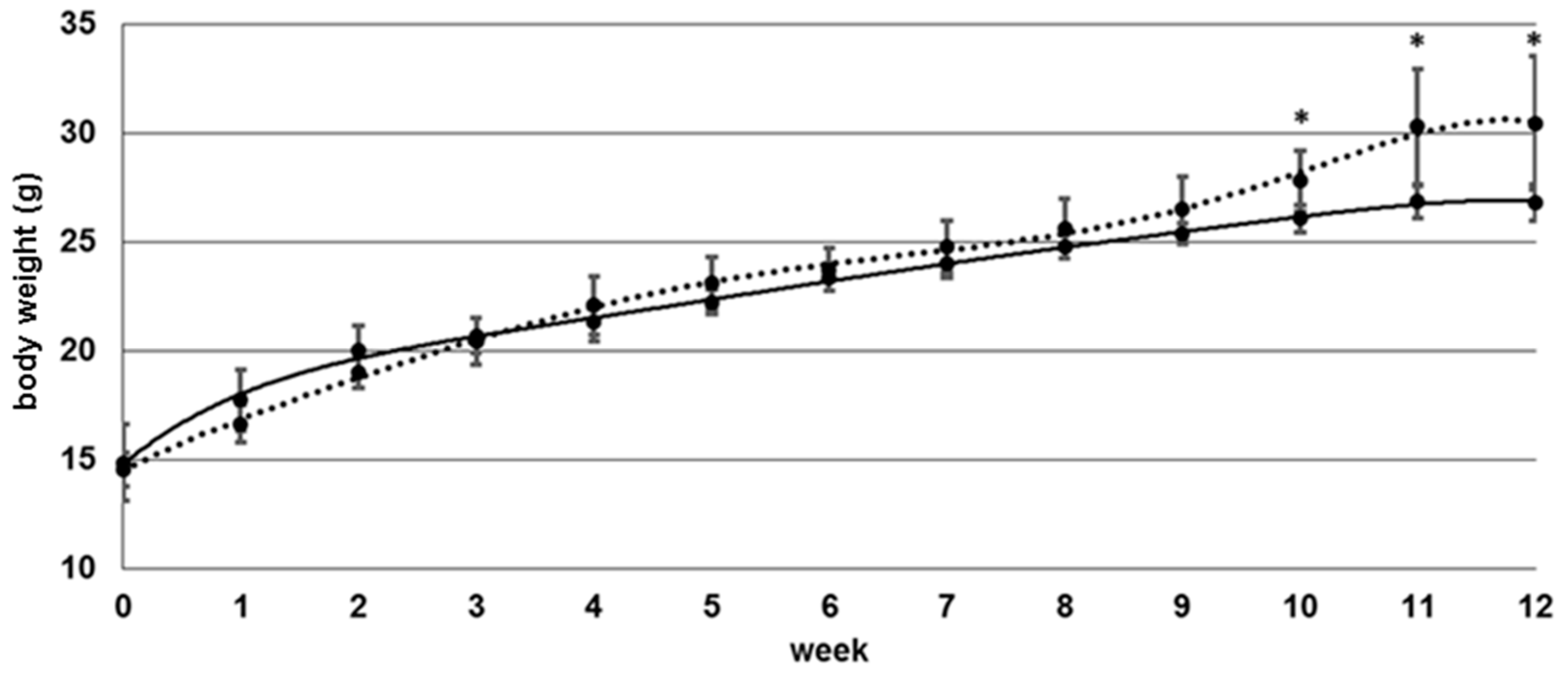
3.1. Body Weight
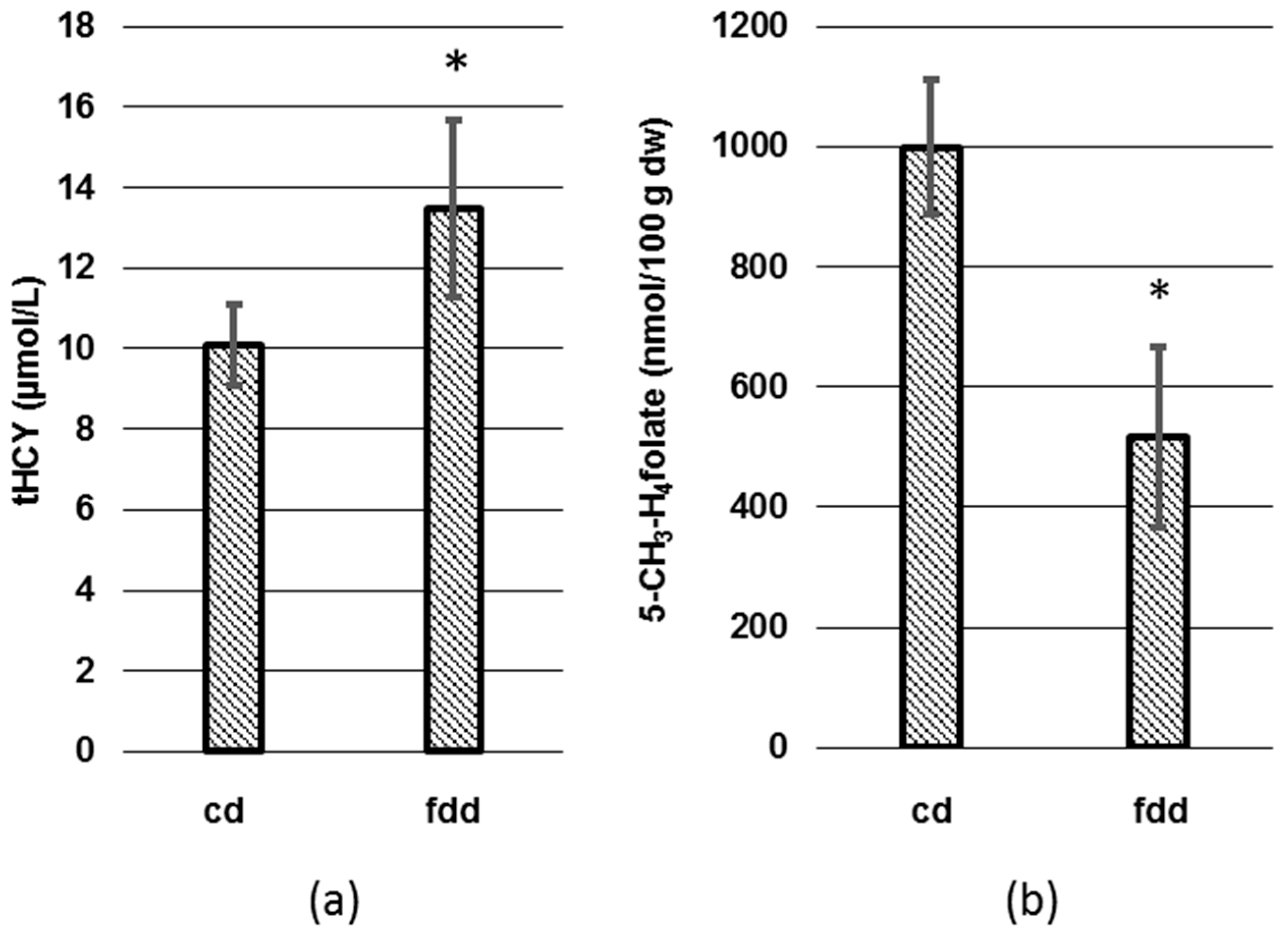
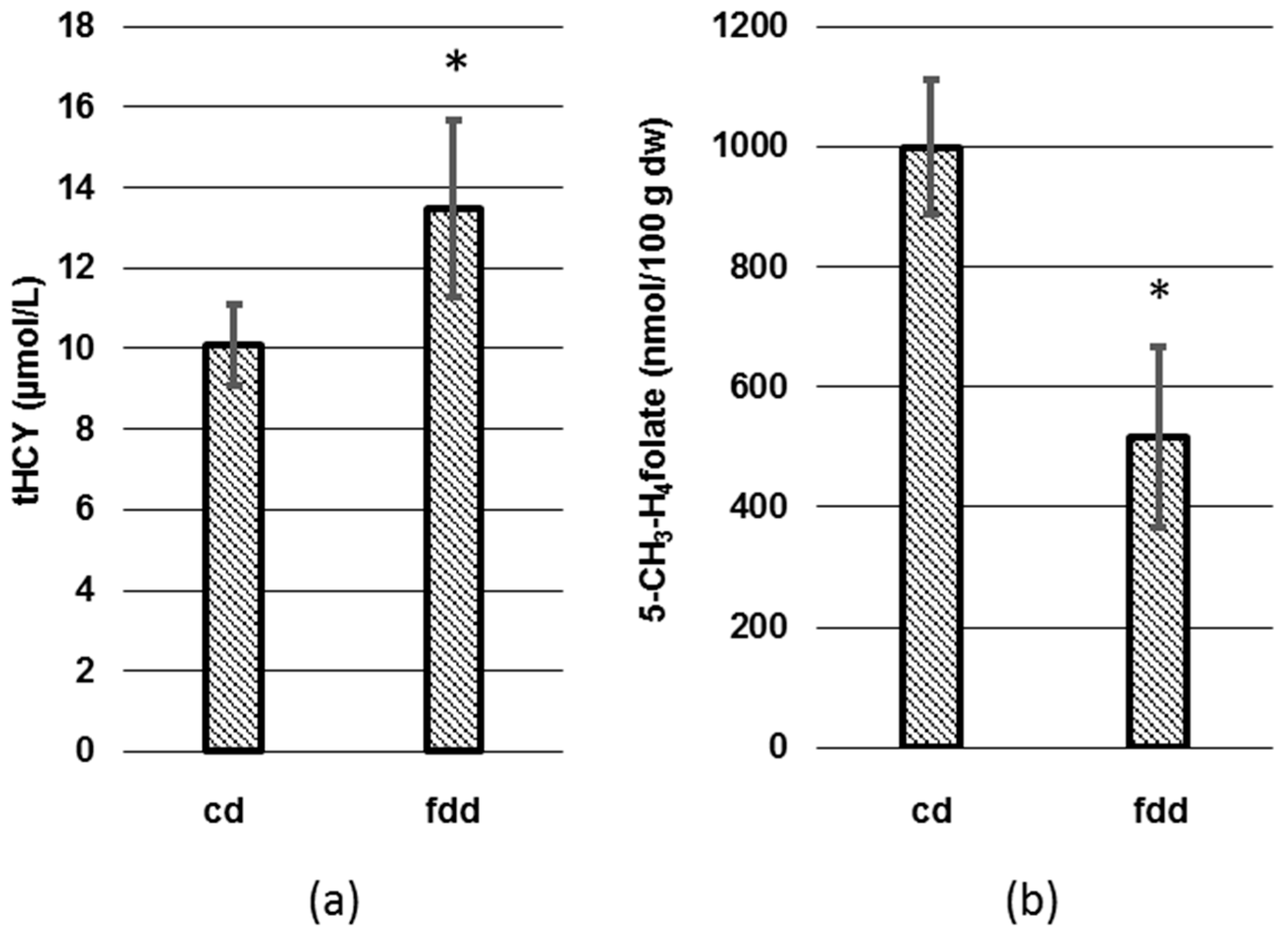
3.2. Analysis of tHcy and Folates in Plasma and Erythrocytes
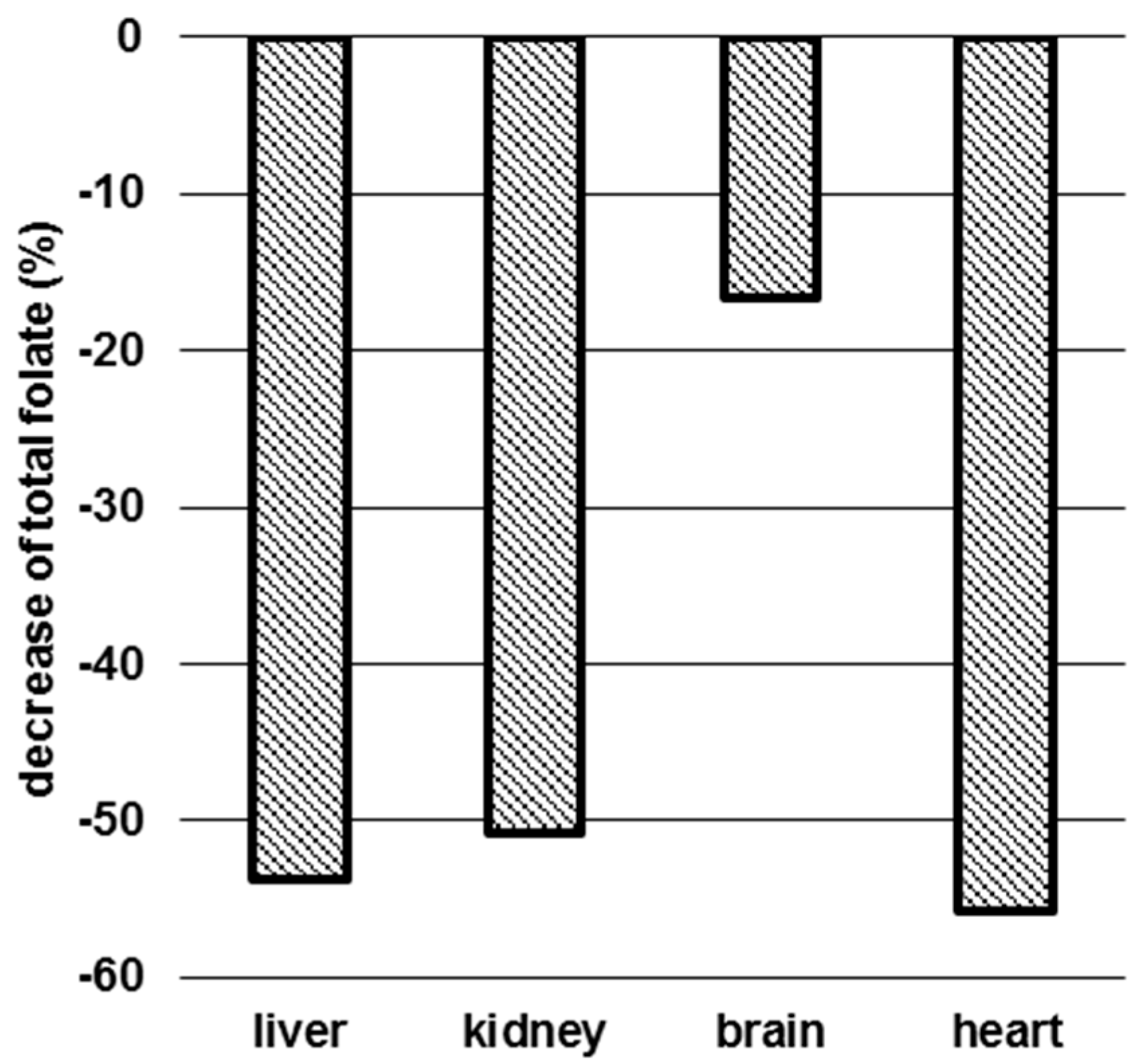
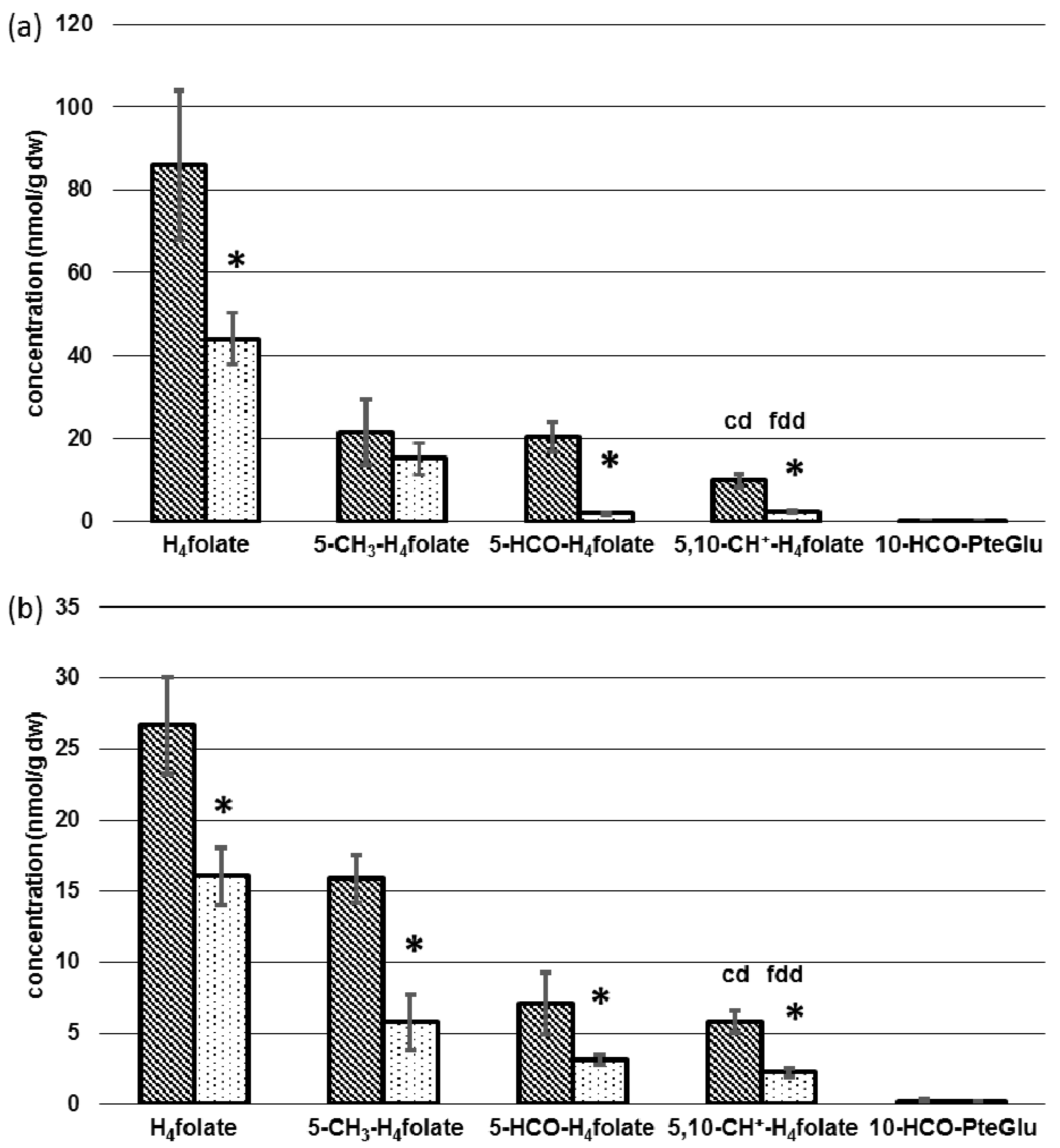
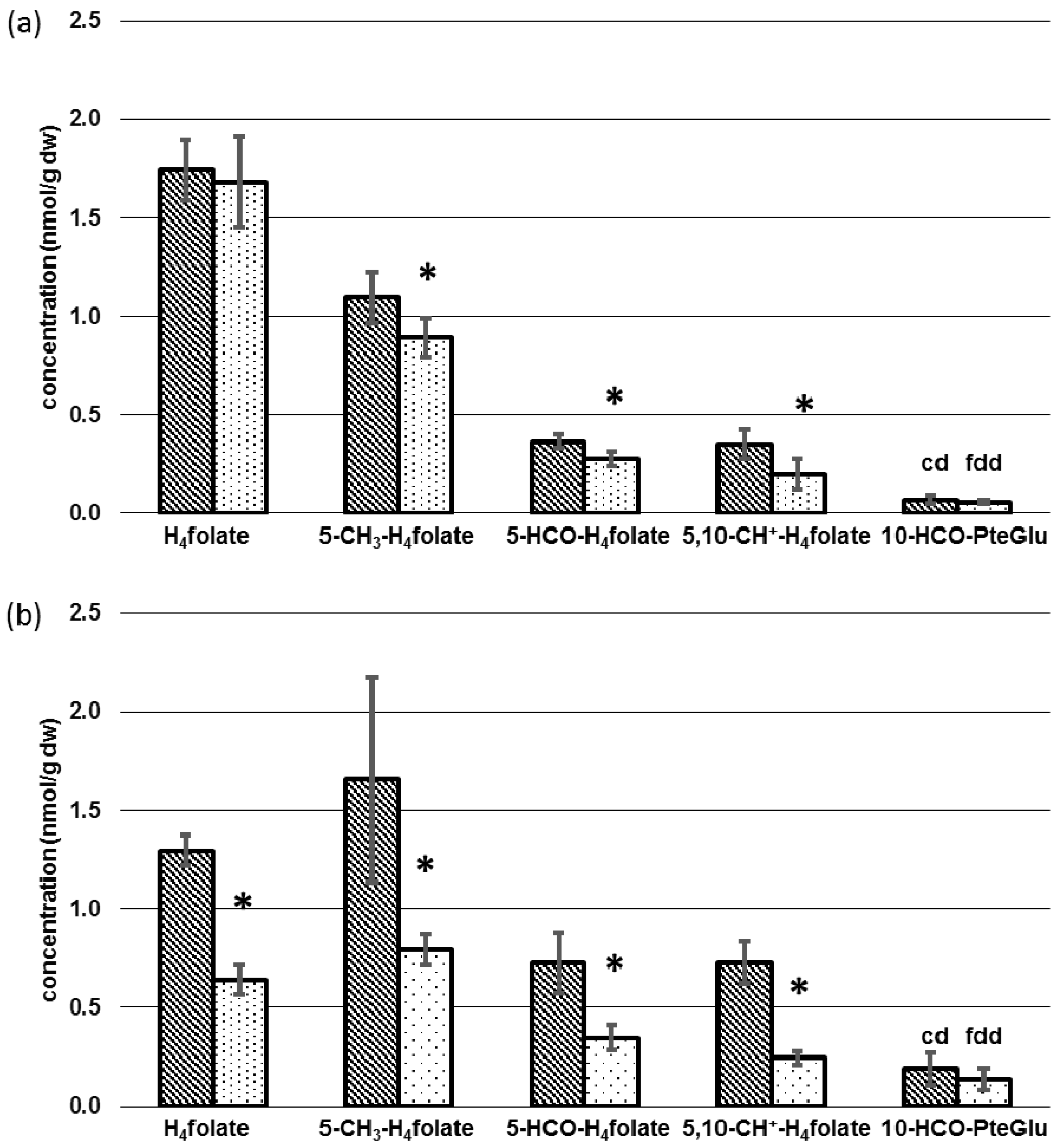
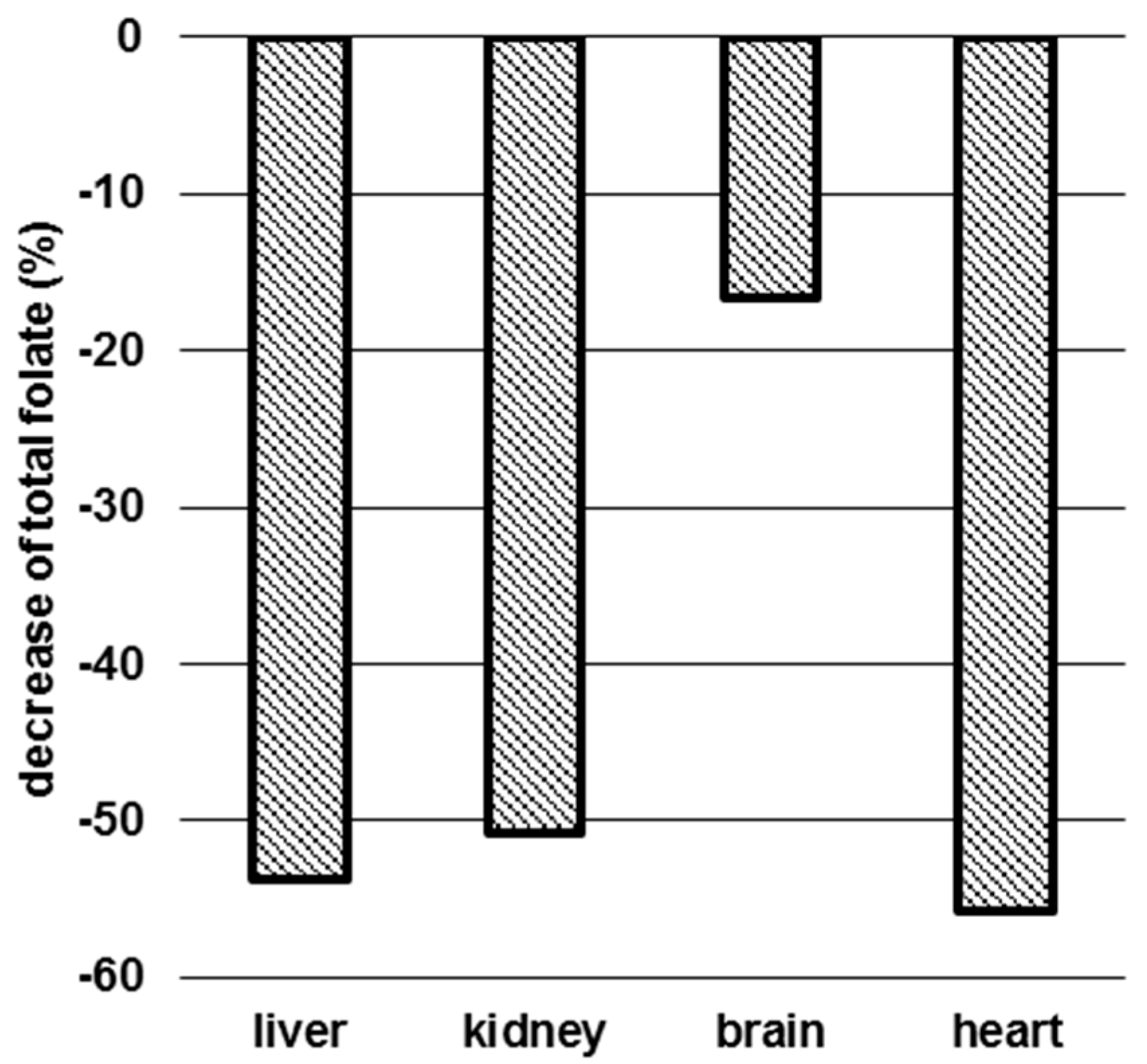
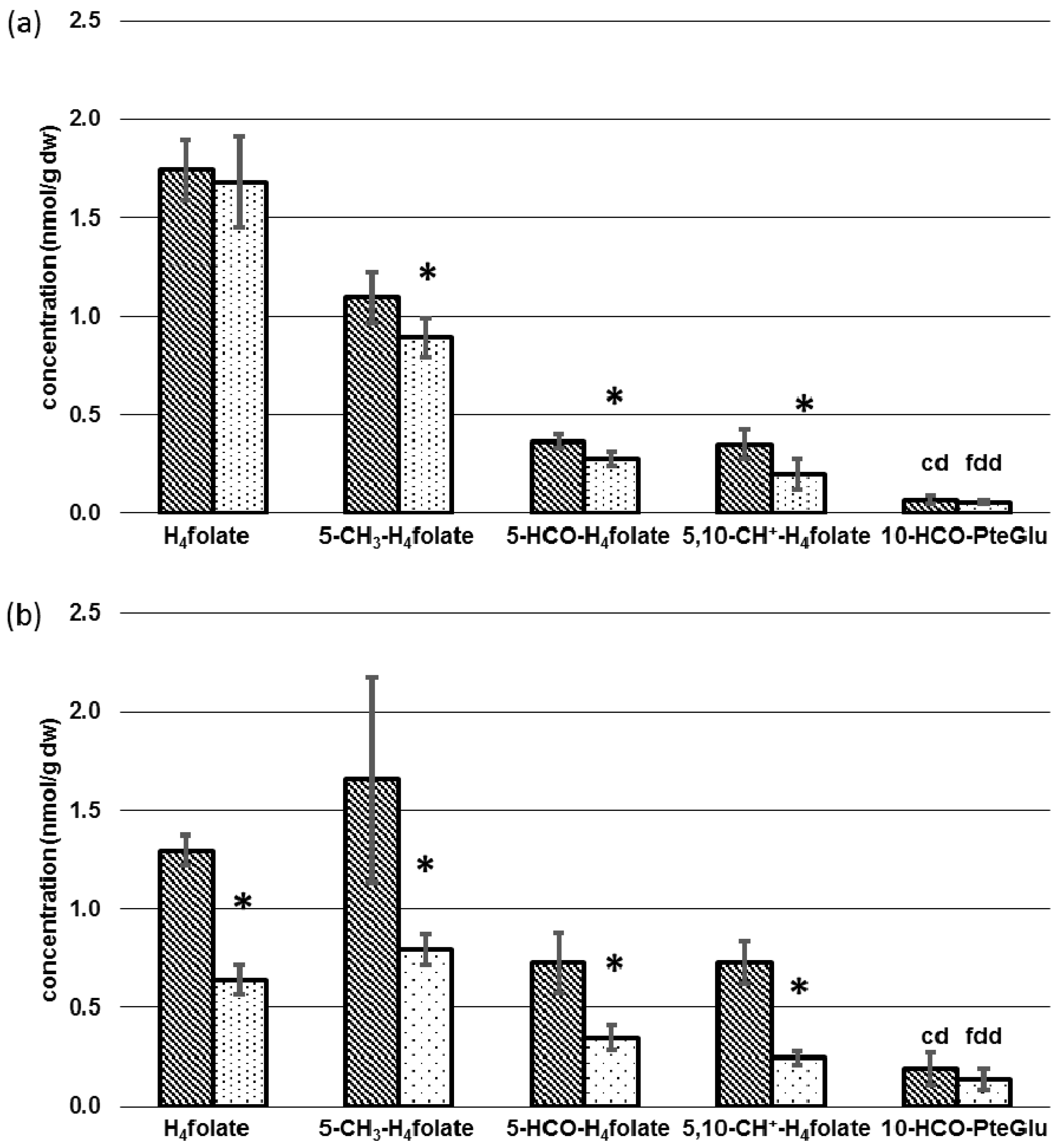
3.3. Determination of Folate Patterns and Total Folate Content in Tissues
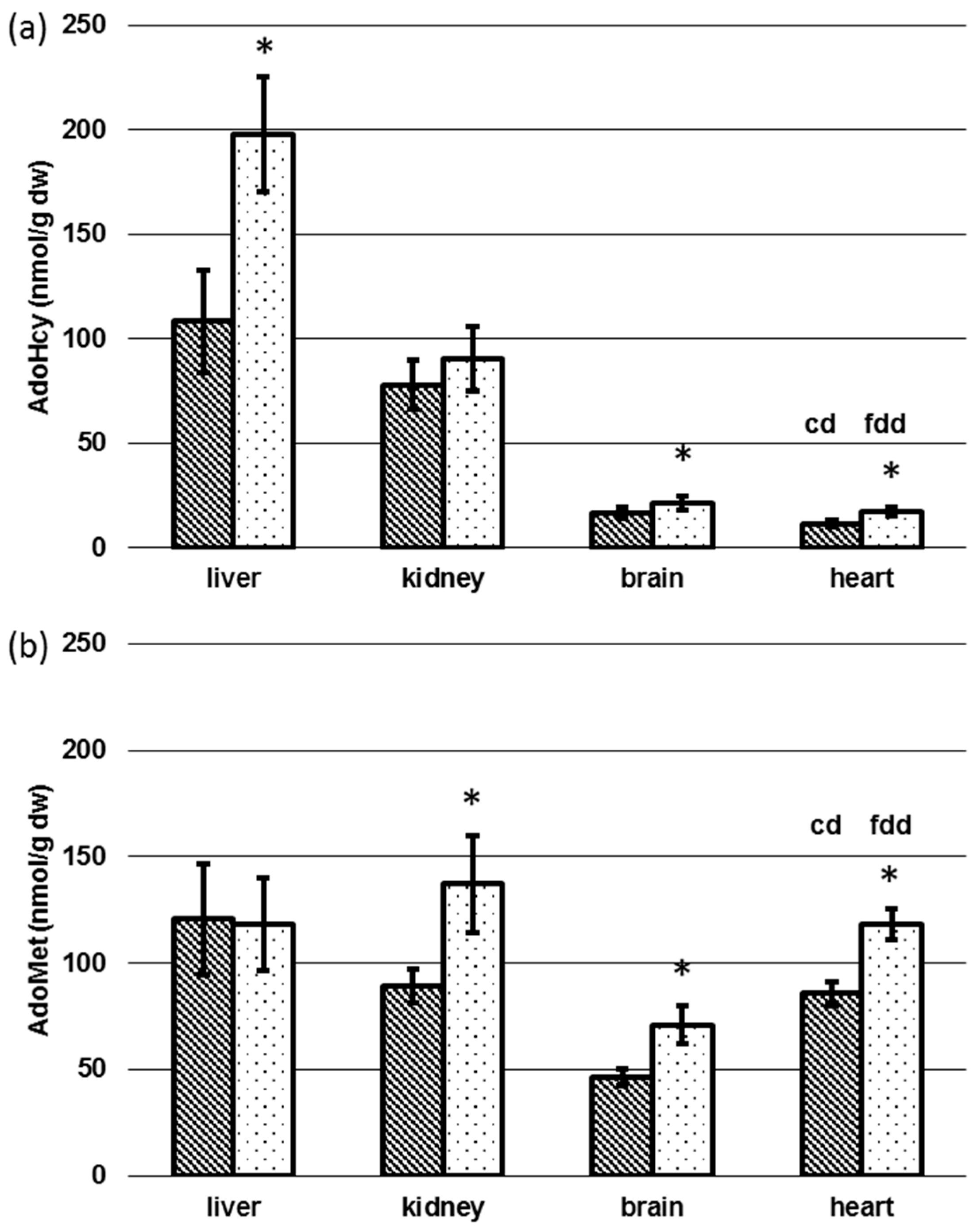
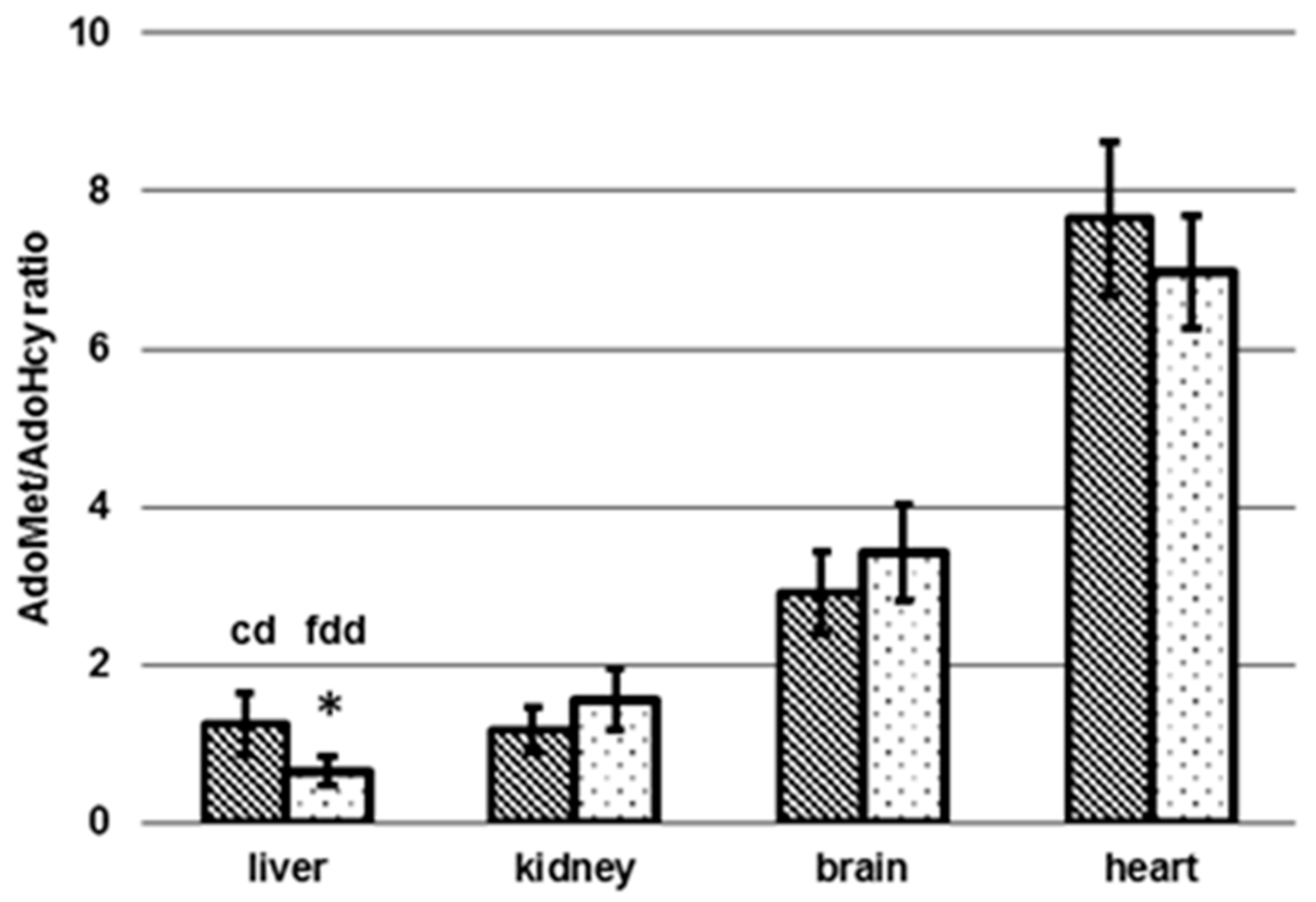
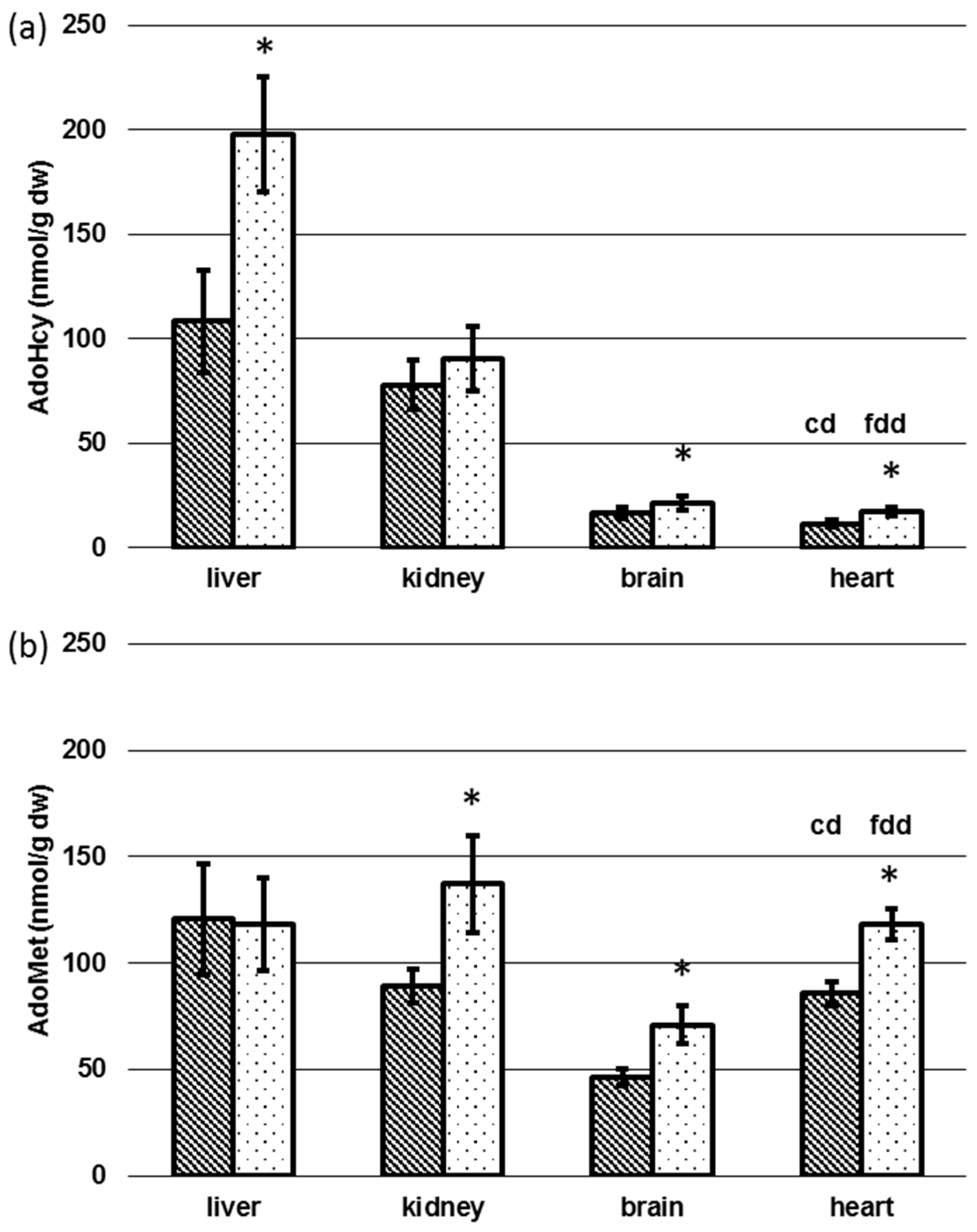
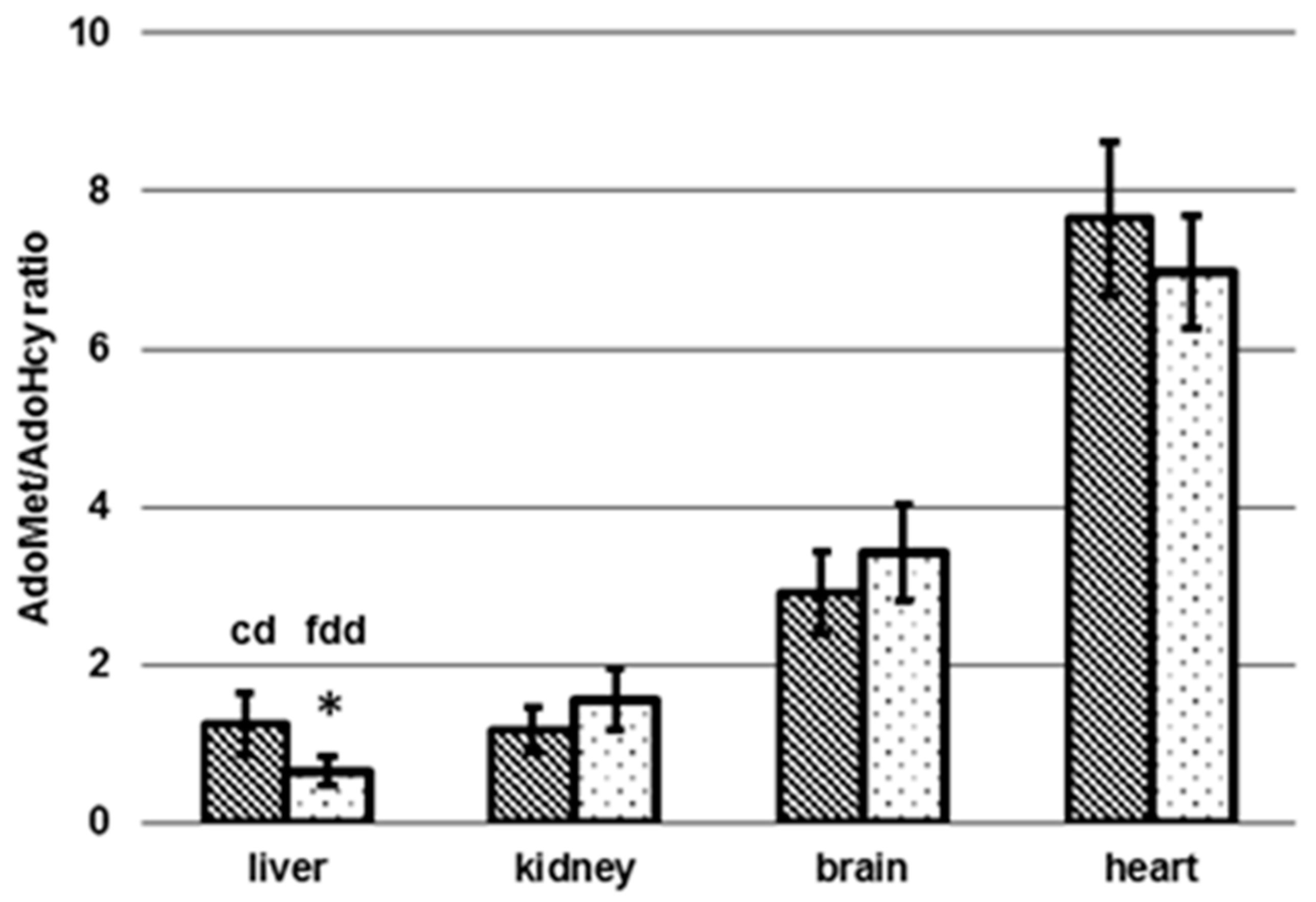
3.4. Determination of AdoMet and AdoHcy in Tissue
4. Discussion
4.1. Body Mass and Erythrocyte Folate Status
4.2. Tissue-Specific Changes of Metabolic Profiles
4.3. Changes of tHcy in Plasma
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Czeizel, A.E.; Dudás, I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N. Engl. J. Med. 1992, 327, 1832–1835. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Pietrzik, K.; Oakley, G.; Kancherla, V.; Holzgreve, W.; Wieser, S. Preventable neural tube defects in Germany. Reprod. Toxicol. 2015, 56, 30–31. [Google Scholar] [CrossRef]
- Robinson, K. Homocysteine, B vitamins, and risk of cardiovascular disease. Heart 2000, 83, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Snowdon, D.A.; Tully, C.L.; Smith, C.D.; Riley, K.P.; Markesbery, W.R. Serum folate and the severity of atrophy of the neocortex in Alzheimer disease: Findings from the Nun study. Am. J. Clin. Nutr. 2000, 71, 993–998. [Google Scholar] [PubMed]
- Hinterberger, M.; Fischer, P. Folate and Alzheimer: When time matters. J. Neural Transm. 2013, 120, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M. The role of folate in reducing chronic and developmental disease risk: An overview. J. Food Sci. 2004, 69, 55–67. [Google Scholar] [CrossRef]
- Choi, S.W.; Mason, J.B. Folate and carcinogenesis: An integrated scheme. J. Nutr. 2000, 130, 129–132. [Google Scholar] [PubMed]
- Nakata, R. Determination of folate derivatives in rat tissues during folate deficiency. J. Nutr. Sci. Vitaminol. 2000, 46, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Varela-Moreiras, G.; Selhub, J. Long-term folate deficiency alters folate content and distribution differentially in rat tissues. J. Nutr. 1992, 122, 986–991. [Google Scholar] [PubMed]
- Ladjimi, H.; Gounelle, J.C.; Auchere, D. Effect of diet on folates levels and distribution in selected tissues of the rat. Arch. Int. Physiol. Biochim. Biophys. 1992, 100, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Clifford, A.J.; Heid, M.K.; Müller, H.G.; Bills, N.D. Tissue distribution and prediction of total body folate of rats. J. Nutr. 1990, 120, 1633–1639. [Google Scholar] [PubMed]
- Pogribny, I.P.; Miller, B.J.; James, S.J. Alterations in hepatic p53 gene methylation patterns during tumor progression with folate/methyl deficiency in the rat. Cancer Lett. 1997, 115, 31–38. [Google Scholar] [CrossRef]
- Caudill, M.A.; Wang, J.C.; Melnyk, S.; Pogribny, I.P.; Jernigan, S.; Collins, M.D.; Santos-Guzman, J.; Swendseid, M.E.; Cogger, E.A.; James, S.J. Intracellular S-adenosylhomocysteine concentrations predict global DNA hypomethylation in tissues of methyl-deficient cystathionine beta-synthase heterozygous mice. J. Nutr. 2001, 131, 2811–2818. [Google Scholar] [PubMed]
- Krijt, J.; Dutá, A.; Kozich, V. Determination of S-adenosylmethionine and S-adenosylhomocysteine by LC-MS/MS and evaluation of their stability in mice tissues. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Dahlhoff, C.; Desmarchelier, C.; Sailer, M.; Fürst, R.W.; Haag, A.; Ulbrich, S.E.; Hummel, B.; Obeid, R.; Geisel, J.; Bader, B.L.; et al. Hepatic methionine homeostasis is conserved in C57BL/6N mice on high-fat diet despite major changes in hepatic one-carbon metabolism. PLoS ONE 2013, 8, e57387. [Google Scholar] [CrossRef]
- Helland, S.; Ueland, P.M. Effect of 2′-deoxycoformycin infusion on S-adenosylhomocysteine hydrolase and the amount of S-adenosylhomocysteine and related compounds in tissues of mice. Cancer Res. 1983, 43, 4142–4147. [Google Scholar] [PubMed]
- Helland, S.; Ueland, P.M. S-adenosylhomocysteine and S-adenosylhomocysteine hydrolase in various tissues of mice given injections of 9-beta-d-arabinofuranosyladenine. Cancer Res. 1983, 43, 1847–1850. [Google Scholar] [PubMed]
- Eloranta, T.O. Tissue distribution of S-adenosylmethionine and S-adenosylhomocysteine in the rat. Effect of age, sex and methionine administration on the metabolism of S-adenosylmethionine, S-adenosylhomocysteine and polyamines. Biochem. J. 1977, 166, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Kopp, M.; Morisset, R.; Koehler, P.; Rychlik, M. Stable isotope dilution assays for clinical analyses of folates and other one-carbon metabolites: Application to folate-deficiency studies. PLoS ONE 2016, 11, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Birn, H. The kidney in vitamin B12 and folate homeostasis: Characterization of receptors for tubular uptake of vitamins and carrier proteins. Am. J. Physiol. Ren. Physiol. 2006, 291, 36. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, N.; Sachdeva, A.; Yadav, S.P.; Choudhury, P. Nutritional Anemia. In Practical Pediatric Hematology: Reference Manual for National Training Project; Sachdeva, A., Yadav, S.P., Arya, S.C., Eds.; Jaypee Brothers Medical Publishers Ltd.: New Delhi, India, 2012; pp. 41–52. [Google Scholar]
- Chen, L.H.; Liu, M.L.; Hwang, H.Y.; Chen, L.S.; Korenberg, J.; Shane, B. Human methionine synthase. cDNA cloning, gene localization, and expression. J. Biol. Chem. 1997, 272, 3628–3634. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.; Poole, J.R. Labile methyl balances for normal humans on various dietary regimens. Metab. Clin. Exp. 1975, 24, 721–735. [Google Scholar] [CrossRef]
- Walker, J.B. Creatine: Biosynthesis, regulation and function. In Advances in Enzymology and Related Areas of Molecular Biology; Meister, A., Ed.; Wiley: New York, NY, USA, 1979; pp. 178–237. [Google Scholar]
- Bottiglieri, T. S-adenosyl-l-methionine (SAMe): From the bench to the bedside—Molecular basis of a pleiotrophic molecule. Am. J. Clin. Nutr. 2002, 76, 1151–1157. [Google Scholar]
- Finkelstein, J.D. Metabolic regulatory properties of S-adenosylmethionine and S-adenosylhomocysteine. Clin. Chem. Lab. Med. 2007, 45, 1694–1699. [Google Scholar] [CrossRef] [PubMed]
- Shane, B. Folate status assessment history: Implications for measurement of biomarkers in NHANES. Am. J. Clin. Nutr. 2011, 94, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Benesch, R.; Waxman, S.; Benesch, R.; Baugh, C. The binding of folyl polyglutamates by hemoglobin. Biochem. Biophys. Res. Commun. 1982, 106, 1359–1363. [Google Scholar] [CrossRef]
- Mönch, S.; Netzel, M.; Netzel, G.; Rychlik, M. Quantitation of folates and their catabolites in blood plasma, erythrocytes, and urine by stable isotope dilution assays. Anal. Biochem. 2010, 398, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Wilcken, D.E.L.; Wilcken, B. Historical Overview and Recent Perspective. In Homocysteine in Health and Disease; Carmel, R., Jacobsen, D.W., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 1–6. [Google Scholar]
- Boushey, C.J.; Beresford, S.A.; Omenn, G.S.; Motulsky, A.G. A quantitative assessment of plasma homocysteine as a risk factor for vascular disease. Probable benefits of increasing folic acid intakes. JAMA 1995, 274, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Refsum, H.; Ueland, P.M.; Nygard, O.; Vollset, S.E. Homocysteine and cardiovascular disease. Annu. Rev. Med. 1998, 49, 31–62. [Google Scholar] [CrossRef] [PubMed]
- Leamon, C.P.; Reddy, J.A.; Dorton, R.; Bloomfield, A.; Emsweller, K.; Parker, N.; Westrick, E. Impact of high and low folate diets on tissue folate receptor levels and antitumor responses toward folate-drug conjugates. J. Pharm. Exp. Ther. 2008, 327, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Chanarin, I.; Slavin, G.; Levi, A.J. Folate deficiency in the alcoholic—Its relationship to clinical and haematological abnormalities, liver disease and folate stores. Br. J. Haematol. 1975, 29, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Qi, H.; Korenberg, J.; Garrow, T.A.; Choi, Y.J.; Shane, B. Purification and properties of human cytosolic folylpoly-gamma-glutamate synthetase and organization, localization, and differential splicing of its gene. J. Biol. Chem. 1996, 271, 13077–13087. [Google Scholar] [PubMed]
- Cook, J.D.; Cichowicz, D.J.; George, S.; Lawler, A.; Shane, B. Mammalian folylpoly-gamma-glutamate synthetase. 4. In vitro and in vivo metabolism of folates and analogues and regulation of folate homeostasis. Biochemistry 1987, 26, 530–539. [Google Scholar] [CrossRef] [PubMed]
- McKeever, M.P.; Weir, D.G.; Molloy, A.; Scott, J.M. Betaine-homocysteine methyltransferase: Organ distribution in man, pig and rat and subcellular distribution in the rat. Clin. Sci. 1991, 81, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, S.E. Mechanisms of folate homeostasis. Am. J. Physiol. 1984, 246, 24. [Google Scholar]
- Pfeiffer, C.; Diehl, J.F.; Schwack, W. Nahrungsfolate—Eine aktuelle Übersicht Stabilität, Physiologische Bedeutung, Bioverfügbarkeit, analytische Bestimmungsmethoden, Einfluß der Lebensmittelbehandlung. Zeitschrift Ernährungswissenschaft 1994, 33, 85–119. [Google Scholar] [CrossRef]
- Finkelstein, J.D. Methionine metabolism in mammals. J. Nutr. Physiol. 1990, 1, 228–237. [Google Scholar] [CrossRef]
- Field, M.S.; Szebenyi, D.M.; Stover, P.J. Regulation of de novo purine biosynthesis by methenyltetrahydrofolate synthetase in neuroblastoma. J. Biol. Chem. 2006, 281, 4215–4221. [Google Scholar] [CrossRef] [PubMed]
- De Brouwer, V.; Zhang, G.-F.; Storozhenko, S.; van der Straeten, D.; Lambert, W.E. pH stability of individual folates during critical sample preparation steps in prevision of the analysis of plant folates. Phytochem. Anal. 2007, 18, 496–508. [Google Scholar] [CrossRef] [PubMed]
- May, M.; Bardos, T.J.; Barger, F.L.; Lansford, M.; Ravel, J.M.; Sutherland, G.L.; Shive, W. Synthetic and degradative investigations of the structure of folinic acid-SF. J. Am. Chem. Soc. 1951, 73, 3067–3075. [Google Scholar] [CrossRef]
- Wilson, S.D.; Horne, D.W. Evaluation of ascorbic acid in protecting labile folic acid derivatives. Proc. Natl. Acad. Sci. USA 1983, 80, 6500–6504. [Google Scholar] [CrossRef] [PubMed]
- Grapp, M.; Wrede, A.; Schweizer, M.; Hüwel, S.; Galla, H.-J.; Snaidero, N.; Simons, M.; Bückers, J.; Low, P.S.; Urlaub, H.; et al. Choroid plexus transcytosis and exosome shuttling deliver folate into brain parenchyma. Nat. Commun. 2013, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fowler, B. Transport and tissue distribution of homocysteine and related S-adenosyl compounds. In Homocysteine in Health and Disease; Carmel, R., Jacobsen, D.W., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2001; pp. 163–175. [Google Scholar]
- Delabar, U.; Kloor, D.; Luippold, G.; Mühlbauer, B. Simultaneous determination of adenosine, S-adenosylhomocysteine and S-adenosylmethionine in biological samples using solid-phase extraction and high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1999, 724, 231–238. [Google Scholar] [PubMed]
- Finkelstein, J.D. The metabolism of homocysteine: Pathways and regulation. Eur. J. Pediatr. 1998, 157, S40–S44. [Google Scholar] [CrossRef] [PubMed]
- Chwatko, G.; Boers, G.H.J.; Strauss, K.A.; Shih, D.M.; Jakubowski, H. Mutations in methylenetetrahydrofolate reductase or cystathionine beta-synthase gene, or a high-methionine diet, increase homocysteine thiolactone levels in humans and mice. FASEB J. 2007, 21, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Hine, R.J.; James, S.J. Increase in plasma homocysteine associated with parallel increases in plasma S-adenosylhomocysteine and lymphocyte DNA hypomethylation. J. Biol. Chem. 2000, 275, 29318–29323. [Google Scholar] [CrossRef] [PubMed]
- Najar, A.; Rahat, B.; Hussain, A.; Thakur, S.; Kaur, J.; Kaur, J.; Hamid, A. Gene specific epigenetic regulation of hepatic folate transport system is responsible for perturbed cellular folate status during aging and exogenous modulation. Mol. Nutr. Food Res. 2016, 60, 1501–1513. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopp, M.; Morisset, R.; Rychlik, M. Characterization and Interrelations of One-Carbon Metabolites in Tissues, Erythrocytes, and Plasma in Mice with Dietary Induced Folate Deficiency. Nutrients 2017, 9, 462. https://doi.org/10.3390/nu9050462
Kopp M, Morisset R, Rychlik M. Characterization and Interrelations of One-Carbon Metabolites in Tissues, Erythrocytes, and Plasma in Mice with Dietary Induced Folate Deficiency. Nutrients. 2017; 9(5):462. https://doi.org/10.3390/nu9050462
Chicago/Turabian StyleKopp, Markus, Rosalie Morisset, and Michael Rychlik. 2017. "Characterization and Interrelations of One-Carbon Metabolites in Tissues, Erythrocytes, and Plasma in Mice with Dietary Induced Folate Deficiency" Nutrients 9, no. 5: 462. https://doi.org/10.3390/nu9050462
APA StyleKopp, M., Morisset, R., & Rychlik, M. (2017). Characterization and Interrelations of One-Carbon Metabolites in Tissues, Erythrocytes, and Plasma in Mice with Dietary Induced Folate Deficiency. Nutrients, 9(5), 462. https://doi.org/10.3390/nu9050462