Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. FA Analyses
2.3. Gene Expression Analysis
2.4. Mineral Analysis
| Element | Minimum Detection Limit |
| 63Cu | 0.63 ng/mL |
| 55Mn | 0.33 ng/mL |
| 66Zn | 2.61 ng/mL |
2.5. Plasma CORT and fT3
2.6. Statistical Analyses
3. Results
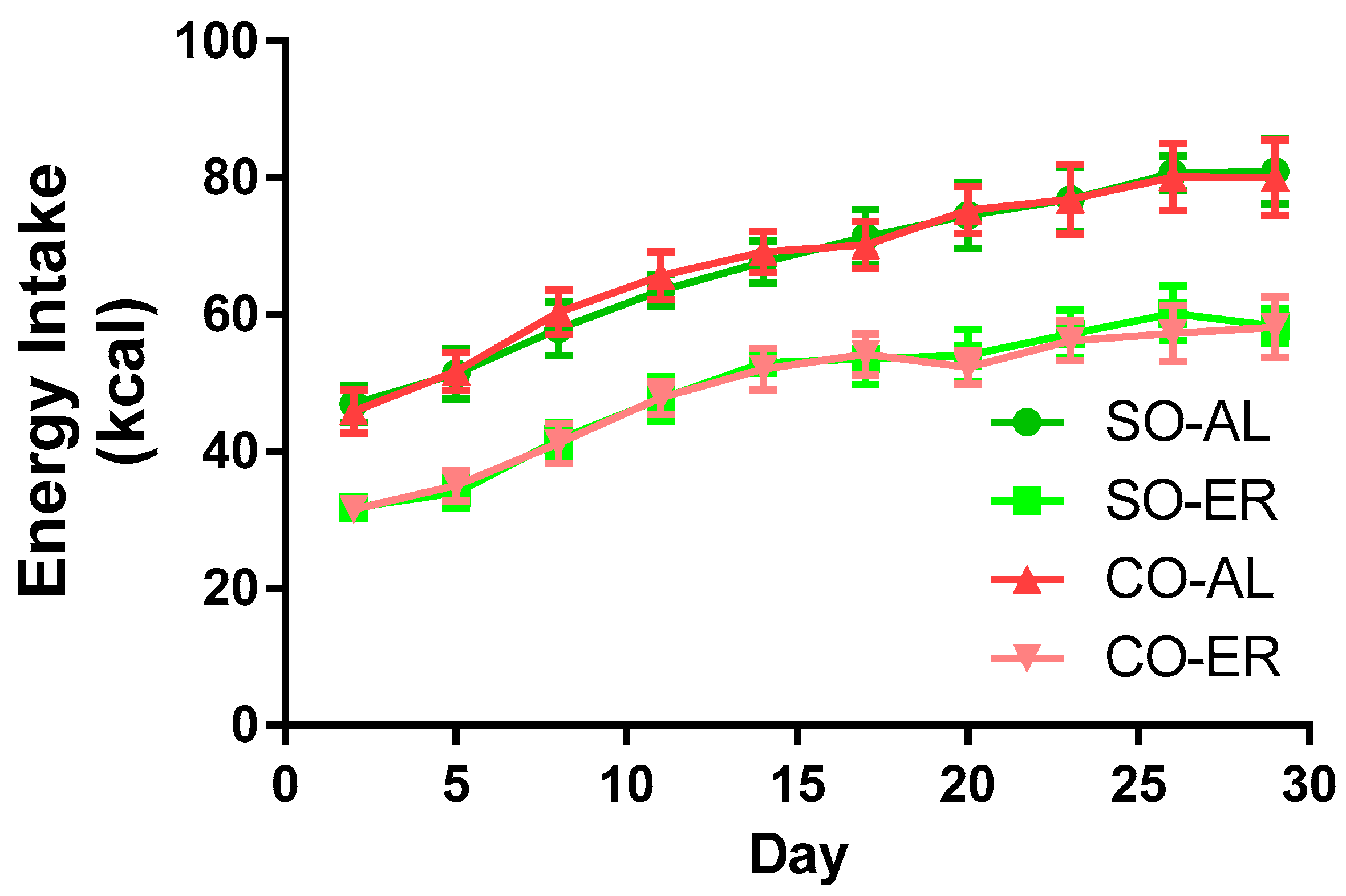
3.1. Energy Intake and Body Composition
3.2. ER and n-3 PUFA-Deficient Diet Effects on Mt1-3 mRNA Expression in the Cerebellum, Cerebral Cortex, Hippocampus, and Liver
3.3. Impact of the Low-n-3 FA Diet and ER on Liver PUFA Content
3.4. Impact of Low-n-3 FA Diet and ER on Cerebellar PUFA
3.5. Impact of Low-n-3 FA Diet and ER on Cerebral Cortex PUFA
3.6. Copper, Manganese, and Zinc Levels in the Cerebellum, Cerebral Cortex, and Liver
3.7. Plasma CORT and fT3 Levels
3.8. ER and n-3 PUFA-Deficient Diet Effects on Glucocorticoid and Mineralocorticoid mRNA Expression in the Cerebellum, Cerebral Cortex, Hippocampus, and Liver
3.9. ER and n-3 PUFA-Deficient Diet Effects on mRNA Expression of TH Regulators in the Cerebellum, Cerebral Cortex, Hippocampus, and Liver
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
| Element | Wavelength | Minimum Detection Limit |
| Ca | 315.887 nm | 101 ppb |
| Cu | 324.752 nm | 10.4 ppb |
| Fe | 259.939 nm | 11 ppb |
| Mg | 279.077 nm | 131 ppb |
| Mn | 257.610 nm | 11.3 ppb |
| P | 214.914 nm | 784 ppb |
| Zn | 213.857 nm | 16 ppb |
| Sc | 361.383 nm | used as the internal standard |
| K | 766.491 nm | 3.328 ppm |
| Na | 588.995 nm | 2.037 ppm |

{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| β-actin | GTG TGG ATT GGT GGC TCT ATC | CAG TCC GCC TAG AAG CAT TT |
| Dio1 | TAC GGG CAA GGT GCT AAT G | GTC TGC TGC CTT GAA TGA AAT C |
| Dio2 | GAA TGC CAC CTT CTT GAC TTT G | CTT GGT TCC GGT GCT TCT TA |
| Dio3 | CGC GAC GTT GAC TTC CTT AT | ATA GGC ACC ATA TGC GGA AC |
| Mct8 | CAG GAG GCA AAC CAG GAA TAT C | TGG CTG CAA ACA CCA CTA TC |
| Mt1 | CCA GAT CTC GGA ATG GAC CCC AAC | GTG CAC TTG TCC GAG GCA CCT TTG |
| Mt2 | GCG ATC TCT CGT TGA TCT CC | CAG GAG CAG GAT CCA TCT GT |
| Mt3 | TGC CCC TGT CCT ACT GGT GGT TCC | CGC CTT TGC AAA CAC AGT CCT TGG |
| Nr3c1 | GGG ACA CGA ATG AGG ATT GTA A | CAG AGG TAC TCA CAC CAT GAA C |
| Nr3c2 | GAG GAC CAC TGA GAA CAA CTA C | GGA CCT GTG ACC ATT CTC TTT |
| Oatp1c1 | GGG CAG GTG TCA GAA AGA TAA | TGA AAG CAT GAG AGT CGT ACA G |
| Thra | CGG TTA TCA CTA CCG CTG TAT C | CAC CCG CTT TGA ATC GTC TA |
| Thrb | GGC ATG TCT CTG TCG TCT TT | CGT CAC CTT CAT CAG GAG TTT |
| ΔCT | ||||
|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER |
| Cerebellum | ||||
| Mt1 | 6.33 ± 0.27 | 6.65 ± 0.33 | 6.38 ± 0.37 | 6.76 ± 0.22 |
| Mt2 | 4.87 ± 0.68 | 5.89 ± 0.36 | 4.51 ± 0.48 | 5.52 ± 0.28 |
| Mt3 | 0.95 ± 0.37 | 1.08 ± 0.19 | 0.95 ± 0.30 | 1.21 ± 0.23 |
| Cerebral Cortex | ||||
| Mt1 | 6.27 ± 0.18 | 6.51 ± 0.36 | 6.58 ± 0.34 | 6.70 ± 0.27 |
| Mt2 | 5.52 ± 0.32 | 5.93 ± 0.31 | 5.78 ± 0.52 | 6.05 ± 0.27 |
| Mt3 | 1.88 ± 0.13 | 1.95 ± 0.14 | 1.98 ± 0.17 | 2.03 ± 0.14 |
| Hippocampus | ||||
| Mt1 | 6.54 ± 0.16 | 6.62 ± 0.22 | 6.50 ± 0.22 | 6.47 ± 0.18 |
| Mt2 | 5.51 ± 0.16 | 5.79 ± 0.27 | 5.50 ± 0.31 | 5.73 ± 0.16 |
| Mt3 | 2.08 ± 0.14 | 2.03 ± 0.15 | 2.17 ± 0.13 | 2.10 ± 0.08 |
| Liver | ||||
| Mt1 | −1.08 ± 0.62 | −0.95 ± 0.73 | 0.19 ± 0.78 | −0.60 ± 0.35 |
| Mt2 | 0.18 ± 0.67 | 0.35 ± 0.91 | 1.64 ± 1.00 | 0.76 ± 0.46 |
| Mt3† | 15.67 ± 0.59 | 15.45 ± 0.80 | 16.05 ± 0.63 | 14.69 ± 1.56 |
| ΔCt | ||||
|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER |
| Cerebellum | ||||
| Nr3c1 | 11.10 ± 0.46 | 11.09 ± 0.51 | 11.49 ± 0.33 | 11.19 ± 0.32 |
| Nr3c2 | 6.81 ± 0.24 | 6.68 ± 0.16 | 6.57 ± 0.21 | 6.62 ± 0.24 |
| Cerebral Cortex | ||||
| Nr3c1 | 9.32 ± 0.49 | 9.29 ± 0.46 | 9.30 ± 0.28 | 9.34 ± 0.38 |
| Nr3c2 | 7.48 ± 0.36 | 7.40 ± 0.30 | 7.33 ± 0.30 | 7.13 ± 0.26 |
| Hippocampus | ||||
| Nr3c1 | 6.31 ± 0.12 | 6.28 ± 0.17 | 6.21 ± 0.17 | 6.42 ± 0.26 |
| Nr3c2 | 6.40 ± 1.40 | 5.92 ± 0.37 | 5.60 ± 0.23 | 5.71 ± 0.15 |
| Liver | ||||
| Nr3c1 | 3.78 ± 0.26 | 2.99 ± 0.49 | 4.05 ± 0.17 | 3.36 ± 0.35 |
| Nr3c2 | 8.54 ± 1.15 | 8.03 ± 0.87 | 8.32 ± 0.72 | 8.00 ± 0.48 |
| ΔCt | ||||
|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER |
| Cerebellum | ||||
| Dio1 | 11.95 ± 0.35 | 11.88 ± 0.42 | 12.02 ± 0.31 | 11.98 ± 0.45 |
| Dio2 | 5.38 ± 0.26 | 5.19 ± 0.19 | 5.41 ± 0.26 | 5.29 ± 0.16 |
| Dio3 | 13.04 ± 0.64 | 12.76 ± 0.28 | 13.32 ± 0.39 | 12.91 ± 0.34 |
| Mct8 | 13.06 ± 0.80 | 13.07 ± 1.08 | 13.34 ± 1.09 | 14.09 ± 0.46 |
| Oatp1c1 | 4.72 ± 0.28 | 4.78 ± 0.25 | 4.92 ± 0.37 | 5.27 ± 0.46 |
| Thra | 4.79 ± 0.09 | 4.73 ± 0.12 | 4.93 ± 0.09 | 4.78 ± 0.15 |
| Thrb | 9.65 ± 0.60 | 9.76 ± 0.18 | 10.10 ± 0.18 | 9.98 ± 0.24 |
| Cerebral Cortex | ||||
| Dio1 | 14.00 ± 1.80 | 13.51 ± 1.21 | 13.19 ± 1.67 | 13.76 ± 0.87 |
| Dio2 | 5.61 ± 0.18 | 5.35 ± 0.24 | 5.15 ± 0.14 | 5.26 ± 0.31 |
| Dio3 | 11.03 ± 0.62 | 11.62 ± 0.67 | 10.95 ± 0.95 | 10.71 ± 1.13 |
| Mct8 | 13.98 ± 0.39 | 13.99 ± 0.30 | 13.83 ± 0.46 | 13.69 ± 0.32 |
| Oatp1c1 | 5.41 ± 0.12 | 5.50 ± 0.27 | 5.44 ± 0.27 | 5.84 ± 0.30 |
| Thra | 3.95 ± 0.09 | 3.84 ± 0.13 | 3.70 ± 0.15 | 3.61 ± 0.18 |
| Thrb | 6.91 ± 0.20 | 6.67 ± 0.26 | 6.62 ± 0.17 | 6.51 ± 0.17 |
| Hippocampus | ||||
| Dio1 | 14.49 ± 1.24 | 13.93 ± 1.30 | 14.16 ± 1.23 | 14.28 ± 1.18 |
| Dio2 | 5.53 ± 0.16 | 5.39 ± 0.18 | 5.50 ± 0.21 | 5.45 ± 0.26 |
| Dio3 | 11.33 ± 0.62 | 11.02 ± 0.76 | 10.85 ± 0.97 | 10.97 ± 0.85 |
| Mct8 | 12.49 ± 0.46 | 12.39 ± 0.52 | 12.32 ± 0.28 | 12.42 ± 0.42 |
| Oatp1c1 | 5.47 ± 0.38 | 5.41 ± 0.31 | 5.42 ± 0.20 | 5.37 ± 0.22 |
| Thra | 2.82 ± 0.24 | 2.75 ± 0.22 | 2.67 ± 0.22 | 2.62 ± 0.19 |
| Thrb | 6.96 ± 0.17 | 6.75 ± 0.29 | 6.84 ± 0.55 | 6.63 ± 0.37 |
| Liver | ||||
| Dio1 | 1.43 ± 0.37 | 0.68 ± 0.33 | 1.75 ± 0.36 | 0.87 ± 0.41 |
| Dio2† | 11.85 ± 0.95 | 13.24 ± 3.13 | 13.27 ± 1.04 | 13.52 ± 1.55 |
| Dio3 | 11.61 ± 0.98 | 12.13 ± 1.60 | 12.72 ± 1.04 | 12.37 ± 1.07 |
| Mct8 | 8.33 ± 0.41 | 8.29 ± 0.36 | 8.77 ± 0.40 | 8.71 ± 0.36 |
| Oatp1c1† | 16.03 ± 1.01 | 15.65 ± 0.91 | 16.43 ± 1.19 | 16.02 ± 0.43 |
| Thra | 6.13 ± 0.18 | 6.92 ± 0.14 | 6.25 ± 0.19 | 6.95 ± 0.24 |
| Thrb | 5.03 ± 0.18 | 4.32 ± 0.16 | 5.31 ± 0.14 | 4.61 ± 0.25 |
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER | pa | pb | pc |
|---|---|---|---|---|---|---|---|
| Cerebellum | |||||||
| β-actin | 14.92 ± 0.18 | 14.81 ± 0.12 | 14.81 ± 0.29 | 14.59 ± 0.20 | 0.73 | 0.15 | 0.75 |
| Cerebral Cortex | |||||||
| β-actin | 14.11 ± 0.27 | 14.00 ± 0.12 | 14.21 ± 0.24 | 14.20 ± 0.21 | 0.72 | >0.99 | 0.82 |
| Hippocampus | |||||||
| β-actin | 14.26 ± 0.22 | 14.13 ± 0.19 | 14.06 ± 0.21 | 14.11 ± 0.19 | 0.57 | 0.95 | 0.20 |
| Liver | |||||||
| β-actin | 16.83 ± 0.33 | 17.15 ± 0.25 | 16.55 ± 0.24 | 16.81 ± 0.25 | 0.11 | 0.22 | 0.17 |
References
- Hulzebos, C.V.; Sauer, P.J. Energy requirements. Semin. Fetal Neonatal Med. 2007, 12, 2–10. [Google Scholar] [PubMed]
- WHO. Global nutrition targets 2025: stunting policy brief. (WHO/NMH/NHD/14.3); World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Cardoso, A.; Marrana, F.; Andrade, J.P. Caloric restriction in young rats disturbs hippocampal neurogenesis and spatial learning. Neurobiol. Learn. Mem. 2016, 133, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Jahng, J.W.; Kim, J.G.; Kim, H.J.; Kim, B.T.; Kang, D.W.; Lee, J.H. Chronic food restriction in young rats results in depression- and anxiety-like behaviors with decreased expression of serotonin reuptake transporter. Brain Res. 2007, 1150, 100–107. [Google Scholar] [PubMed]
- Messaoudi, I.; Fischer, M.; Warner, J.; Park, B.; Mattison, J.; Ingram, D.K.; Totonchy, T.; Mori, M.; Nikolich-Zugich, J. Optimal window of caloric restriction onset limits its beneficial impact on T-cell senescence in primates. Aging Cell 2008, 7, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Remmers, F.; Verhagen, L.A.; Adan, R.A.; Delemarre-van de Waal, H.A. Hypothalamic neuropeptide expression of juvenile and middle-aged rats after early postnatal food restriction. Endocrinology 2008, 149, 3617–3625. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Dietary (n-3) fatty acids and brain development. J. Nutr. 2007, 137, 855–859. [Google Scholar] [PubMed]
- Madore, C.; Nadjar, A.; Delpech, J.C.; Sere, A.; Aubert, A.; Portal, C.; Joffre, C.; Laye, S. Nutritional n-3 PUFAs deficiency during perinatal periods alters brain innate immune system and neuronal plasticity-associated genes. Brain Behav. Immun. 2014, 41, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Ximenes da Silva, A.; Lavialle, F.; Gendrot, G.; Guesnet, P.; Alessandri, J.M.; Lavialle, M. Glucose transport and utilization are altered in the brain of rats deficient in n-3 polyunsaturated fatty acids. J. Neurochem. 2002, 81, 1328–1337. [Google Scholar] [PubMed]
- Picklo, M.J., Sr; Johnson, L.; Idso, J. PPAR mRNA Levels Are Modified by Dietary n-3 Fatty Acid Restriction and Energy Restriction in the Brain and Liver of Growing Rats. J. Nutr. 2017, 147, 161–169. [Google Scholar] [PubMed]
- Aschner, M. The functional significance of brain metallothioneins. FASEB J. 1996, 10, 1129–1136. [Google Scholar] [PubMed]
- Carrasco, J.; Penkowa, M.; Hadberg, H.; Molinero, A.; Hidalgo, J. Enhanced seizures and hippocampal neurodegeneration following kainic acid-induced seizures in metallothionein-I + II-deficient mice. Eur. J. Neurosci. 2000, 12, 2311–2322. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, J.; Belloso, E.; Hernandez, J.; Gasull, T.; Molinero, A. Role of Glucocorticoids on Rat Brain Metallothionein-I and -III Response to Stress. Stress 1997, 1, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, I.; Suzuki, J.S.; Kanazawa, H.; Hara, A.; Saio, M.; Inuzuka, T.; Miyairi, S.; Naganuma, A.; Tohyama, C. Metallothionei n-3 is expressed in the brain and various peripheral organs of the rat. Neurosci. Lett. 2008, 438, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Szrok, S.; Stelmanska, E.; Turyn, J.; Bielicka-Gieldon, A.; Sledzinski, T.; Swierczynski, J. Metallothioneins 1 and 2, but not 3, are regulated by nutritional status in rat white adipose tissue. Genes Nutr. 2016, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system, Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Yeiser, E.C.; Fitch, C.A.; Horning, M.S.; Rutkoski, N.; Levenson, C.W. Regulation of metallothionei n-3 mRNA by thyroid hormone in developing rat brain and primary cultures of rat astrocytes and neurons. Dev. Brain Res. 1999, 115, 195–200. [Google Scholar] [CrossRef]
- Swindell, W.R. Comparative analysis of microarray data identifies common responses to caloric restriction among mouse tissues. Mech. Ageing Dev. 2008, 129, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R. Genes regulated by caloric restriction have unique roles within transcriptional networks. Mech. Ageing Dev. 2008, 129, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R. Genes and gene expression modules associated with caloric restriction and aging in the laboratory mouse. BMC Genomics 2009, 10, 585. [Google Scholar] [CrossRef] [PubMed]
- Olivo-Marston, S.E.; Zhu, Y.; Lee, R.Y.; Cabanes, A.; Khan, G.; Zwart, A.; Wang, Y.; Clarke, R.; Hilakivi-Clarke, L. Gene signaling pathways mediating the opposite effects of prepubertal low-fat and high-fat n-3 polyunsaturated fatty acid diets on mammary cancer risk. Cancer Prev. Res. 2008, 1, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.H.; Wood, A.M.; Newman, A.M.; Bremner, I.; Choo, K.H.; Michalska, A.E.; Duncan, J.S.; Trayhurn, P. Obesity and hyperleptinemia in metallothionein (-I and -II) null mice. Proc. Natl. Acad. Sci. USA 1998, 95, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Li, Q.; Sreejayan, N.; Nunn, J.M.; Ren, J. Metallothionein prevents high-fat diet induced cardiac contractile dysfunction, role of peroxisome proliferator activated receptor gamma coactivator 1alpha and mitochondrial biogenesis. Diabetes 2007, 56, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kawakami, T.; Kondoh, M.; Takiguchi, M.; Kadota, Y.; Himeno, S.; Suzuki, S. Development of high-fat-diet-induced obesity in female metallothionein-null mice. FASEB J. 2010, 24, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Lindeque, J.Z.; Jansen van Rensburg, P.J.; Louw, R.; van der Westhuizen, F.H.; Florit, S.; Ramirez, L.; Giralt, M.; Hidalgo, J. Obesity and metabolomics, metallothioneins protect against high-fat diet-induced consequences in metallothionein knockout mice. Omics 2015, 19, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Herlihy, J.T.; Stacy, C.; Bertrand, H.A. Long-term food restriction depresses serum thyroid hormone concentrations in the rat. Mech. Ageing Dev. 1990, 53, 9–16. [Google Scholar] [CrossRef]
- Levay, E.A.; Tammer, A.H.; Penman, J.; Kent, S.; Paolini, A.G. Calorie restriction at increasing levels leads to augmented concentrations of corticosterone and decreasing concentrations of testosterone in rats. Nutr. Res. 2010, 30, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.J.; Sandgren, E.P.; Brinster, R.L.; Palmiter, R.D. A pair of adjacent glucocorticoid response elements regulate expression of two mouse metallothionein genes. Proc. Natl. Acad. Sci. USA 1997, 94, 10045–10050. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838s–841s. [Google Scholar] [PubMed]
- Picklo, M.J.; Thyfault, J.P. Vitamin E and vitamin C do not reduce insulin sensitivity but inhibit mitochondrial protein expression in exercising obese rats. Appl. Physiol. Nutr. Metab. 2015, 40, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Picklo, M.J.; Murphy, E.J. A High-Fat, High-Oleic Diet, But Not a High-Fat, Saturated Diet, Reduces Hepatic α-Linolenic Acid and Eicosapentaenoic Acid Content in Mice. Lipids 2016, 51, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Thavarajah, D.; Thavarajah, P.; Vial, E.; Gebhardt, M.; Lacher, C.; Kumar, S.; Combs, G.F. Will selenium increase lentil (Lens culinaris Medik) yield and seed quality? Front Plant Sci. 2015, 6, 356. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D.; Whitmore, T.E.; Durnam, D.M. MT-III, a brain-specific member of the metallothionein gene family. Proc. Natl. Acad. Sci. USA 1992, 89, 6333–6337. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, R.A.; Cai, Y.; Shi, Q.; Larson, A.A. The distribution of zinc selenite and expression of metallothionein-III mRNA in the spinal cord and dorsal root ganglia of the rat suggest a role for zinc in sensory transmission. J. Neurosci. 1999, 19, 2288–2300. [Google Scholar] [PubMed]
- Kobayashi, K.; Kuroda, J.; Shibata, N.; Hasegawa, T.; Seko, Y.; Satoh, M.; Tohyama, C.; Takano, H.; Imura, N.; Sakabe, K.; et al. Induction of metallothionein by manganese is completely dependent on interleukin-6 production. J. Pharmacol. Exp. Ther. 2007, 320, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Neville, H.E.; Chase, H.P. Undernutrition and cerebellar development. Exp. Neurol. 1971, 33, 485–497. [Google Scholar] [CrossRef]
- Lesnik, P.G.; Ciesielski, K.T.; Hart, B.L.; Benzel, E.C.; Sanders, J.A. Evidence for cerebellar-frontal subsystem changes in children treated with intrathecal chemotherapy for leukemia, enhanced data analysis using an effect size model. Arch. Neurol. 1998, 55, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Tiemeier, H.; Lenroot, R.K.; Greenstein, D.K.; Tran, L.; Pierson, R.; Giedd, J.N. Cerebellum development during childhood and adolescence, a longitudinal morphometric MRI study. Neuroimage 2010, 49, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Riva, D.; Giorgi, C. The cerebellum contributes to higher functions during development, evidence from a series of children surgically treated for posterior fossa tumours. Brain 2000, 123 Pt 5, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Bourre, J.M.; Durand, G.; Pascal, G.; Youyou, A. Brain cell and tissue recovery in rats made deficient in n-3 fatty acids by alteration of dietary fat. J. Nutr. 1989, 119, 15–22. [Google Scholar] [PubMed]
- Carrie, I.; Smirnova, M.; Clement, M.; De, J.D.; Frances, H.; Bourre, J.M. Docosahexaenoic acid-rich phospholipid supplementation, effect on behavior, learning ability, and retinal function in control and n-3 polyunsaturated fatty acid deficient old mice. Nutr. Neurosci. 2002, 5, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Doser, T.A.; Fang, C.X.; Nunn, J.M.; Janardhanan, R.; Zhu, M.; Sreejayan, N.; Quinn, M.T.; Ren, J. Metallothionein prolongs survival and antagonizes senescence-associated cardiomyocyte diastolic dysfunction, role of oxidative stress. FASEB J. 2006, 20, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Giralt, M.; Molinero, A.; Carrasco, J.; Hidalgo, J. Effect of dietary zinc deficiency on brain metallothionein-I and -III mRNA levels during stress and inflammation. Neurochem. Int. 2000, 36, 555–562. [Google Scholar] [CrossRef]
- Ghoshal, K.; Wang, Y.; Sheridan, J.F.; Jacob, S.T. Metallothionein induction in response to restraint stress. Transcriptional control, adaptation to stress, and role of glucocorticoid. J. Bio.l Chem. 1998, 273, 27904–27910. [Google Scholar] [CrossRef]
- Hidalgo, J.; Campmany, L.; Marti, O.; Armario, A. Metallothionein-I induction by stress in specific brain areas. Neurochem. Res. 1991, 16, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.T.; Ghoshal, K.; Sheridan, J.F. Induction of metallothionein by stress and its molecular mechanisms. Gene Expr. 1999, 7, 301–310. [Google Scholar] [PubMed]
- Mayo, K.E.; Palmiter, R.D. Glucocorticoid regulation of metallothionein-I mRNA synthesis in cultured mouse cells. J. Bio.l Chem. 1981, 256, 2621–2624. [Google Scholar]
- Gursoy, E.; Cardounel, A.; Hu, Y.; Kalimi, M. Biological effects of long-term caloric restriction, adaptation with simultaneous administration of caloric stress plus repeated immobilization stress in rats. Exp. Biol. Med. 2001, 226, 97–102. [Google Scholar] [CrossRef]
- Koibuchi, N. The role of thyroid hormone on cerebellar development. Cerebellum 2008, 7, 530–533. [Google Scholar] [CrossRef] [PubMed]
- De Vito, P.; Incerpi, S.; Pedersen, J.Z.; Luly, P.; Davis, F.B.; Davis, P.J. Thyroid hormones as modulators of immune activities at the cellular level. Thyroid 2011, 21, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Back, C.M.; Stohr, S.; Schafer, E.A.; Biebermann, H.; Boekhoff, I.; Breit, A.; Gudermann, T.; Buch, T.R. TSH induces metallothionein 1 in thyrocytes via Gq/11- and PKC-dependent signaling. J. Mol. Endocrinol. 2013, 51, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Riol, M.; Martinez-Arconada, M.J.; Alonso, N.; Soldevila, B.; Marchena, D.; Armengol, M.P.; Sanmarti, A.; Pujol-Borrell, R.; Martinez-Caceres, E.M. Overexpression of metallothionein I/II, a new feature of thyroid follicular cells in Graves’ disease. J. Clin. Endocrinol. Metab. 2012, 97, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, E.; Redman, L.M.; Rochon, J.; Das, S.K.; Fontana, L.; Kraus, W.E.; Romashkan, S.; Williamson, D.A.; Meydani, S.N.; Villareal, D.T.; et al. A 2-Year Randomized Controlled Trial of Human Caloric Restriction, Feasibility and Effects on Predictors of Health Span and Longevity. J. Gerontol. A-Biol. Sci. Med. Sci. 2015, 70, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.L.; de Andrade, B.M.; de Figueiredo, A.S.; da Silva, M.L.; Marassi, M.P.; Pereira Vdos, S.; Bouskela, E.; Carvalho, D.P. Low replacement doses of thyroxine during food restriction restores type 1 deiodinase activity in rats and promotes body protein loss. J. Endocrinol. 2008, 198, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.X.; Dhahbi, J.M.; Mote, P.L.; Spindler, S.R. Genomic profiling of short- and long-term caloric restriction effects in the liver of aging mice. Proc. Natl. Acad. Sci. USA 2001, 98, 10630–10635. [Google Scholar] [CrossRef] [PubMed]
- Heilbronn, L.K.; Ravussin, E. Calorie restriction and aging, review of the literature and implications for studies in humans. Am. J. Clin. Nutr. 2003, 78, 361–369. [Google Scholar] [PubMed]
- Long, E.K.; Picklo, M.J., Sr. Trans-4-hydroxy-2-hexenal, a product of n-3 fatty acid peroxidation, make some room HNE. Free Radic. Biol. Med. 2010, 49, 1–8. [Google Scholar] [PubMed]
- Gao, L.; Wang, J.; Sekhar, K.R.; Yin, H.; Yared, N.F.; Schneider, S.N.; Sasi, S.; Dalton, T.P.; Anderson, M.E.; Chan, J.Y.; et al. Novel n-3 fatty acid oxidation products activate Nrf2 by destabilizing the association between Keap1 and Cullin3. J. Biol. Chem. 2007, 282, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Van Lookeren Campagne, M.; Thibodeaux, H.; van Bruggen, N.; Cairns, B.; Lowe, D.G. Increased binding activity at an antioxidant-responsive element in the metallothionein-1 promoter and rapid induction of metallothionein-1 and -2 in response to cerebral ischemia and reperfusion. J. Neurosci. 2000, 20, 5200–5207. [Google Scholar] [PubMed]
- Ohtsuji, M.; Katsuoka, F.; Kobayashi, A.; Aburatani, H.; Hayes, J.D.; Yamamoto, M. Nrf1 and Nrf2 play distinct roles in activation of antioxidant response element-dependent genes. J. Biol. Chem. 2008, 283, 33554–33562. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Hartley, D.; Klaassen, C.D.; Shehin-Johnson, S.E.; Lucas, A.; Cohen, S.D. Metallothionein-I/II knockout mice are sensitive to acetaminophen-induced hepatotoxicity. J. Pharmacol. Exp. Ther. 1999, 289, 580–586. [Google Scholar] [PubMed]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein Protection of Cadmium Toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Itoh, N.; Takehara, M.; Oguro, I.; Ishizaki, J.I.; Nakanishi, T.; Tanaka, K. Sensitivity of metallothionein-null mice to LPS/d-galactosamine-induced lethality. Biochem. Biophys. Res. Commun. 2001, 280, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Shimada, A.; Satoh, M. Metallothionein as an Anti-Inflammatory Mediator. Mediat. Inflamm. 2009, 2009, 101659. [Google Scholar] [CrossRef] [PubMed]
- Cesani, M.; Cavalca, E.; Macco, R.; Leoncini, G.; Terreni, M.R.; Lorioli, L.; Furlan, R.; Comi, G.; Doglioni, C.; Zacchetti, D.; et al. Metallothioneins as dynamic markers for brain disease in lysosomal disorders. Ann. Neurol. 2014, 75, 127–137. [Google Scholar] [CrossRef] [PubMed]

| p | |||||||
|---|---|---|---|---|---|---|---|
| Endpoint | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| Body Mass (g) | |||||||
| Begin | 54.6 ± 1.7 | 54.7 ± 1.6 | 54.5 ± 1.7 | 54.3 ± 2.4 | 0.92 | 0.72 | 0.83 |
| End | 258.4 ± 9.4 | 197.5 ± 7.5 | 251.0 ± 10.8 | 192.3 ± 10.0 | <0.01 | 0.07 | 0.75 |
| Liver Mass (g) | 7.6 ± 0.4 | 6.5 ± 0.7 | 7.2 ± 0.8 | 6.0 ± 0.7 | <0.01 | 0.07 | 0.71 |
| Brain Mass (g) | 1.6 ± 0.1 | 1.6 ± 0.1 | 1.7 ± 0.1 | 1.6 ± 0.1 | 0.01 | 0.82 | 0.38 |
| Brain/Body (%) | 0.63 ± 0.03 | 0.80 ± 0.04 | 0.66 ± 0.02 | 0.81 ± 0.06 | <0.01 | 0.25 | 0.65 |
| mRNA Fold-Change Compared with SO-AL | p | ||||||
|---|---|---|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| Cerebellum | |||||||
| Mt1 | 1.02 ± 0.18 | 0.82 ± 0.19 | 0.99 ± 0.26 | 0.75 ± 0.12 | <0.01 | 0.44 | 0.79 |
| Mt2 | 1.11 ± 0.61 | 0.50 ± 0.12 | 1.34 ± 0.45 | 0.64 ± 0.11 | <0.01 | 0.04 | 0.97 |
| Mt3 | 1.03 ± 0.29 | 0.92 ± 0.12 | 1.02 ± 0.22 | 0.84 ± 0.14 | 0.06 | 0.48 | 0.51 |
| Cerebral Cortex | |||||||
| Mt1 | 1.00 ± 0.13 | 0.87 ± 0.21 | 0.83 ± 0.21 | 0.76 ± 0.14 | 0.10 | 0.03 | 0.57 |
| Mt2 | 1.02 ± 0.27 | 0.77 ± 0.17 | 0.88 ± 0.32 | 0.70 ± 0.12 | 0.02 | 0.15 | 0.62 |
| Mt3 | 1.00 ± 0.09 | 0.96 ± 0.09 | 0.94 ± 0.11 | 0.91 ± 0.09 | 0.23 | 0.10 | 0.86 |
| Hippocampus | |||||||
| Mt1 | 1.01 ± 0.11 | 0.96 ± 0.15 | 1.04 ± 0.16 | 1.06 ± 0.12 | 0.72 | 0.18 | 0.42 |
| Mt2 | 1.01 ± 0.12 | 0.84 ± 0.16 | 1.03 ± 0.22 | 0.86 ± 0.10 | <0.01 | 0.69 | 0.75 |
| Mt3 | 1.00 ± 0.10 | 1.04 ± 0.11 | 0.94 ± 0.09 | 0.99 ± 0.06 | 0.21 | 0.08 | 0.81 |
| Liver | |||||||
| Mt1 | 1.08 ± 0.40 A | 1.01 ± 0.44 A | 0.47 ± 0.24 B | 0.73 ± 0.18 A,B | 0.16 | <0.01 | 0.05 |
| Mt2 | 1.09 ± 0.48 | 1.05 ± 0.58 | 0.44 ± 0.27 | 0.70 ± 0.22 | 0.21 | <0.01 | 0.07 |
| Mt3 † | - | - | - | - | - | - | - |
| Tissue Concentration, µmol/g | p | ||||||
|---|---|---|---|---|---|---|---|
| FAs | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| n-6 PUFAs | |||||||
| 18:2 | 21.33 ± 4.26 | 18.88 ± 4.14 | 18.29 ± 4.57 | 15.46 ± 3.29 | 0.08 | 0.03 | 0.90 |
| 18:3 | 0.30 ± 0.10 | 0.30 ± 0.07 | 0.26 ± 0.06 | 0.25 ± 0.12 | 0.88 | 0.18 | 0.87 |
| 20:2 | 0.34 ± 0.06 | 0.38 ± 0.08 | 0.34 ± 0.08 | 0.30 ± 0.14 | 0.95 | 0.26 | 0.22 |
| 20:3 | 0.31 ± 0.05 A | 0.63 ± 0.18 B | 0.24 ± 0.06 A | 0.37 ± 0.18 A | <0.01 | <0.01 | 0.04 |
| 20:4 | 21.98 ± 3.32 | 23.11 ± 8.00 | 25.47 ± 5.94 | 23.66 ± 4.41 | 0.87 | 0.32 | 0.47 |
| 22:4 | 0.39 ± 0.08 | 0.39 ± 0.13 | 0.97 ± 0.19 | 0.98 ± 0.19 | 0.92 | <0.01 | 0.92 |
| 22:5 | 0.10 ± 0.11 | 0.19 ± 0.09 | 2.23 ± 0.62 | 2.46 ± 0.57 | 0.29 | <0.01 | 0.67 |
| n-6 LCPUFAs 1 | 23.10 ± 3.53 | 24.70 ± 8.41 | 29.26 ± 6.48 | 27.77 ± 5.23 | 0.98 | 0.04 | 0.49 |
| n-3 PUFAs | |||||||
| 18:3 | 0.87 ± 0.37 A | 0.43 ± 0.15 B | 0.07 ± 0.05 C | 0.02 ± 0.03 C | <0.01 | <0.01 | 0.01 |
| 20:5 | 0.26 ± 0.18 | 0.30 ± 0.06 | BDL | BDL | 0.54 | <0.01 | 0.49 |
| 22:5 | 1.06 ± 0.19 A | 1.42 ± 0.55 A | 0.54 ± 0.13 B | 0.39 ± 0.20 B | 0.34 | <0.01 | 0.03 |
| 22:6 | 6.71 ± 1.04 | 7.11 ± 2.65 | 3.26 ± 0.88 | 2.91 ± 0.46 | 0.97 | <0.01 | 0.49 |
| n-3 LCPUFAs 2 | 8.03 ± 1.23 | 8.83 ± 3.20 | 3.80 ± 1.00 | 3.30 ± 0.65 | 0.82 | <0.01 | 0.32 |
| Tissue Concentration, µmol/g | p | ||||||
|---|---|---|---|---|---|---|---|
| FAs | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| n-6 PUFAs | |||||||
| 18:2 | 1.90 ± 0.14 | 1.81 ± 0.20 | 1.65 ± 0.10 | 2.08 ± 1.49 | 0.53 | 0.97 | 0.34 |
| 20:2 | 0.55 ± 0.07 | 0.53 ± 0.10 | 0.52 ± 0.06 | 0.51 ± 0.05 | 0.63 | 0.30 | 0.89 |
| 20:3 | 0.68 ± 0.05 | 0.65 ± 0.10 | 0.61 ± 0.07 | 0.56 ± 0.03 | 0.08 | <0.01 | 0.71 |
| 20:4 | 9.19 ± 0.58 | 9.06 ± 0.80 | 9.69 ± 0.52 | 9.93 ± 0.72 | 0.81 | <0.01 | 0.44 |
| 22:4 | 2.92 ± 0.22 | 2.84 ± 0.35 | 3.38 ± 0.19 | 3.26 ± 0.18 | 0.27 | <0.01 | 0.80 |
| 22:5 | 0.27 ± 0.11 | 0.35 ± 0.03 | 1.68 ± 0.24 | 1.93 ± 0.20 | <0.01 | <0.01 | 0.16 |
| n-6 LCPUFAs 1 | 13.61 ± 0.78 | 13.43 ± 1.26 | 15.87 ± 0.94 | 16.18 ± 0.79 | 0.84 | <0.01 | 0.48 |
| n-3 PUFAs | |||||||
| 22:5 | 0.27 ± 0.03 | 0.25 ± 0.02 | 0.14 ± 0.02 | 0.15 ± 0.04 | 0.25 | <0.01 | 0.20 |
| 22:6 | 15.11 ± 1.16 | 15.13 ± 1.12 | 13.67 ± 0.70 | 14.07 ± 0.63 | 0.53 | <0.01 | 0.57 |
| n-3 LCPUFAs 2 | 15.38 ± 1.19 | 15.37 ± 1.11 | 13.82 ± 0.71 | 14.22 ± 0.61 | 0.56 | <0.01 | 0.54 |
| Tissue Concentration, µmol/g | p | ||||||
|---|---|---|---|---|---|---|---|
| FAs | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Oil X Energy |
| n-6 PUFAs | |||||||
| 18:2 | 1.23 ± 0.11 | 1.19 ± 0.09 | 1.06 ± 0.07 | 1.01 ± 0.08 | 0.21 | <0.01 | 0.93 |
| 20:2 | 0.18 ± 0.02 | 0.18 ± 0.02 | 0.19 ± 0.02 | 0.19 ± 0.03 | 0.95 | 0.39 | 0.73 |
| 20:3 | 0.41 ± 0.04 | 0.42 ± 0.04 | 0.36 ± 0.03 | 0.38 ± 0.05 | 0.28 | <0.01 | 0.93 |
| 20:4 | 12.08 ± 0.75 | 12.93 ± 0.59 | 12.77 ± 1.19 | 13.42 ± 0.72 | 0.02 | 0.06 | 0.74 |
| 22:4 | 3.20 ± 0.23 | 3.45 ± 0.09 | 4.01 ± 0.41 | 4.19 ± 0.43 | 0.06 | <0.01 | 0.73 |
| 22:5 | 0.72 ± 0.04 | 0.83 ± 0.09 | 2.37 ± 0.37 | 2.66 ± 0.18 | 0.01 | <0.01 | 0.23 |
| n-6 LCPUFAs 1 | 16.59 ± 1.01 | 17.82 ± 0.63 | 19.69 ± 1.90 | 20.84 ± 1.23 | 0.01 | <0.01 | 0.92 |
| n-3 PUFAs | |||||||
| 22:5 | 0.21 ± 0.03 | 0.23 ± 0.02 | 0.15 ± 0.02 | 0.15 ± 0.02 | 0.25 | <0.01 | 0.40 |
| 22:6 | 14.32 ± 1.29 | 15.59 ± 0.92 | 14.53 ± 1.27 | 15.19 ± 1.50 | 0.04 | 0.84 | 0.50 |
| n-3 LCPUFAs 2 | 14.53 ± 1.31 | 15.82 ± 0.93 | 14.67 ± 1.29 | 15.34 ± 1.50 | 0.04 | 0.71 | 0.49 |
| p | |||||||
|---|---|---|---|---|---|---|---|
| Mineral (µg/g) | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| Cerebellum | |||||||
| Copper | 7.8 ± 3.6 | 9.9 ± 2.7 | 7.7 ± 1.2 | 8.8 ± 1.7 | 0.08 | 0.54 | 0.56 |
| Manganese | 2.6 ± 1.6 | 2.2 ± 0.5 | 2.2 ± 0.3 | 2.0 ± 0.3 | 0.27 | 0.28 | 0.79 |
| Zinc | 34.5 ± 10.7 | 51.9 ± 21.9 | 39.6 ± 6.0 | 39.1 ± 5.7 | 0.07 | 0.40 | 0.06 |
| Cerebral Cortex | |||||||
| Copper | 10.3 ± 5.4 | 10.1 ± 1.5 | 11.6 ± 3.8 | 12.0 ± 3.9 | 0.97 | 0.25 | 0.85 |
| Manganese | 2.2 ± 0.5 | 1.8 ± 0.2 | 2.0 ± 0.4 | 1.9 ± 0.2 | 0.07 | 0.58 | 0.09 |
| Zinc | 48.7 ± 11.2 A | 77.7 ± 11.2 B | 77.2 ± 11.3 B | 73.8 ± 12.5 B | <0.01 | <0.01 | <0.01 |
| Liver | |||||||
| Copper | 15.5 ± 2.2 | 14.9 ± 3.9 | 15.2 ± 2.0 | 17.1 ± 2.0 | 0.50 | 0.31 | 0.19 |
| Manganese | 9.0 ± 0.8 | 8.9 ± 0.9 | 9.7 ± 0.6 | 9.7 ± 0.7 | 0.82 | <0.01 | 0.86 |
| Zinc | 107.7 ± 10.1 | 107.8 ± 13.4 | 113.8 ± 7.3 | 112.2 ± 5.1 | 0.82 | 0.13 | 0.81 |
| p | |||||||
|---|---|---|---|---|---|---|---|
| Analyte | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| CORT (ng/mL) | 51.6 ± 25.7 | 125.6 ± 42.7 | 89.9 ± 34.2 | 132.3 ± 79.6 | <0.01 | 0.21 | 0.38 |
| fT3 (pg/mL) | 2.5 ± 0.3 | 2.9 ± 0.4 | 2.8 ± 0.3 | 2.7 ± 0.3 | 0.16 | 0.51 | 0.06 |
| mRNA Fold-Change Compared with SO-AL | p | ||||||
|---|---|---|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| Cerebellum | |||||||
| Nr3c1 | 1.05 ± 0.33 | 1.07 ± 0.34 | 0.78 ± 0.16 | 0.96 ± 0.21 | 0.29 | 0.10 | 0.33 |
| Nr3c2 | 1.01 ± 0.17 | 1.10 ± 0.12 | 1.19 ± 0.18 | 1.15 ± 0.19 | 0.58 | 0.07 | 0.26 |
| Cerebral Cortex | |||||||
| Nr3c1 | 1.06 ± 0.41 | 1.07 ± 0.36 | 1.03 ± 0.21 | 1.02 ± 0.26 | 0.96 | 0.92 | 0.80 |
| Nr3c2 | 1.03 ± 0.24 | 1.07 ± 0.25 | 1.13 ± 0.24 | 1.29 ± 0.23 | 0.22 | 0.06 | 0.57 |
| Hippocampus | |||||||
| Nr3c1 | 1.00 ± 0.08 | 1.02 ± 0.12 | 1.00 ± 0.14 | 0.94 ± 0.17 | 0.17 | 0.81 | 0.08 |
| Nr3c2 | 1.30 ± 0.66 | 1.43 ± 0.34 | 1.75 ± 0.24 | 1.61 ± 0.17 | 0.49 | 0.06 | 0.27 |
| Liver | |||||||
| Nr3c1 | 1.02 ± 0.20 | 1.81 ± 0.65 | 0.84 ± 0.10 | 1.36 ± 0.30 | <0.01 | 0.01 | 0.68 |
| Nr3c2 | 1.25 ± 0.76 | 1.59 ± 0.54 | 1.28 ± 0.52 | 1.53 ± 0.50 | 0.17 | 0.67 | 0.76 |
| mRNA Fold-Change Compared with SO-AL | p | ||||||
|---|---|---|---|---|---|---|---|
| Tissue/Gene | SO-AL | SO-ER | CO-AL | CO-ER | Energy | Oil | Energy X Oil |
| Cerebellum | |||||||
| Dio1 | 1.03 ± 0.26 | 1.09 ± 0.30 | 0.97 ± 0.23 | 1.02 ± 0.27 | 0.70 | 0.54 | 0.91 |
| Dio2 | 1.02 ± 0.20 | 1.15 ± 0.15 | 1.00 ± 0.18 | 1.07 ± 0.12 | 0.05 | 0.41 | 0.62 |
| Dio3 | 1.08 ± 0.43 | 1.23 ± 0.23 | 0.85 ± 0.20 | 1.12 ± 0.32 | 0.04 | 0.18 | 0.68 |
| Mct8 | 1.16 ± 0.76 | 1.31 ± 1.25 | 1.02 ± 0.60 | 0.52 ± 0.16 | 0.24 | 0.05 | 0.25 |
| Oatp1c1 | 1.02 ± 0.18 | 0.98 ± 0.18 | 0.90 ± 0.20 | 0.71 ± 0.19 | 0.11 | <0.01 | 0.25 |
| Thra | 1.00 ± 0.06 | 1.04 ± 0.09 | 0.90 ± 0.05 | 1.01 ± 0.11 | 0.02 | 0.03 | 0.23 |
| Thrb | 1.10 ± 0.64 | 0.94 ± 0.13 | 0.77 ± 0.10 | 0.81 ± 0.14 | 0.87 | 0.02 | 0.48 |
| Cerebral Cortex | |||||||
| Dio1 | 1.61 ± 1.45 | 1.85 ± 1.31 | 3.20 ± 3.70 | 1.39 ± 0.92 | 0.93 | 0.59 | 0.31 |
| Dio2 | 1.01 ± 0.12A | 1.21 ± 0.21A,B | 1.39 ± 0.15B | 1.30 ± 0.26B | 0.39 | <0.01 | 0.03 |
| Dio3 | 1.07 ± 0.37 | 0.75 ± 0.50 | 1.33 ± 1.18 | 1.56 ± 0.92 | 0.58 | 0.12 | 0.19 |
| Mct8 | 1.03 ± 0.30 | 1.01 ± 0.23 | 1.16 ± 0.38 | 1.25 ± 0.28 | 0.63 | 0.10 | 0.57 |
| Oatp1c1 | 1.00 ± 0.09 | 0.96 ± 0.18 | 0.99 ± 0.19 | 0.76 ± 0.15 | 0.01 | 0.05 | 0.10 |
| Thra | 1.00 ± 0.07 | 1.09 ± 0.10 | 1.20 ± 0.13 | 1.28 ± 0.15 | 0.05 | <0.01 | 0.77 |
| Thrb | 1.01 ± 0.14 | 1.20 ± 0.21 | 1.23 ± 0.15 | 1.32 ± 0.16 | 0.02 | <0.01 | 0.33 |
| Hippocampus | |||||||
| Dio1 | 1.51 ± 1.83 | 1.96 ± 1.25 | 1.75 ± 1.51 | 1.55 ± 1.24 | 0.62 | 0.98 | 0.44 |
| Dio2 | 1.01 ± 0.11 | 1.11 ± 0.13 | 1.03 ± 0.15 | 1.07 ± 0.19 | 0.21 | 0.84 | 0.53 |
| Dio3 | 1.08 ± 0.42 | 1.41 ± 0.82 | 1.75 ± 1.42 | 1.49 ± 0.83 | 0.74 | 0.36 | 0.45 |
| Mct8 | 1.05 ± 0.34 | 1.14 ± 0.48 | 1.15 ± 0.24 | 1.09 ± 0.33 | 0.99 | 0.65 | 0.50 |
| Oatp1c1 | 1.03 ± 0.29 | 1.07 ± 0.22 | 1.04 ± 0.14 | 1.08 ± 0.15 | 0.58 | 0.68 | 0.97 |
| Thra | 1.01 ± 0.16 | 1.05 ± 0.15 | 1.12 ± 0.17 | 1.15 ± 0.15 | 0.47 | 0.08 | 0.94 |
| Thrb | 1.01 ± 0.12 | 1.18 ± 0.24 | 1.14 ± 0.32 | 1.29 ± 0.31 | 0.13 | 0.39 | >0.99 |
| Liver | |||||||
| Dio1 | 1.03 ± 0.26 | 1.72 ± 0.41 | 0.82 ± 0.19 | 1.53 ± 0.37 | <0.01 | 0.07 | 0.61 |
| Dio2 † | - | - | - | - | - | - | - |
| Dio3 | 1.19 ± 0.68 | 1.44 ± 2.41 | 0.59 ± 0.46 | 0.78 ± 0.69 | 0.85 | 0.12 | 0.32 |
| Mct8 | 1.04 ± 0.30 | 1.06 ± 0.26 | 0.77 ± 0.22 | 0.79 ± 0.22 | 0.72 | <0.01 | 0.96 |
| Oatp1c1 † | - | - | - | - | - | - | - |
| Thra | 1.01 ± 0.12 | 0.58 ± 0.06 | 0.93 ± 0.12 | 0.57 ± 0.09 | <0.01 | 0.31 | 0.50 |
| Thrb | 1.01 ± 0.13 | 1.64 ± 0.18 | 0.82 ± 0.08 | 1.36 ± 0.23 | <0.01 | <0.01 | 0.99 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehus, A.A.; Picklo Sr, M.J. Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats. Nutrients 2017, 9, 1145. https://doi.org/10.3390/nu9101145
Mehus AA, Picklo Sr MJ. Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats. Nutrients. 2017; 9(10):1145. https://doi.org/10.3390/nu9101145
Chicago/Turabian StyleMehus, Aaron A., and Matthew J. Picklo Sr. 2017. "Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats" Nutrients 9, no. 10: 1145. https://doi.org/10.3390/nu9101145
APA StyleMehus, A. A., & Picklo Sr, M. J. (2017). Brain and Hepatic Mt mRNA Is Reduced in Response to Mild Energy Restriction and n-3 Polyunsaturated Fatty Acid Deficiency in Juvenile Rats. Nutrients, 9(10), 1145. https://doi.org/10.3390/nu9101145