A Survey of Plant Iron Content—A Semi-Systematic Review

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Study Eligibility and Data Extraction
2.3. Statistical Analysis
3. Results
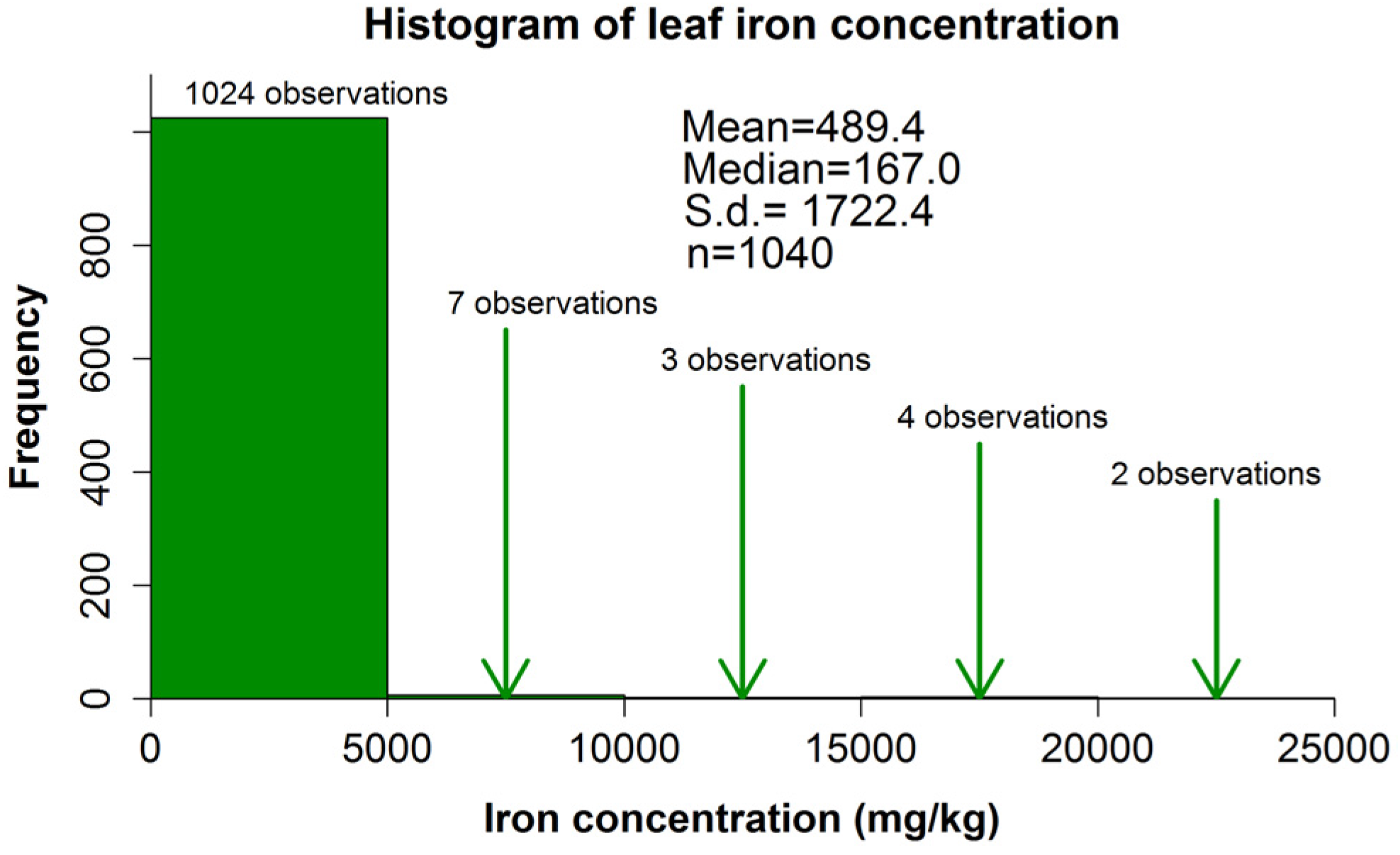
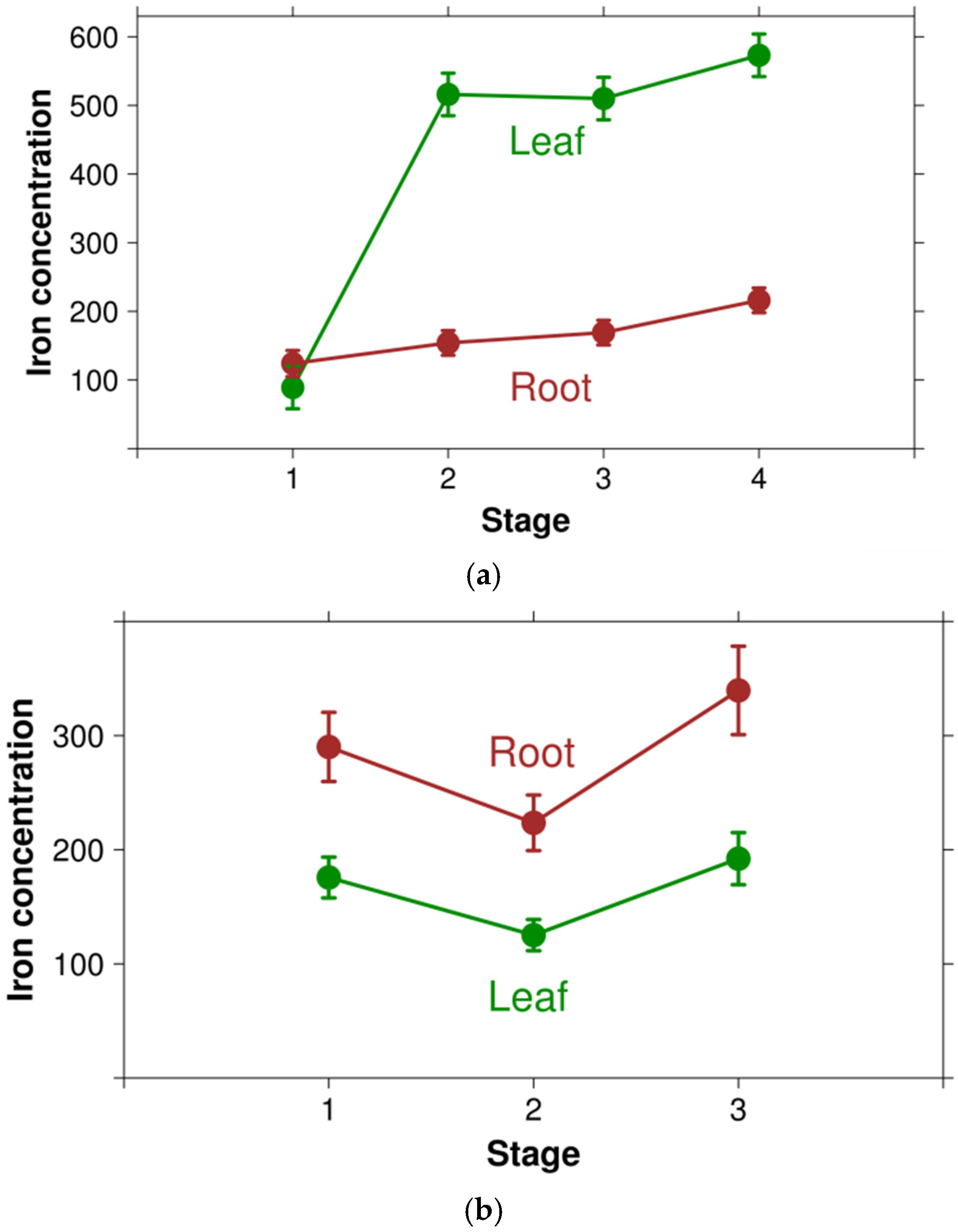
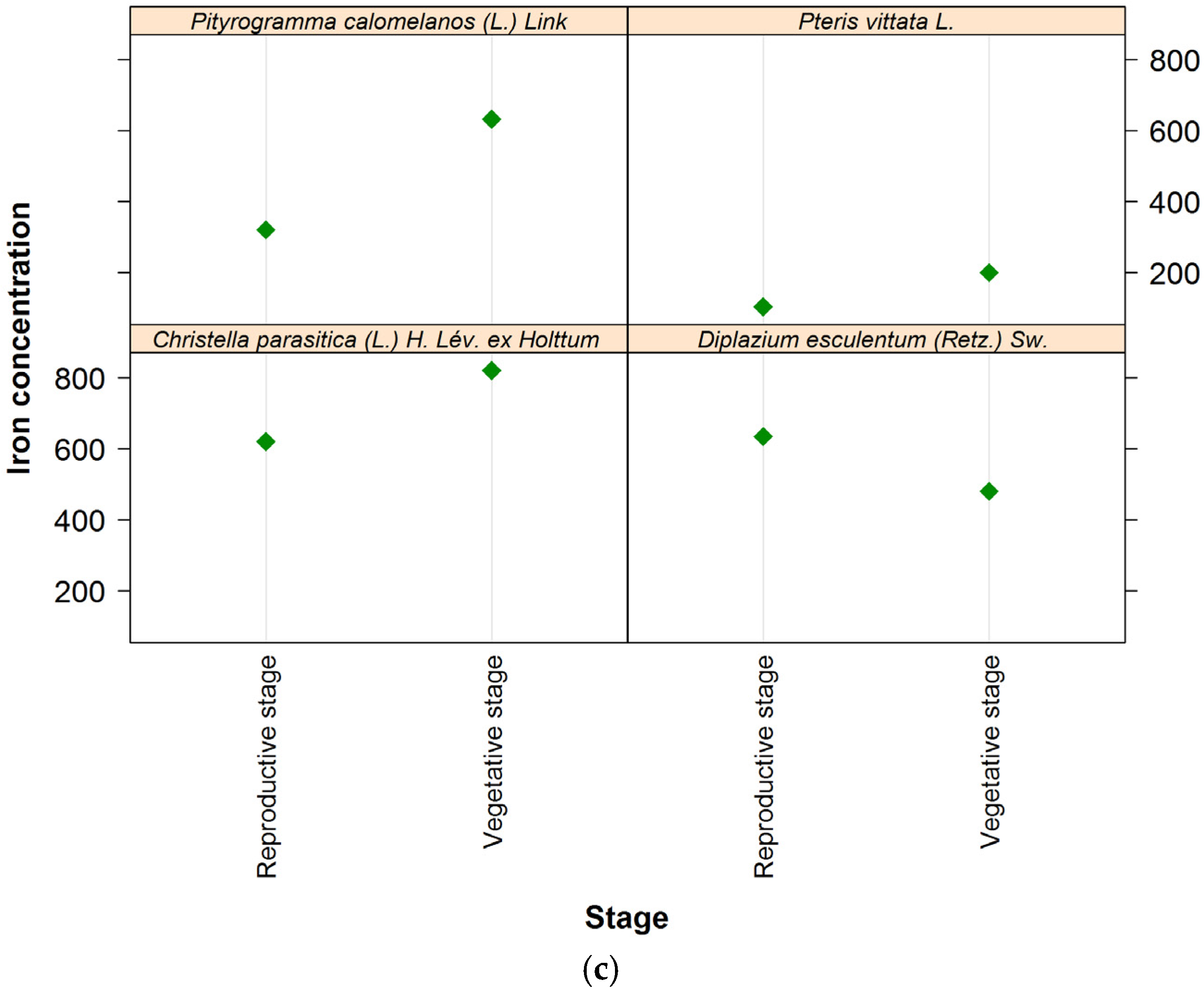
3.1. Extent of Iron Content Investigation in Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Part | Number of Species a | Number of Families | Minimum Iron Conc. (mg/kg, dwb b) | Maximum Iron Conc. (mg/kg, dwb b) | Median Iron Conc. (95% CI) (mg/kg, dwb b) | Mean Iron Conc. (95% CI) (mg/kg, dwb b) |
|---|---|---|---|---|---|---|
| Root | 66 | 33 | 1.9 | 111,200.0 | 502.4 (259.3–691.0) | 5706.0 (2750, 11,560) |
| Stem | 60 | 34 | 7.3 | 25,650.0 | 171.0 (69.2–313.4) | 1431.0 (829, 2696) |
| Shoot | 32 | 22 | 20.2 | 9418.0 | 91.0 (72.7–101.5) | 513.5 (227.1–1113.8) |
| Bark | 41 c | 19 | 3.6 | 1585.0 | 45.0 (35.0–57.0) | 106.8 (74.3–188.8) |
| Leaf | 632 d | 155 | 0.1 | 24,070.0 | 167.0 (155.2–186.6) | 489.4 (401.8–618.4) |
| Aerial parts | 295 | 89 | 0.0 | 27,100.0 | 240.1 (216.5–263.3) | 596.9 (468.4–900.8) |
| Flower | 28 | 15 | 15.7 | 5139.0 | 159.9 (91.2–194.1) | 426.1 (187.5–1008.1) |
| Fruit | 200 e | 62 | 0.0 | 8424.0 | 72.6 (61.0, 87.7) | 257.9 (195.2–393.3) |
| Seed | 104 | 42 | 0.0 | 11,610.0 | 70.2 (53.8–90.0) | 522.6 (333.0–894.4) |
| Whole plant | 41 | 25 | 11.4 | 70,480.0 | 156.0 (89–747) | 2785.0 (1072–9184) |
| Wood | 35 | 15 | 0.0 | 35.0 | 0.0 (N/A) | 3.4 (1.9–6.5) |
| Other parts f | 30 | 28 | 0.7 | 3730.0 | 141.0 (80.0–215.0) | 293.1 (179.3–657.2) |

3.2. Iron Contents in Various Herbal Parts
| Plant Part | Pteridophytes (Median) (95% CI) (n a) | Gymnosperms (Median) (95% CI) (n) | Magnoliids (Median) (95% CI) (n) | Dicots (Median) (95% CI) (n) | Monocots (Median) (95% CI) (n) | Relevant Statistical Comparisons |
|---|---|---|---|---|---|---|
| Root | 296.5 | NA | 259.3 | 426.5 | 573.9 | M b versus D c: p = 0.443 (Mann-Whitney) |
| NA | NA | 194.0–37856.4 | 186.0–985.9 | 394.3–1100.0 | ||
| 2 | 0 | 3 | 44 | 17 | ||
| Stem | 42.0 | 175.65 | 7783.7 | 140.8 | 325.0 | M b versus D c: p = 0.584 (Mann-Whitney) |
| 27–50 | NA | NA | 59–441 | 115.1–550.0 | ||
| 3 | 1 | 1 | 39 | 16 | ||
| Leaf | 200.0 | 133.6 | 253.2 | 163.0 | 188.0 | G d versus D: p = 0.017; |
| 109.5–238.6 | 109.0–155.0 | 166.6–277.5 | 152.0–193.0 | 141.0–240.0 | G versus M: p = 0.038; | |
| 33 | 42 | 34 | 438 | 82 | G: versus Mag e: p = 0.005; * | |
| Shoot | 119.2 | NA | NA | 94.4 | 89.0 | M b versus D c versus P d: p = 0.941 |
| 53.4–128.0 | NA | NA | 52.15–134.50 | 72.7–92.0 | ||
| 4 | 0 | 0 | 20 | 8 | ||
| Aerial parts | 156.0 | 353.5 | 487.0 | 225.0 | 305.0 | M vs. D: p = 0.022 (nparcomp) (Hedges’s g 0.416) |
| 109.0–223.0 | NA | NA | 200.0–243.0 | 220.0–522.0 | ||
| 25 | 2 | 2 | 186 | 64 | ||
| Flower | NA | NA | 2631.45 | 159.9 | NA | NA |
| NA | NA | NA | 88.3–193.6 | NA | ||
| 0 | 0 | 2 | 26 | 0 | ||
| Fruit | NA | NA | 96.42 | 69.9 | 67.8 | Kruskal Wallis: p = 0.486 |
| NA | NA | 77.85–155.00 | 58.00–87.70 | 37.60–186.20 | ||
| 0 | 0 | 9 | 178 | 13 | ||
| Seed | NA | 7.2 | 14.5 | 80.5 | 59 | M vs. D: p = 0.098 (Mann-Whitney) |
| NA | 1.5–41.1 | 0.80–264.60 | 60.0–99.8 | 4.0–70.0 | ||
| 0 | 5 | 5 | 83 | 11 | ||
| Whole plant | 83.0 | 35.2 | NA | 427.0 | 118.0 | M vs. D: p= 0.7976 (Mann-Whitney) |
| 45.0–106.5 | NA | NA | 83.2–1317.5 | 72.5–3041.0 | ||
| 3 | 1 | 0 | 26 | 9 | ||
| Wood | NA | 1.2 | 1.75 | 0 | NA | NA |
| NA | 0.0–3.1 | NA | NA | NA | ||
| 0 | 15 | 2 | 18 | 0 | ||
| Bark | NA | 60.5 | 14.5 | 42.0 | 20.0 | G vs. D: p = 0.089 (Welch t on ranks) |
| NA | 37.0–120.0 | 9.85–123.65 | 24.0–45.7 | NA | ||
| 0 | 15 | 3 | 23 | 1 | ||
| Other parts | 240 | 35.6 | 2065 | 117.9 | 248.0 | NA |
| NA | 15.40, 37.15 | NA | 71.1–190.8 | 45.0–317.0 | ||
| 1 | 2 | 1 | 18 | 8 |
| Plant Part | Herb (Median) (95% CI) (n) | Tree (Median) (95% CI) (n) | Shrub (Median) (95% CI) (n) | Subshrub (Median) (95% CI) (n) | Vine (Median) (95% CI) (n) | Relevant Statistical Comparisons |
|---|---|---|---|---|---|---|
| Root | 506.2 | 226.7 | NA | 559.3 | 1520.0 | NA |
| 288.4–893.6 | 125.0–1401.8 | NA | 137.8–15,777.0 | NA | ||
| 57 | 4 | 0 | 4 | 1 | ||
| Stem | 171.0 | 313.3 | 59.0 | 160.85 | 485.6 | NA |
| 58.0–441.0 | 38.0–313.4 | 37.0–73.0 | NA | 13.6–1300.0 | ||
| 46 | 4 | 4 | 2 | 4 | ||
| Leaf | 200.0 | 149.2 | 162.0 | 263.4 | 397.2 | H a vs. T b: p < 0.0001 (Welch t) (Hedges’s g 0.216) |
| 161.6–218.0 | 134.5–160.0 | 120.0–210.7 | 133.8, 351.0 | 78.8, 532.0 | ||
| 273 | 255 | 73 | 21 | 7 | ||
| Shoot | 91.0 | 49.05 | 108.6 | 6293.9 | 6010.0 | NA |
| 72.7–92.8 | 34.50–82.55 | 28.5–138.4 | NA | NA | ||
| 21 | 4 | 4 | 2 | 1 | ||
| Aerial parts | 240.2 | 220.5 | 259.0 | 266.5 | NA | Kruskal Wallis: p = 0.963 |
| 209.0–274.2 | 200.0–300.0 | 206.0–308.4 | 127.0–458.0 | NA | ||
| 195 | 28 | 52 | 18 | 0 | ||
| Flower | 159.9 | 161.9 | 16.7 | 84.4 | NA | NA |
| 79.7–261.8 | 108.5–204.6 | NA | NA | NA | ||
| 13 | 13 | 1 | 1 | 0 | ||
| Fruit | 100.83 | 68.0 | 50.5 | 152.4 | 134 | H vs. T: p < 0.0001 (Hedges’s g 0.411) |
| 67.0–240.0 | 56.5–81.4 | 40.0–129.1 | NA | 77.5–155.0 | ||
| 24 | 146 | 19 | 1 | 3 | ||
| Seed | 76.0 | 53.2 | 151.9 | 2380 | 11 | H vs. T: p = 0.244 (Welch t on ranks) |
| 59.5–94.9 | 41.1–122.2 | 4.3–4954.0 | NA | NA | ||
| 69 | 27 | 5 | 1 | 2 | ||
| Whole plant | 172 | 29.9 | 520 | NA | NA | NA |
| 85.0–793.0 | NA | 26.5, 1317.5 | NA | NA | ||
| 33 | 2 | 6 | 0 | 0 | ||
| Wood | NA | 0 | NA | NA | NA | NA |
| NA | NA | NA | NA | NA | ||
| 0 | 35 | 0 | 0 | 0 | ||
| Bark | NA | 45.7 | NA | NA | NA | NA |
| NA | 35.1–58.5 | NA | NA | NA | ||
| 0 | 41 | 0 | 0 | 0 | ||
| Other parts | 233.5 | 113.1 | 164.8 | 164.1 | NA | NA |
| 41.0–395.9 | 67.9–190.8 | NA | NA | NA | ||
| 13 | 14 | 2 | 1 | 0 |
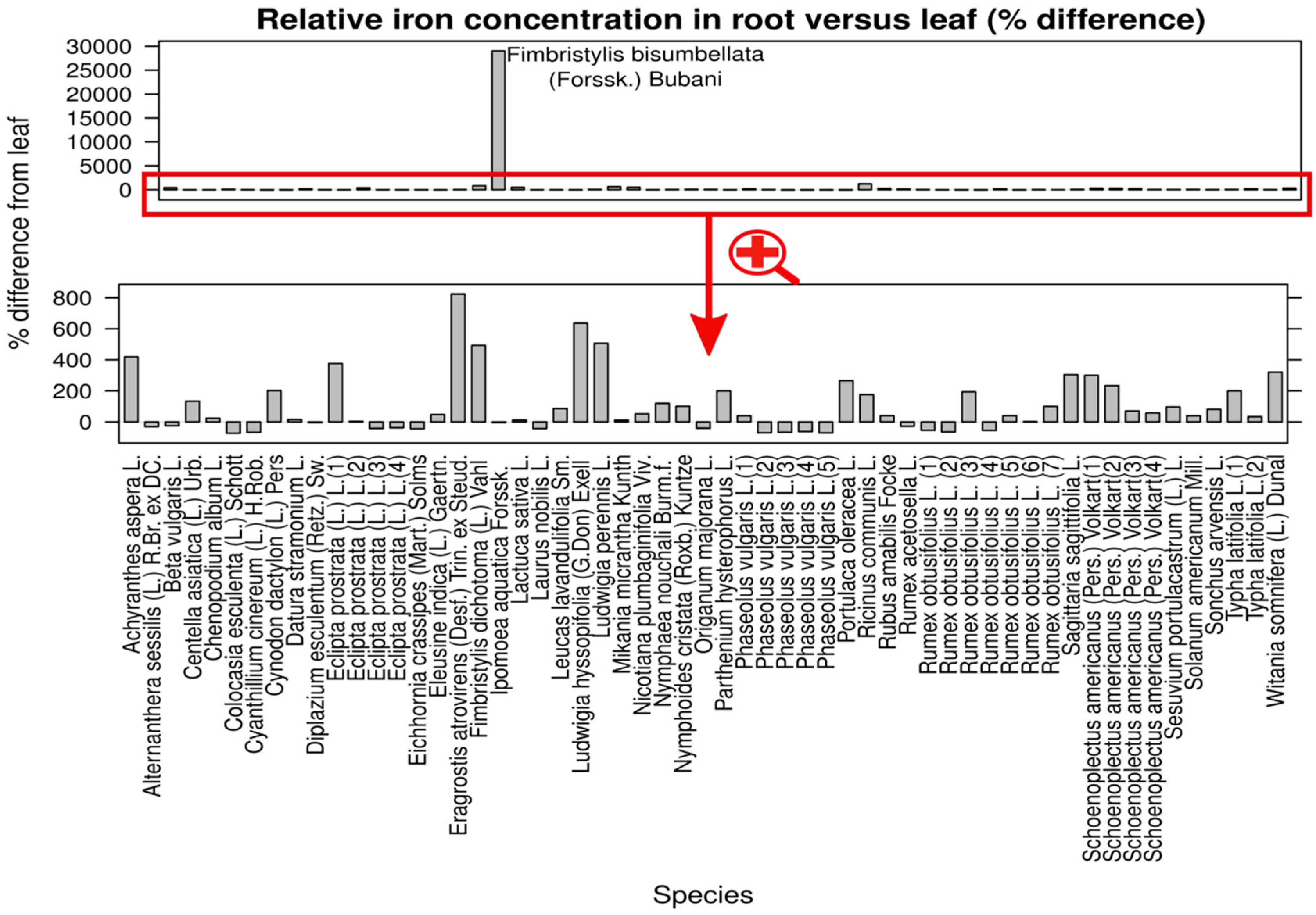
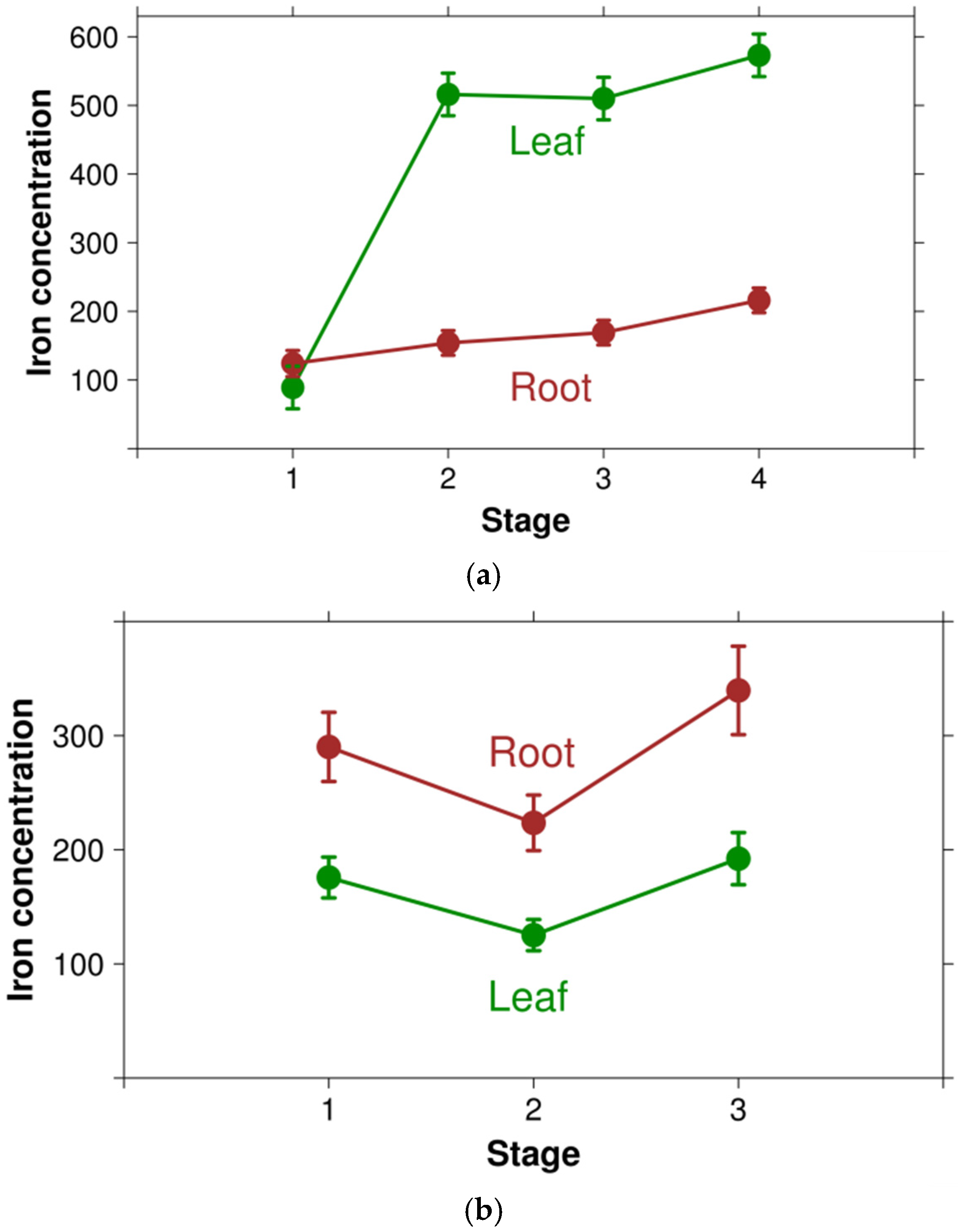
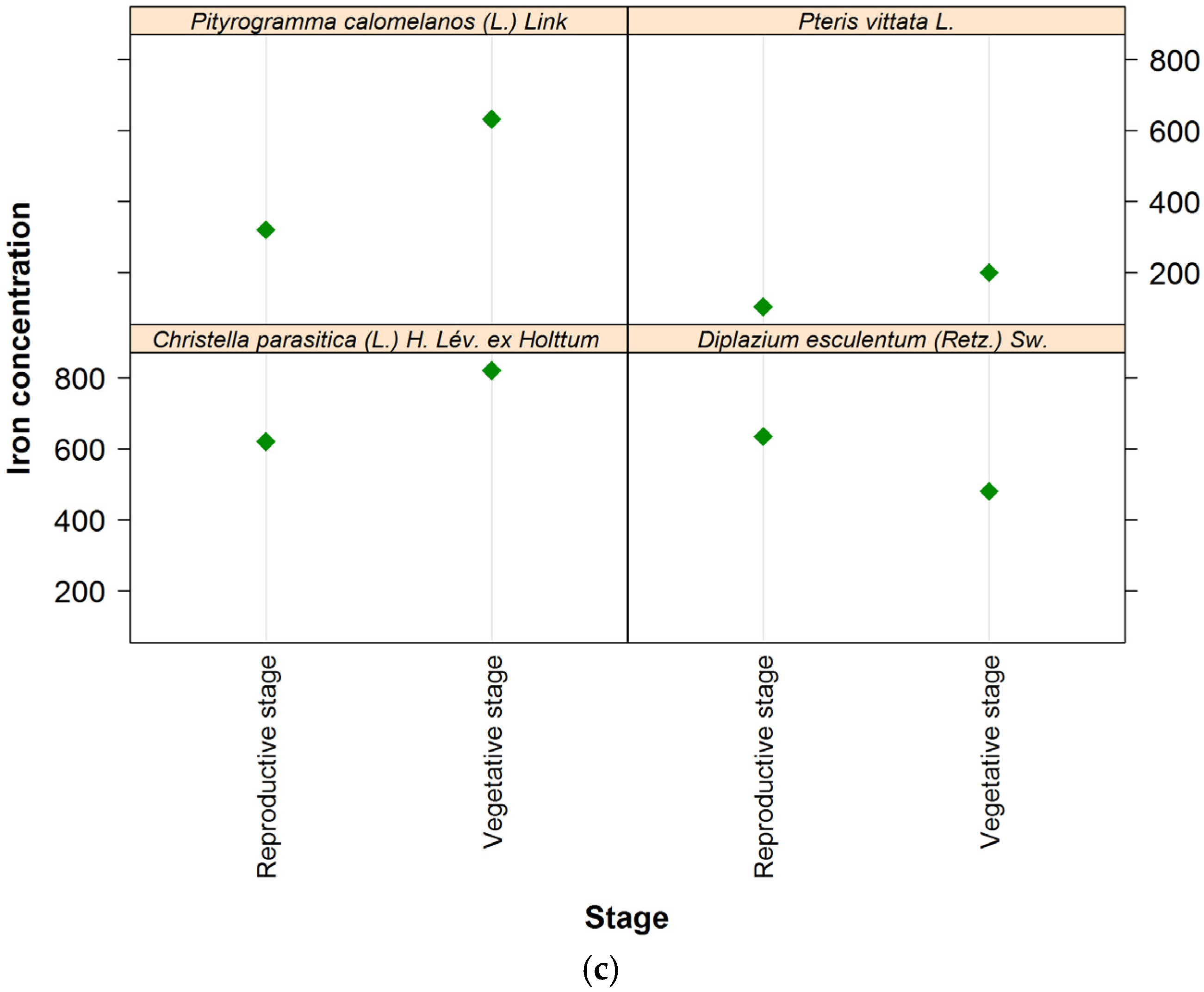
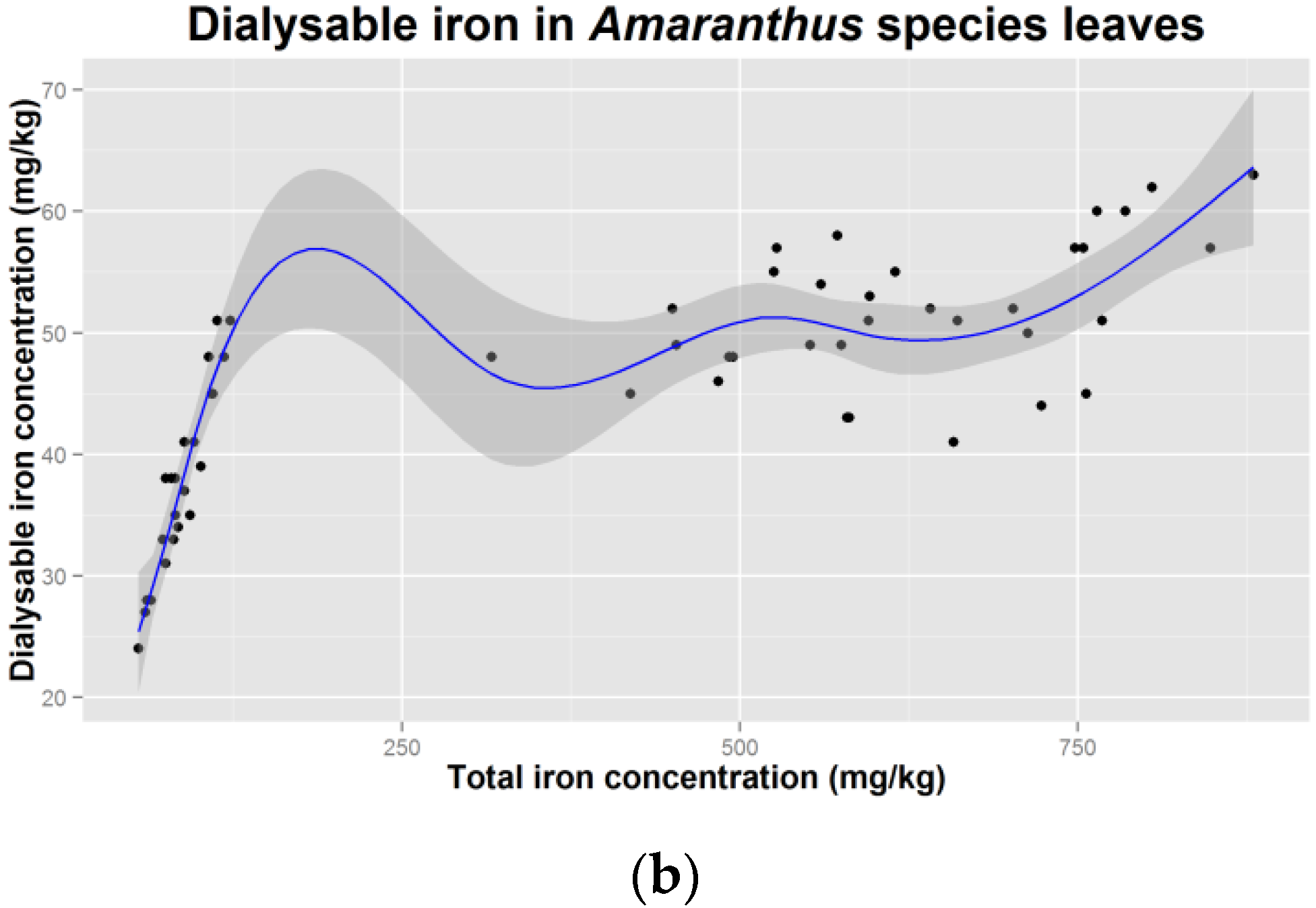
4. Discussion
4.1. Extent of Iron Contents Investigation in Plants
4.2. Iron Contents in Different Organs
4.3. Variability of Iron Contents in the Plant World
4.4. Limitations
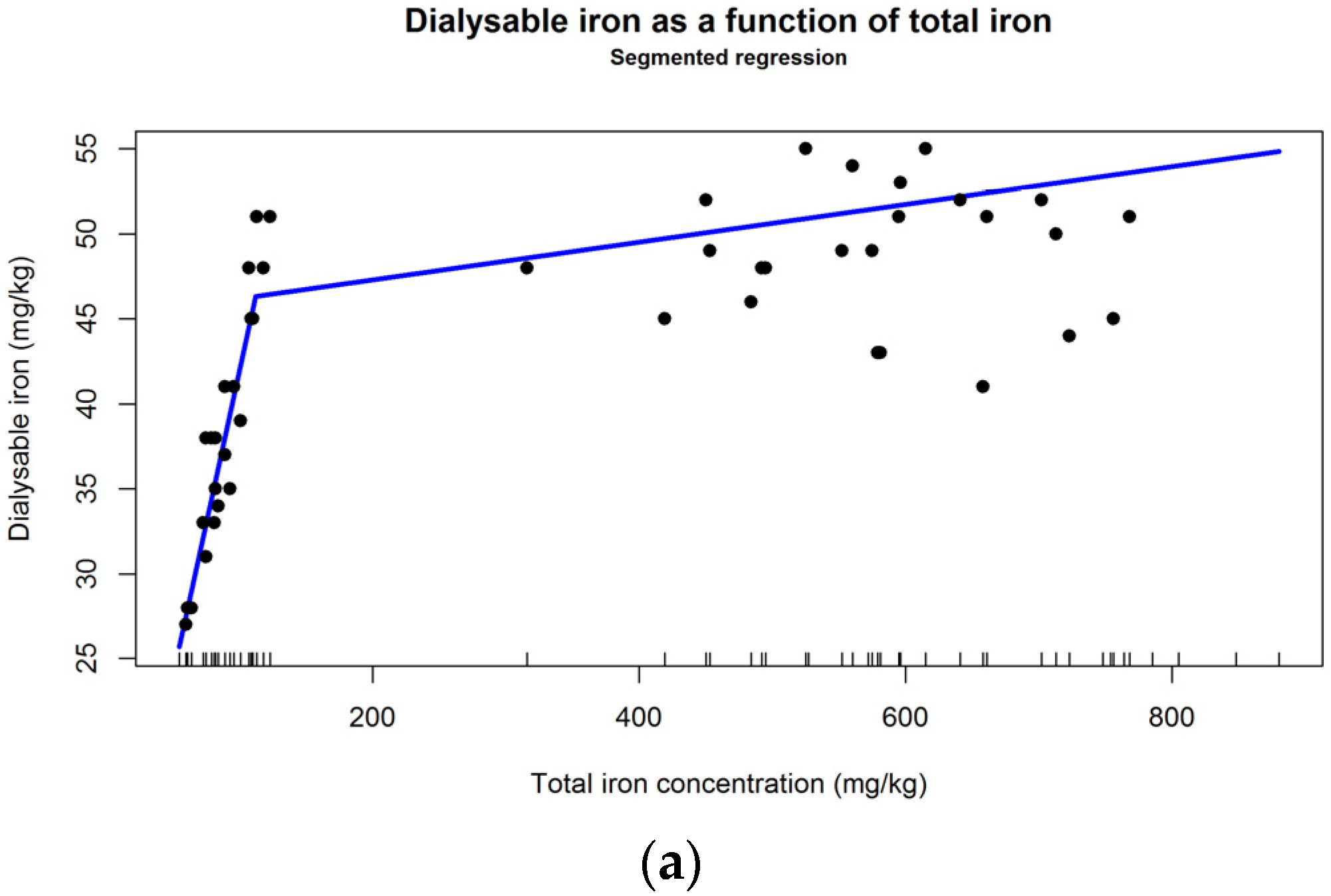
4.5. Absorption of Iron of Plant Origin in Humans
| Factor | Nature of Evidence | Comments |
|---|---|---|
| Phytic acid, phytates (from the plant source itself or from other foods, e.g., cereal bran) | In vitro, nonclinical, clinical (interventional) | It is the most studied factor influencing non-heme iron uptake, with very robust supporting evidence (e.g., [92,93,94,95,96,97,98,99,100]. |
| Polyphenols (from the plant source itself or from other food). | In vitro, nonclinical, clinical (interventional) | There is convincing evidence that polyphenols may interfere with human iron uptake from food, but not all polyphenols “are created equal” and as yet there is no complete picture of their effects, especially in the presence of other phytochemicals. In the case of cowpea (Vigna unguiculata (L.) Walp.) flour fortified with iron, there was no difference in iron absorption between a variety with low polyphenol content and one with high polyphenol content. The authors speculated that the reason might be that both had a similar phytate:iron ratio, which might be much more relevant for iron uptake than the concentration of polyphenols [96]. |
| Tannins (from the plant source itself or from other food). | In vitro, nonclinical, clinical (interventional) | Animal data (rat) and in vitro studies support an inhibitory role of tannins on iron uptake. 500 mg tannic acid added to a broccoli meal significantly decreased iron uptake (geometric mean 0.015 versus 0.297) [101]. It is also speculated that the inhibitory effect of green tea is related to its tannin and polyphenol contents [102]. But the matrix remains essential: although brown rice has significantly higher levels of tannic acid and phytate than milled rice, in a study in healthy adults no significant difference was identified in the amount of iron absorbed from the two types of rice [103]. |
| Green tea | Nonclinical (rat), Clinical (interventional) | A clinical trial [104] and several rat studies suggest that tea (through its polyphenols and aluminum) dose-dependently decrease iron uptake. An observational study in humans found no effect of black, green or herbal tea on serum ferritin or the iron depletion risk [105] (limitations of observational studies have to be taken into account, however). |
| Chilli | Clinical (observational, interventional) | An interventional study carried out in women [106] and an observational study in subjects of both genders found an inhibitory effect in female, but not in male chili consumers [107]. Unlike for humans, data in rats suggested that capsaicin (similarly to piperine or ginger) increases iron absorption [108]. |
| Iron mineral competitors | Nonclinical, clinical | Fe, Zn and Ca may interact with each other, reciprocally decreasing their bioavailability [109]. The effect of Zn is perceptible only when present in high levels (from 90 mg/kg upwards) [110]. Initially contradictory results have been published con calcium, some suggesting that calcium supplementation would negatively influence iron absorption [111,112], while more recent several long-term intervention studies have shown that long-term use of calcium supplements by post-menopausal women does not affect iron status. It has been suggested that iron absorption might be perturbed only on short-term, while on long term adaptive responses of the body might reestablish the iron balance [113]. |
| Certain fruits (orange, guava, kiwi) | Clinical (interventional) | The increase in uptake might be related to the ascorbic acid and possibly beta-carotene contents [114]. |
| Ascorbic acid | In vitro, nonclinical, clinical (interventional) | There is convincing evidence that ascorbic acid facilitates iron uptake, it may partially offset the negative effects of phytates [115] and of small amounts of polyphenols [116], but not the inhibitory effects of tannins [115] or of high amounts of polyphenols (as indicated by in vitro data) [116]. It seems more effective than EDTA [117] but has little effect in the presence of the former [93]. |
| Other organic acids (citric, erythorbic, malic, tartaric, succinic, fumaric, aminoacids, especially cysteine) | In vitro, nonclinical, clinical | The evidence is scarcer and less robust than for ascorbic acid and these organic acids seem substantially less effective than vitamin C [118]. Contradictory evidence exists for lactic and oxalic acids (no effect in some studies [119,120,121], positive effect in others [122,123,124], a slightly inhibitory effect in a study for oxalic acid [101]). |
| EDTA | Clinical (interventional) | EDTA facilitates iron absorption. Its effects seem inferior to those of the ascorbic acid [117], but the body of research is less extensive for EDTA. |
| Iron amount | Nonclinical, clinical | Although the relationship is nonlinear, there is relatively robust evidence that higher iron intake leads to higher (but not proportionally so) absorption [125]. |
| Meat protein (beef, fish and chicken) | In vitro, nonclinical, clinical | There is a relatively large body of evidence that animal protein (the so-called “meat factor”) favors iron uptake [126]. In a study, egg protein had a moderate inhibitory effect [127]. Several decades ago it was estimated that 30 grams of meat, poultry or fish are roughly equivalent to 25 mg ascorbic acid [128].There are quantitative differences among different meat sources (animal species) [129,130]. There are also differences in the effect of proteins on absorption in humans and rodents [131]. Plant-derived proteins have either no effect [132] or variable effects (most often inhibitory [133], rarely facilitatory [134]) depending on source and properties. In vitro data indicate that high molecular proteins have a better influence on iron absorption than low molecular weight proteins, irrespective of the source (animal, plant) [135], but the relevance of in vitro data for the clinical context is limited. |
| Plant species or variety | Nonclinical, in vitro | Different plant species (of the same genus even [136]) and different varieties of the same species [137] have different effects on iron uptake, probably depending on their phytochemical matrices. |
| Prebiotics | In vitro, nonclinical, clinical | Different prebiotics seem to have different effects on iron metabolism and uptake [138,139]. There are also discrepancies between the in vitro and in vivo data (the latter are more relevant to the clinical context) [140,141]. |
| Probiotics | In vitro, clinical (interventional) | Although theoretical mechanisms have been proposed by which various bacterial species might increase iron absorption, limited in vitro data show that this is not always the case [100,142], the bacterial species being also relevant. Certain lactic fermentation bacteria have been shown to facilitate iron absorption [119,140,143] and it seems that the effect is not due to lactic acid [119]. |
| Fructose | Nonclinical, clinical | In one clinical trial fructose (but not high fructose corn syrup) was reported to favor iron excretion and diminish the iron balance, probably by the induced diarrhea and consecutive lower absorption [144]. Nonclinical data (in vitro and in rats) claimed both increased [145] and limited [146] iron absorption. |
| Fructo-oligosaccharides | Nonclinical, clinical | Negative [147], neutral [148] and positive effects [149,150] have been recorded in various nonclinical experiments with fructooligosaccharides. In a clinical trial, no significant difference was observed versus the control group [151]. |
| l-alpha-glycerophosphocholine | Clinical | Identified in one study as the so-called “meat factor” and claimed to have improved iron absorption [126], in a different study it did not seem to influence iron uptake [93]. |
| Iron source (salt, complex) | In vitro, nonclinical, clinical | A number of studies have shown that iron is absorbed differently from various salts (e.g., NaFeEDTA is better absorbed from fortified soy sauce than FeSO4 [152]). Ion chelates with aminoacids (e.g., glycine) seem to be subject to less influence from inhibitors and enhancers than ferrous sulfate [153]. |
| Food cooking/processing (boiling, roasting, decortication, germination, fermentation, etc.). | In vitro | Various processing methods have different impacts on iron uptake (increase, decrease or no influence on availability), depending on the nature of treatment and of the food [154,155,156,157,158,159,160,161]. |
| Vitamin A, beta-carotene | Clinical, nonclinical | In a clinical experiment, vitamin A increased iron absorption from rice up to twofold, 0.8-fold from wheat (i.e., it caused a slight decrease in absorption) and 1.4-fold from corn. Beta-carotene increased iron absorption more than 3 times for rice and 1.8-fold for wheat and corn [162]. In a rat study, carotene was claimed to hinder iron absorption [163]. Clinical data should be considered more relevant and thus it is likely that carotene rather increases absorption. |
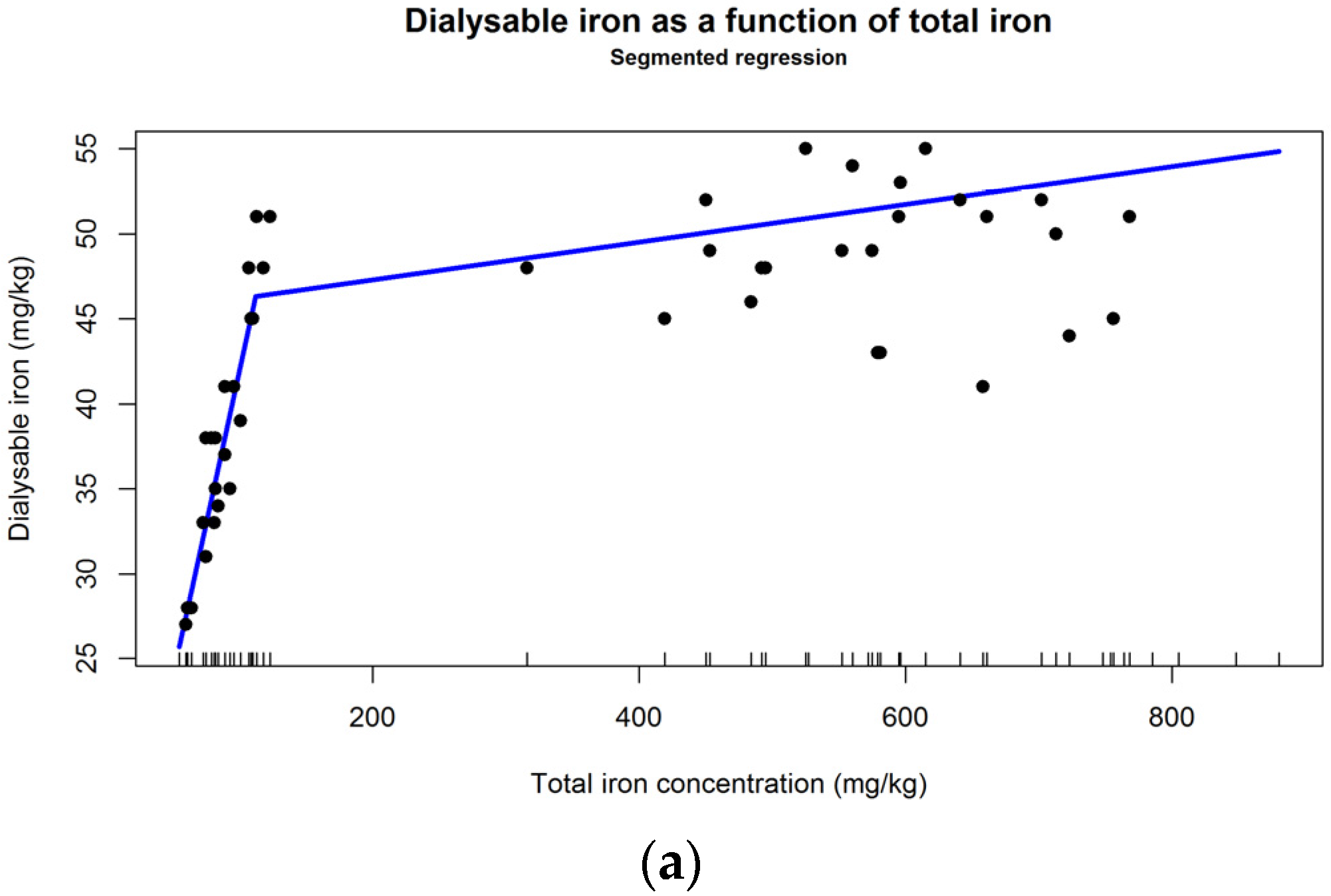
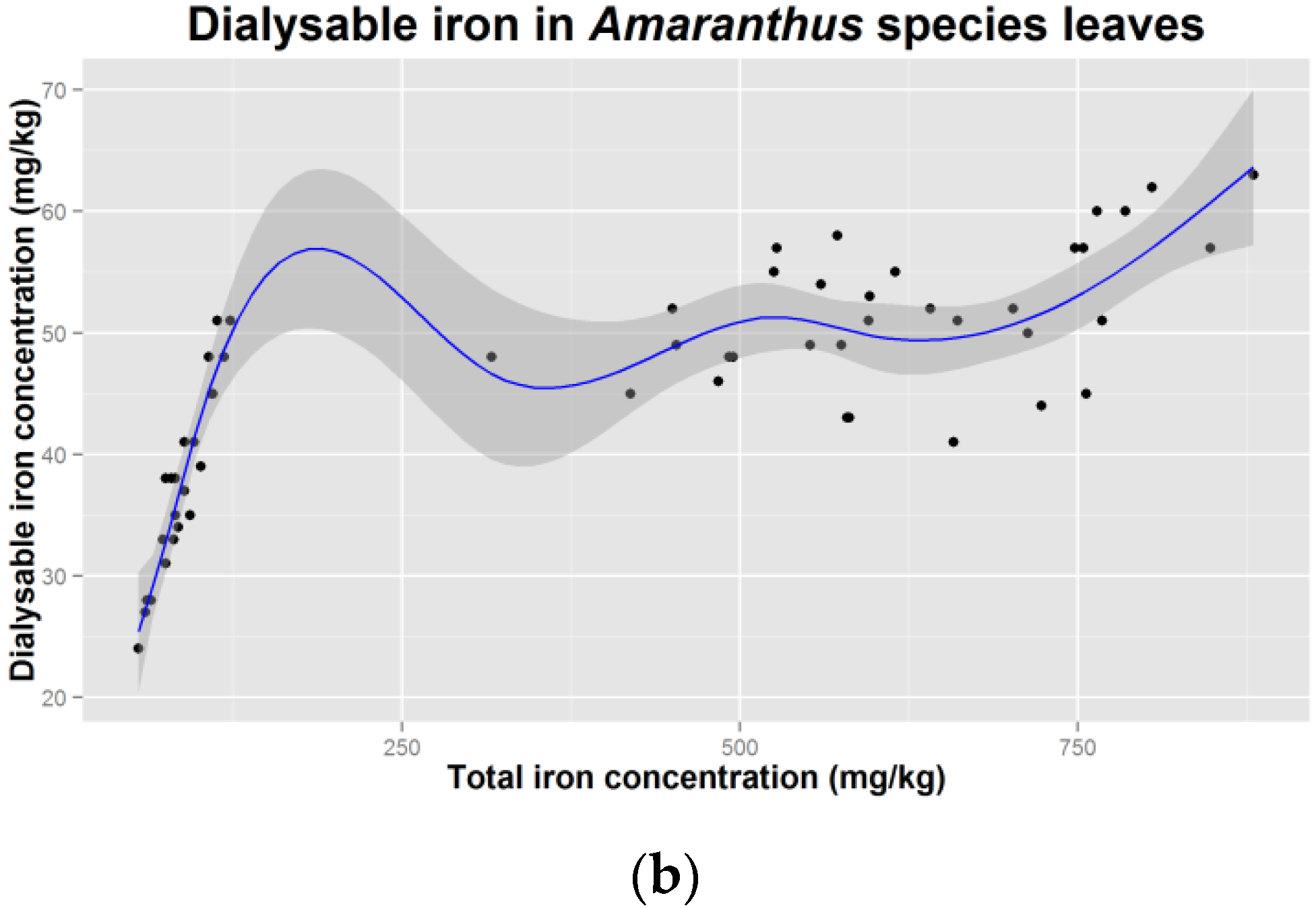
4.6. Could Iron of Herbal Origin Be a Meaningful Supplementation Option?
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Anderson, C.P.; Shen, M.; Eisenstein, R.S.; Leibold, E.A. Mammalian iron metabolism and its control by iron regulatory proteins. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2012, 1823, 1468–1483. [Google Scholar] [CrossRef] [PubMed]
- Rolfes, S.R.; Pinna, K.; Whitney, E.N. Understanding Normal and Clinical Nutrition, 8th ed.; Wadsworth/Cengage Learning: Belmont, CA, USA, 2009; p. 442. [Google Scholar]
- Webb, G.P. Dietary Supplements and Functional Foods, Kindle ed.; Blackwell Publishing: Chichester, UK, 2011. [Google Scholar]
- Sizer, F.; Whitney, E. Nutrition: Concepts and Controversies, 11th ed.; Thomson Higher Education: Belmont, CA, USA, 2008; p. 297. [Google Scholar]
- Khambalia, A.Z.; Zlotkin, S.H. Iron. In Nutrition in Pediatrics. Basic Science. Clinical Applications; Duggan, C., Watkins, J.B., Walker, W.A., Eds.; BC Dekker: Hamilton, ON, Canada, 2007; pp. 86–87. [Google Scholar]
- World Health Organisation and United Nations Children’s Fund (UNICEF). Focusing on Anaemia: Towards an Integrated Approach for Effective Anaemia Control. Available online: http://www.who.int/nutrition/publications/micronutrients/WHOandUNICEF_statement_anaemia_en.pdf?ua=1 (accessed on 6 December 2015).
- Parker-Williams, E.J. Investigation and management of anaemia. Medicine (Baltimore) 2009, 37, 137–142. [Google Scholar] [CrossRef]
- Blanc, B.; Finch, C.; Hallberg, L.; Herbert, V.; Lawcowicz, W.; Layrisse, M.; Mollin, D.; Rachmilewitz, M.; Ramalingaswami, V.; Sánchez-Medal, L.; et al. Nutritional anaemias. Report of a WHO Scientific Group. WHO Tech. Rep. Ser. 1968, 405, 1–40. [Google Scholar]
- Beutler, E.; Waalen, J. The definition of anemia: What is the lower limit of normal of the blood hemoglobin concentration? Blood 2006, 107, 1747–1750. [Google Scholar] [CrossRef] [PubMed]
- Boehringer, P.A.; Darden, I.L. The quest for the normal hemoglobin concentration. Blood 2006, 108, 777–778. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.; Greene-Finestone, L.; Lowell, H.; Levesque, J.; Robinson, S. Iron sufficiency of Canadians. Health Rep. 2012, 23, 41–48. [Google Scholar] [PubMed]
- Ho, C.-H.; Yu, Y.-B.; Wu, P.-H. The prevalence of iron deficiency anemia and its clinical implications in patients with colorectal carcinoma. J. Chin. Med. Assoc. JCMA 2008, 71, 119–122. [Google Scholar] [CrossRef]
- Barroso, F.; Allard, S.; Kahan, B.C.; Connolly, C.; Smethurst, H.; Choo, L.; Khan, K.; Stanworth, S. Prevalence of maternal anaemia and its predictors: A multi-centre study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Stoltzfus, R.J.; Dreyfuss, M.L. Guidelines for the Use of Iron Supplements to Prevent and Treat Iron Deficiency Anemia; Ilsi Press: Washington, DC, USA, 1998. [Google Scholar]
- The Plant List. A Working List of All Plant Species. Available online: Http://www.theplantlist.org (accessed on 8 August 2015).
- Eddelbuettel, D.D. Random: True Random Numbers Using Random.Org. 2013. R Package Version 0.2.2. Available online: http://CRAN.R-project.org/package=random (accessed on 25 August 2015).
- Plants for A Future: A Resource and Information Centre for Edible and Otherwise Useful Plants. Available online: http://www.pfaf.org (accessed on 25 August 2015).
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar]
- Champely, S. Pwr: Basic Functions for Power Analysis. R Package Version 1.1-2. 2015. Available online: http://CRAN.R-project.org/package=pwr (accessed on 25 August 2015).
- Tokalıoğlu, Ş. Determination of trace elements in commonly consumed medicinal herbs by ICP-MS and multivariate analysis. Food Chem. 2012, 134, 2504–2508. [Google Scholar]
- Ştef, D.S.; Gergen, I.; Traşcă, T.I.; Hărmănescu, M.; Ştef, L.; Drugă, M.; Biron, R.; Hegheduş-Mîndru, G. Screening of 33 Medicinal Plants for the Microelements Content. Anim. Sci. Biotechnol. 2010, 43, 127–132. [Google Scholar]
- Kolasani, A.; Xu, H.; Millikan, M. Evaluation of mineral content of Chinese medicinal herbs used to improve kidney function with chemometrics. Food Chem. 2011, 127, 1465–1471. [Google Scholar] [CrossRef]
- Nomita Devi, K.; Nandakumar Sarma, H. PIXE–PIGE analysis of some Indian medicinal plants. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2010, 268, 2144–2147. [Google Scholar] [CrossRef]
- Başgel, S.; Erdemoğlu, S.B. Determination of mineral and trace elements in some medicinal herbs and their infusions consumed in Turkey. Sci. Total Environ. 2006, 359, 82–89. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing, 2015. Available online: Http://www.R-project.org/ (accessed on 25 August 2015).
- Rmetrics Core Team; Wuertz, D.; Setz, T.; Chalabi, Y. fBasics: Rmetrics—Markets and Basic Statistics, 2014. R Package Version 3011.87. 2014. Available online: Http://CRAN.R-project.org/package=fBasics (accessed on 25 August 2015).
- Gastwirth, J.L.; Gel, Y.R.; Hui, W.L.W.; Miao, W.; Noguchi, K. lawstat: Tools for Biostatistics, Public Policy, and Law. R Package Version 2.5. 2015. Available online: Http://CRAN.R-project.org/package=lawstat (accessed on 25 August 2015).
- Peng, R.D. Simpleboot: Simple Bootstrap Routines. R Package Version 1.1-3. 2008. Available online: Https://cran.r-project.org/web/packages/simpleboot/ (accessed on 25 August 2015).
- Van der Loo, M.P.J. Distribution Based Outlier Detection for Univariate Data; Discussion paper 10003; Statistics Netherlands: The Hague, The Netherlands, 2010. [Google Scholar]
- Konietschke, F.; Placzek, M.; Schaarschmidt, F.; Hothorn, L.A. nparcomp: An R Software Package for Nonparametric Multiple Comparisons and Simultaneous Confidence Intervals. J. Stat. Softw. 2015, 64, 1–17. [Google Scholar] [CrossRef]
- Cribbie, R.A.; Wilcox, R.R.; Bewell, C.; Keselman, H.J. Tests for Treatment Group Equality When Data are Nonnormal and Heteroscedastic. J. Mod. Appl. Stat. Methods 2007, 6. Available online: Http://digitalcommons.wayne.edu/jmasm/vol6/iss1/12 (accessed on 25 August 2015). [Google Scholar]
- Konietschke, F.; Hothorn, L.A.; Brunner, E. Rank-based multiple test procedures and simultaneous confidence intervals. Electron. J. Stat. 2012, 6, 738–759. [Google Scholar] [CrossRef]
- Torchiano, M. Effsize: Efficient Effect Size Computation. R Package Version 0.5.4. 2015. Available online: Http://CRAN.R-project.org/package=effsize (accessed on 25 May 2015).
- Cohen, B.H. Explaining Psychological Statistics; John Wiley & Sons: Hoboken, NJ, USA, 2008; p. 750. [Google Scholar]
- Kinnear, P.R.; Gray, C.D.; Kinnear, P.R. PASW Statistics 17 Made Simple; Psychology Press: Hove, East Sussex; New York, NY, USA, 2010; p. 255. [Google Scholar]
- Tomczak, M.; Tomcak, E. The need to report effect size estimates revisited. An overview of some recommended measures of effect size. Trends Sport Sci. 2014, 1, 19–25. [Google Scholar]
- Feinstein, A.R. Principles of Medical Statistics; Chapman & Hall/CRC: Boca Raton, FL, USA, 2002; pp. 250–251. [Google Scholar]
- Encyclopedia of Research Design; volume 3; Salkind, N.J. (Ed.) SAGE Publications: Thousand Oaks, CA, USA, 2010; pp. 1647–1648.
- Muggeo, V.M.R. Segmented: An R Package to Fit Regression Models with Broken-Line Relationships. R News 2008, 8, 20–25. [Google Scholar]
- Muggeo, V.M.R. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Use R! Springer: New York, NY, USA, 2009. [Google Scholar]
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Use R! Springer: New York, NY, USA, 2008. [Google Scholar]
- Marbaniang, D.G.; Baruah, P.; Decruse, R.; Dkhar, E.R.; Diengdoh, D.F.; Nongpiur, C.L. Study of the Trace Metal Concentration in Some Local Vegetables Available in Shillong City, Meghlaya, India. Int. J. Environ. Prot. 2012, 2, 24–28. [Google Scholar]
- Karmakar, K.; Muslim, T.; Rahman, M. Chemical Composition of Some Leafy Vegetables of Bangladesh. Dhaka Univ. J. Sci. 2013, 61, 199–201. [Google Scholar]
- Ayala-Vela, J.; Guía-González, M.; Espinosa-Huerta, E.; Acosta-Gallegos, J.A.; Guzmán-Maldonado, S.H.; Mora-Avilés, M.A. Iron Content and Ferritin Gene Expression in Common Bean (Phaseolus vulgaris L.). Agric. Tech. Mex. 2008, 34, 481–489. [Google Scholar]
- Pillay, V.; Jonnalagadda, S.B. Elemental uptake by edible herbs and lettuce (Latuca sativa). J. Environ. Sci. Health Part B 2007, 42, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Masal, V.P.; Meena, M. Screening of Pteridophytes for Inorganic Constituents in Some Species from Ratnagiri District of Maharashtra (India). Bionano Front. 2010, 3, 273–275. [Google Scholar]
- Liu, Y.-J.; Ding, H.; Zhu, Y. Metal bioaccumulation in plant leaves from an industrious area and the botanical garden in Beijing. J. Environ. Sci. China 2005, 17, 294–300. [Google Scholar] [PubMed]
- Essiett, U.A.; Effiong, G.S.; Ogbemudia, F.O.; Bruno, E.J. Heavy metal concentrations in plants growing in crude oil contaminated soil in Akwa Ibom State, South-Eastern Nigeria. Afr. J. Pharm. Pharmacol. 2010, 4, 465–470. [Google Scholar]
- Hussain, I.; Khan, H. Investigation of Heavy Metal Content in Medicinal Plant, Eclipta alba L. J. Chem. Soc. Pak. 2010, 32. [Google Scholar]
- Rangarajan, A.; Kelly, J.F. Iron bioavailability fromAmaranthus species: 1—In vitro dialysable iron for estimation of genetic variation. J. Sci. Food Agric. 1998, 78, 267–273. [Google Scholar] [CrossRef]
- Musa, A.; Ogbadoyi, E.O. Effect of Plant Leaf Positions on Some Micronutrients, Anti-nutrients and Toxic Substances in Telfairia occidentalis at the Vegetative Phase. Am. J. Exp. Agric. 2012, 2, 219–232. [Google Scholar] [CrossRef]
- Aciksoz, S.B.; Ozturk, L.; Gokmen, O.O.; Römheld, V.; Cakmak, I. Effect of nitrogen on root release of phytosiderophores and root uptake of Fe(III)- phytosiderophore in Fe-deficient wheat plants. Physiol. Plant. 2011, 142, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Yue, S.; Zhang, W.; Liu, D.; Cui, Z.; Chen, X.; Ye, Y.; Zou, C. Zinc, iron, manganese and copper uptake requirement in response to nitrogen supply and the increased grain yield of summer maize. PLoS ONE 2014, 9, e93895. [Google Scholar] [CrossRef] [PubMed]
- Rode, K.D.; Chapman, C.A.; Chapman, L.J.; McDowell, L.R. Mineral Resource Availability and Consumption by Colobus in Kibale National Park, Uganda. Int. J. Primatol. 2003, 24. [Google Scholar] [CrossRef]
- Vardanyan, L.G.; Ingole, B.S. Studies on heavy metal accumulation in aquatic macrophytes from Sevan (Armenia) and Carambolim (India) lake systems. Environ. Int. 2006, 32, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, K.; Das, S. Phytoremediation of Pb, Zn, Fe, and Mg with 25 wetland plant species from a paper mill contaminated site in North East India. Environ. Sci. Pollut. Res. Int. 2015, 22, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Ernst, W.H.O. Element Nutrition of Two Contrasted Dune Annuals. J. Ecol. 1983, 71, 197–209. [Google Scholar] [CrossRef]
- Mayer, A.M.; Gorham, E. The Iron and Manganese Content of Plants present in the Natural Vegetation of the English Lake District. Ann. Bot. 1951, 15, 247–263. [Google Scholar]
- Uyar, G.; Ören, M.; Yildirim, Y.; Öncel, S. Biomonitoring of Metal Deposition in the Vicinity of Eregli Steel Plant in Turkey. Environ. Forensics 2008, 9, 350–363. [Google Scholar] [CrossRef]
- Mohamed, A.I.; Hussein, A.S. Chemical composition of purslane (Portulaca oleracea). Plant Foods Hum. Nutr. Dordr. Neth. 1994, 45, 1–9. [Google Scholar] [CrossRef]
- Liu, J.; Reid, R.J.; Smith, F.A. The mechanism of cobalt toxicity in mung beans. Physiol. Plant. 2000, 110, 104–110. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation. Plant J. 2008, 54, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.; Guerinot, M.L. Iron Uptake and Transport in Plants: The Good, the Bad, and the Ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef] [PubMed]
- Grusak, M. Whole -root iron(III)-reductase activity throughout the life cycle of iron-grown Pisum sativum L. (Fabaceae): Relevance to the iron nutrition of developing seeds. Planta 1995, 197. [Google Scholar] [CrossRef]
- Kim, S.A.; Guerinot, M.L. Mining iron: Iron uptake and transport in plants. FEBS Lett. 2007, 581, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Lung’aho, M.G.; Mwaniki, A.M.; Szalma, S.J.; Hart, J.J.; Rutzke, M.A.; Kochian, L.V.; Glahn, R.P.; Hoekenga, O.A. Genetic and physiological analysis of iron biofortification in maize kernels. PLoS ONE 2011, 6, e20429. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Srivastava, J.; Singh, N.; Singh, S.P. Mineral profile and variability in vegetable amaranth (Amaranthus tricolor). Plant Foods Hum. Nutr. Dordr. Neth. 2006, 61, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Thavarajah, D.; Thavarajah, P.; Sarker, A.; Vandenberg, A. Lentils (Lens culinaris Medikus Subspecies culinaris): A whole food for increased iron and zinc intake. J. Agric. Food Chem. 2009, 57, 5413–5419. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.; Hernandez, A.J.; Estalrich, E.; Oliver, S. Soil Factors and Fe Content In Wild Herbaceous Plants. In Iron Nutrition in Soil and Plants; Abadia, J., Ed.; Springer Science + Business Media: Dordrecht, The Netherlands, 1995; pp. 159–166. [Google Scholar]
- Gupta, K.K.; Bhattacharjee, S.; Kar, S.; Chakrabarty, S.; Thakur, P.; Bhattacharyya, G.; Srivastava, S.C. Mineral Compositions of Eight Common Spices. Commun. Soil Sci. Plant Anal. 2003, 34, 681–693. [Google Scholar] [CrossRef]
- Ozkutlu, F. Determination of cadmium and trace elements in some spices cultivated in Turkey. Asian J. Chem. 2008, 20, 1081–1088. [Google Scholar]
- Alamin, M.B.; Mhapes, A.A.; Bejey, A.M.; Sadek, A.; Atweer, R.H.; Dubali, K.; Saad, D.M. Determination of essential and toxic elements in Libyan foodstuff using instrumental neutron activation analysis (INAA). J. Radioanal. Nucl. Chem. 2007, 271, 247–250. [Google Scholar] [CrossRef]
- Brudzińska-Kosior, A.; Kosior, G.; Samecka-Cymerman, A.; Kolon, K.; Mróz, L.; Kempers, A.J. Metal contents in Centaurium erythraea and its biometry at various levels of environmental pollution. Ecotoxicol. Environ. Saf. 2012, 80, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Dodson, H.G. Total Iron in Selected Genotypes of Spinach and Cowpeas and the in Vitro Iron Bioavailability of Selected Spinach Genotypes, University of Arkansas. 2008. Available online: http://tinyurl.com/qxqu4gb (accessed on 1 September 2015).
- Van den Heuvel, E.G.H.M.; van Dokkum, W.; Schaafsma, G. Methods to measure iron absorption in humans: A review. Food Rev. Int. 1997, 13, 91–102. [Google Scholar] [CrossRef]
- Insel, P.M.; Ross, D.; McMahon, K.; Bernstein, M. Discovering Nutrition, 4th ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2013; p. 411. [Google Scholar]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef] [PubMed]
- López, M.A.A.; Martos, F.C. Iron availability: An updated review. Int. J. Food Sci. Nutr. 2004, 55, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, A.; Cleton, F.; Finch, C.A. Iron Absorption. IV. the Absorption of Hemoglobin Iron *. J. Clin. Invest. 1962, 41, 1897–1907. [Google Scholar] [CrossRef] [PubMed]
- Layrisse, M.; Martínez-Torres, C. Model for measuring dietary absorption of heme iron: Test with a complete meal. Am. J. Clin. Nutr. 1972, 25, 401–411. [Google Scholar] [PubMed]
- Bezwoda, W.R.; Bothwell, T.H.; Charlton, R.W.; Torrance, J.D.; MacPhail, A.P.; Derman, D.P.; Mayet, F. The relative dietary importance of haem and non-haem iron. S. Afr. Med. J. 1983, 64, 552–556. [Google Scholar] [PubMed]
- Callender, S.T.; Mallett, B.J.; Smith, M.D. Absorption of Haemoglobin Iron. Br. J. Haematol. 1957, 3, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, L. Iron requirements and bioavailability of dietary iron. Experientia. Suppl. 1983, 44, 223–244. [Google Scholar] [PubMed]
- Nakajima, V.M.; Costa, N.M.; Martino, H.S.; Queiroz, V.A.; Guimarães, P.E.; Oliveira, P.V. Iron bioavailability of different maize genotypes developed in a breeding program: In vitro and in vivo studies. Arch. Latinoam. Nutr. 2012, 62, 103–111. [Google Scholar] [PubMed]
- Zielińska-Dawidziak, M.; Hertig, I.; Piasecka-Kwiatkowska, D.; Staniek, H.; Nowak, K.W.; Twardowski, T. Study on iron availability from prepared soybean sprouts using an iron-deficient rat model. Food Chem. 2012, 135, 2622–2627. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, F.; Latunde-Dada, G.O. Iron bioavailibity from a tropical leafy vegetable in anaemic mice. Nutr. Metab. 2011, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Lung’aho, M.G.; Glahn, R.P. In vitro estimates of iron bioavailability in some Kenyan complementary foods. Food Nutr. Bull. 2009, 30, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xie, W.; Hao, Z.; Jin, X.; Wang, Q. Use of shallot ( Allium ascalonicum) and leek (Allium tuberosum) to improve the in vitro available iron and zinc from cereals and legumes. CyTA J. Food 2014, 12, 195–198. [Google Scholar] [CrossRef]
- Thankachan, P.; Kalasuramath, S.; Hill, A.L.; Thomas, T.; Bhat, K.; Kurpad, A.V. A mathematical model for the hemoglobin response to iron intake, based on iron absorption measurements from habitually consumed Indian meals. Eur. J. Clin. Nutr. 2012, 66, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Beiseigel, J.M.; Hunt, J.R.; Glahn, R.P.; Welch, R.M.; Menkir, A.; Maziya-Dixon, B.B. Iron bioavailability from maize and beans: A comparison of human measurements with Caco-2 cell and algorithm predictions. Am. J. Clin. Nutr. 2007, 86, 388–396. [Google Scholar] [PubMed]
- Koréissi-Dembélé, Y.; Fanou-Fogny, N.; Moretti, D.; Schuth, S.; Dossa, R.A.M.; Egli, I.; Zimmermann, M.B.; Brouwer, I.D. Dephytinisation with intrinsic wheat phytase and iron fortification significantly increase iron absorption from fonio (Digitaria exilis) meals in West African women. PLoS ONE 2013, 8, e70613. [Google Scholar] [CrossRef] [PubMed]
- Troesch, B.; Egli, I.; Zeder, C.; Hurrell, R.F.; de Pee, S.; Zimmermann, M.B. Optimization of a phytase-containing micronutrient powder with low amounts of highly bioavailable iron for in-home fortification of complementary foods. Am. J. Clin. Nutr. 2009, 89, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Campion, B.; Nielsen, E.; Hurrell, R. Genetic reduction of phytate in common bean (Phaseolus vulgaris L.) seeds increases iron absorption in young women. J. Nutr. 2013, 143, 1219–1224. [Google Scholar] [CrossRef] [PubMed]
- Cercamondi, C.I.; Egli, I.M.; Mitchikpe, E.; Tossou, F.; Hessou, J.; Zeder, C.; Hounhouigan, J.D.; Hurrell, R.F. Iron bioavailability from a lipid-based complementary food fortificant mixed with millet porridge can be optimized by adding phytase and ascorbic acid but not by using a mixture of ferrous sulfate and sodium iron EDTA. J. Nutr. 2013, 143, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Abizari, A.-R.; Moretti, D.; Schuth, S.; Zimmermann, M.B.; Armar-Klemesu, M.; Brouwer, I.D. Phytic acid-to-iron molar ratio rather than polyphenol concentration determines iron bioavailability in whole-cowpea meal among young women. J. Nutr. 2012, 142, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Troesch, B.; van Stuijvenberg, M.E.; van Stujivenberg, M.E.; Smuts, C.M.; Kruger, H.S.; Biebinger, R.; Hurrell, R.F.; Baumgartner, J.; Zimmermann, M.B. A micronutrient powder with low doses of highly absorbable iron and zinc reduces iron and zinc deficiency and improves weight-for-age Z-scores in South African children. J. Nutr. 2011, 141, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.; Taylor, J.R.N.; Du, X.; de Moura, F.F.; Lönnerdal, B.; Oelofse, A. Effect of phytate reduction of sorghum, through genetic modification, on iron and zinc availability as assessed by an in vitro dialysability bioaccessibility assay, Caco-2 cell uptake assay, and suckling rat pup absorption model. Food Chem. 2013, 141, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Penella, J.M.; Laparra, J.M.; Sanz, Y.; Haros, M. Assessment of iron bioavailability in whole wheat bread by addition of phytase-producing bifidobacteria. J. Agric. Food Chem. 2012, 60, 3190–3195. [Google Scholar] [CrossRef] [PubMed]
- Gillooly, M.; Bothwell, T.H.; Torrance, J.D.; MacPhail, A.P.; Derman, D.P.; Bezwoda, W.R.; Mills, W.; Charlton, R.W.; Mayet, F. The effects of organic acids, phytates and polyphenols on the absorption of iron from vegetables. Br. J. Nutr. 1983, 49, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Marouani, N.; Chahed, A.; Hédhili, A.; Hamdaoui, M.H. Both aluminum and polyphenols in green tea decoction (Camellia sinensis) affect iron status and hematological parameters in rats. Eur. J. Nutr. 2007, 46, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Trinidad, T.P.; Mallillin, A.C.; Sagum, R.S.; Briones, D.P.; Encabo, R.R.; Juliano, B.O. Iron absorption from brown rice/brown rice-based meal and milled rice/milled rice-based meal. Int. J. Food Sci. Nutr. 2009, 60, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Disler, P.B.; Lynch, S.R.; Charlton, R.W.; Torrance, J.D.; Bothwell, T.H.; Walker, R.B.; Mayet, F. The effect of tea on iron absorption. Gut 1975, 16, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Mennen, L.; Hirvonen, T.; Arnault, N.; Bertrais, S.; Galan, P.; Hercberg, S. Consumption of black, green and herbal tea and iron status in French adults. Eur. J. Clin. Nutr. 2007, 61, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Tuntipopipat, S.; Judprasong, K.; Zeder, C.; Wasantwisut, E.; Winichagoon, P.; Charoenkiatkul, S.; Hurrell, R.; Walczyk, T. Chili, but not turmeric, inhibits iron absorption in young women from an iron-fortified composite meal. J. Nutr. 2006, 136, 2970–2974. [Google Scholar] [PubMed]
- Li, J.; Wang, R.; Xiao, C. Association between chilli food habits with iron status and insulin resistance in a Chinese population. J. Med. Food 2014, 17, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Prakash, U.N.S.; Srinivasan, K. Enhanced intestinal uptake of iron, zinc and calcium in rats fed pungent spice principles—Piperine, capsaicin and ginger (Zingiber officinale). J. Trace Elem. Med. Biol. 2013, 27, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Anjum, F.M.; Ur Rehman, S.; Randhawa, M.A.; Farooq, U. Bioavailability of calcium, iron and zinc fortified whole wheat flour chapatti. Plant Foods Hum. Nutr. Dordr. Neth. 2008, 63, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Pizarro, F.; de Romaña, D.L. Effect of zinc sulfate fortificant on iron absorption from low extraction wheat flour co-fortified with ferrous sulfate. Biol. Trace Elem. Res. 2013, 151, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Seligson, F.H.; Hughes, V.A. Effects of calcium carbonate and hydroxyapatite on zinc and iron retention in postmenopausal women. Am. J. Clin. Nutr. 1986, 44, 83–88. [Google Scholar] [PubMed]
- Cook, J.D.; Dassenko, S.A.; Whittaker, P. Calcium supplementation: Effect on iron absorption. Am. J. Clin. Nutr. 1991, 53, 106–111. [Google Scholar] [PubMed]
- Bendich, A. Calcium supplementation and iron status of females. Nutr. Burbank Los Angel. Cty. Calif. 2001, 17, 46–51. [Google Scholar] [CrossRef]
- Purushothaman, V.; Amirthaveni, M.; Tsou, S.C.S.; Shanmugasundaram, S. Supplementing iron bioavailability enhanced mung bean. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 99–102. [Google Scholar] [PubMed]
- Jin, F.; Frohman, C.; Thannhauser, T.W.; Welch, R.M.; Glahn, R.P. Effects of ascorbic acid, phytic acid and tannic acid on iron bioavailability from reconstituted ferritin measured by an in vitro digestion-Caco-2 cell model. Br. J. Nutr. 2009, 101, 972–981. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Kim, E.-Y.; Lindsay, E.A.; Han, O. Bioactive dietary polyphenols inhibit heme iron absorption in a dose-dependent manner in human intestinal Caco-2 cells. J. Food Sci. 2011, 76, H143–H150. [Google Scholar] [CrossRef] [PubMed]
- Cercamondi, C.I.; Egli, I.M.; Zeder, C.; Hurrell, R.F. Sodium iron EDTA and ascorbic acid, but not polyphenol oxidase treatment, counteract the strong inhibitory effect of polyphenols from brown sorghum on the absorption of fortification iron in young women. Br. J. Nutr. 2014, 111, 481–489. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Teucher, B.; Olivares, M.; Cori, H. Enhancers of iron absorption: Ascorbic acid and other organic acids. Int. J. Vitam. Nutr. Res. 2004, 74, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Bering, S.; Suchdev, S.; Sjøltov, L.; Berggren, A.; Tetens, I.; Bukhave, K. A lactic acid-fermented oat gruel increases non-haem iron absorption from a phytate-rich meal in healthy women of childbearing age. Br. J. Nutr. 2006, 96, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Baynes, R.D.; Macfarlane, B.J.; Bothwell, T.H.; Siegenberg, D.; Bezwoda, W.R.; Schmidt, U.; Lamparelli, R.D.; Mayet, F.; MacPhail, A.P. The promotive effect of soy sauce on iron absorption in human subjects. Eur. J. Clin. Nutr. 1990, 44, 419–424. [Google Scholar] [PubMed]
- Genannt Bonsmann, S.S.; Walczyk, T.; Renggli, S.; Hurrell, R.F. Oxalic acid does not influence nonhaem iron absorption in humans: A comparison of kale and spinach meals. Eur. J. Clin. Nutr. 2008, 62, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Derman, D.P.; Bothwell, T.H.; Torrance, J.D.; Bezwoda, W.R.; MacPhail, A.P.; Kew, M.C.; Sayers, M.H.; Disler, P.B.; Charlton, R.W. Iron absorption from maize (Zea mays) and sorghum (Sorghum vulgare) beer. Br. J. Nutr. 1980, 43, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.T.; Chao, L.S. Relationship of components in wheat bran and spinach to iron bioavailability in the anemic rat. J. Nutr. 1984, 114, 526–535. [Google Scholar] [PubMed]
- Van Campen, D.R.; Welch, R.M. Availability to rats of iron from spinach: Effects of oxalic acid. J. Nutr. 1980, 110, 1618–1621. [Google Scholar] [PubMed]
- Cercamondi, C.I.; Egli, I.M.; Mitchikpe, E.; Tossou, F.; Zeder, C.; Hounhouigan, J.D.; Hurrell, R.F. Total iron absorption by young women from iron-biofortified pearl millet composite meals is double that from regular millet meals but less than that from post-harvest iron-fortified millet meals. J. Nutr. 2013, 143, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Armah, C.N.; Sharp, P.; Mellon, F.A.; Pariagh, S.; Lund, E.K.; Dainty, J.R.; Teucher, B.; Fairweather-Tait, S.J. l-alpha-glycerophosphocholine contributes to meat’s enhancement of nonheme iron absorption. J. Nutr. 2008, 138, 873–877. [Google Scholar] [PubMed]
- Reddy, M.B.; Hurrell, R.F.; Juillerat, M.A.; Cook, J.D. The influence of different protein sources on phytate inhibition of nonheme-iron absorption in humans. Am. J. Clin. Nutr. 1996, 63, 203–207. [Google Scholar] [PubMed]
- Monsen, E.R.; Hallberg, L.; Layrisse, M.; Hegsted, D.M.; Cook, J.D.; Mertz, W.; Finch, C.A. Estimation of available dietary iron. Am. J. Clin. Nutr. 1978, 31, 134–141. [Google Scholar] [PubMed]
- Hurrell, R.F.; Lynch, S.R.; Trinidad, T.P.; Dassenko, S.A.; Cook, J.D. Iron absorption in humans: Bovine serum albumin compared with beef muscle and egg white. Am. J. Clin. Nutr. 1988, 47, 102–107. [Google Scholar] [PubMed]
- Chabchoub-Ellouze, S.; Hamdaoui, M.H. Comparative effects of the addition of mutton or beef to a bean seed ragout on iron status in growing rats. Ann. Nutr. Metab. 2006, 50, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.B.; Cook, J.D. Assessment of dietary determinants of nonheme-iron absorption in humans and rats. Am. J. Clin. Nutr. 1991, 54, 723–728. [Google Scholar] [PubMed]
- Zhou, Y.; Alekel, D.L.; Dixon, P.M.; Messina, M.; Reddy, M.B. The effect of soy food intake on mineral status in premenopausal women. J. Womens Health 2011, 20, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Visentín, A.N.; Drago, S.R.; Osella, C.A.; de la Torre, M.A.; Sánchez, H.D.; González, R.J. Effect of the addition of soy flour and whey protein concentrate on bread quality and mineral dialyzability. Arch. Latinoam. Nutr. 2009, 59, 325–331. [Google Scholar] [PubMed]
- Hallberg, L.; Rossander, L. Improvement of iron nutrition in developing countries: Comparison of adding meat, soy protein, ascorbic acid, citric acid, and ferrous sulphate on iron absorption from a simple Latin American-type of meal. Am. J. Clin. Nutr. 1984, 39, 577–583. [Google Scholar] [PubMed]
- Pérez-Llamas, F.; Diepenmaat-Wolters, M.G.; Zamora, S. In vitro availability of iron and zinc: Effects of the type, concentration and fractions of digestion products of the protein. Br. J. Nutr. 1996, 76, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Akdogan, M.; Gultekin, F.; Yontem, M. Effect of Mentha piperita (Labiatae) and Mentha spicata (Labiatae) on iron absorption in rats. Toxicol. Ind. Health 2004, 20, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Hoekenga, O.A.; Kochian, L.V.; Glahn, R.P. High bioavailability iron maize (Zea mays L.) developed through molecular breeding provides more absorbable iron in vitro (Caco-2 model) and in vivo (Gallus gallus). Nutr. J. 2013, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P.; Knez, M.; Stangoulis, J.C. The effect of wheat prebiotics on the gut bacterial population and iron status of iron deficient broiler chickens. Nutr. J. 2014, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P. Intra-amniotic administration and dietary inulin affect the iron status and intestinal functionality of iron-deficient broiler chickens. Poult. Sci. 2012, 91, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Glahn, R.P.; Miller, D.D. Assessing potential effects of inulin and probiotic bacteria on Fe availability from common beans (Phaseolus vulgaris L.) to Caco-2 cells. J. Food Sci. 2009, 74, H40–H46. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Glahn, R.P.; Welch, R.M.; Lei, X.; Yasuda, K.; Miller, D.D. Dietary inulin affects the expression of intestinal enterocyte iron transporters, receptors and storage protein and alters the microbiota in the pig intestine. Br. J. Nutr. 2008, 99, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Nalepa, B.; Siemianowska, E.; Skibniewska, K.A. Influence of Bifidobacterium bifidum on release of minerals from bread with differing bran content. J. Toxicol. Environ. Health A 2012, 75, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bergqvist, S.W.; Andlid, T.; Sandberg, A.-S. Lactic acid fermentation stimulated iron absorption by Caco-2 cells is associated with increased soluble iron content in carrot juice. Br. J. Nutr. 2006, 96, 705–711. [Google Scholar] [PubMed]
- Ivaturi, R.; Kies, C. Mineral balances in humans as affected by fructose, high fructose corn syrup and sucrose. Plant Foods Hum. Nutr. Dordr. Neth. 1992, 42, 143–151. [Google Scholar] [CrossRef]
- Christides, T.; Sharp, P. Sugars increase non-heme iron bioavailability in human epithelial intestinal and liver cells. PLoS ONE 2013, 8, e83031. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.O.; Varnell, T.R. Effect of dietary carbohydrates on intestinal transport and accumulation of 59Fe++ in iron deficients rats. Int. J. Vitam. Nutr. Res. 1977, 47, 292–295. [Google Scholar] [PubMed]
- Della Lucia, C.M.; das Vaz Tostes, M.G.; Silveira, C.M.M.; Bordalo, L.A.; Rodrigues, F.C.; Pinheiro-Sant’Ana, H.M.; Martino, H.S.D.; Costa, N.M.B. Iron bioavailability in Wistar rats fed with fortified rice by Ultra Rice technology with or without addition of yacon flour (Smallanthus sonchifolius). Arch. Latinoam. Nutr. 2013, 63, 64–73. [Google Scholar] [PubMed]
- Wolf, B.W.; Firkins, J.L.; Zhang, X. Varying dietary concentrations of fructooligosaccharides affect apparent absorption and balance of minerals in growing rats. Nutr. Res. 1998, 18, 1791–1806. [Google Scholar] [CrossRef]
- Ohta, A.; Sakai, K.; Takasaki, M.; Uehara, M.; Tokunaga, T.; Adachi, T. Dietary heme iron does not prevent postgastrectomy anemia but fructooligosaccharides improve bioavailability of heme iron in rats. Int. J. Vitam. Nutr. Res. 1999, 69, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Ohtsuki, M.; Uehara, M.; Hosono, A.; Hirayama, M.; Adachi, T.; Hara, H. Dietary fructooligosaccharides prevent postgastrectomy anemia and osteopenia in rats. J. Nutr. 1998, 128, 485–490. [Google Scholar] [PubMed]
- Van den Heuvel, E.G.; Schaafsma, G.; Muys, T.; van Dokkum, W. Nondigestible oligosaccharides do not interfere with calcium and nonheme-iron absorption in young, healthy men. Am. J. Clin. Nutr. 1998, 67, 445–451. [Google Scholar] [PubMed]
- Huo, J.-S.; Yang, X.-G.; Piao, J.-H.; Gao, J.-Q.; Miao, H.; Yu, B.; Lu, C.-Q.; Chen, J.-S. NaFeEDTA fortified soy sauce showed higher iron absorption rate in Chinese females. Biomed. Environ. Sci. BES 2007, 20, 126–130. [Google Scholar] [PubMed]
- Hertrampf, E.; Olivares, M. Iron amino acid chelates. Int. J. Vitam. Nutr. Res. 2004, 74, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco-Valencia, R.A.; Encina, C.R.; Binaghi, M.J.; Greco, C.B.; Ronayne de Ferrer, P.A. Effects of roasting and boiling of quinoa, kiwicha and kañiwa on composition and availability of minerals in vitro. J. Sci. Food Agric. 2010, 90, 2068–2073. [Google Scholar] [CrossRef] [PubMed]
- Gahlawat, P.; Sehgal, S. In vitro starch and protein digestibility and iron availability in weaning foods as affected by processing methods. Plant Foods Hum. Nutr. Dordr. Neth. 1994, 45, 165–173. [Google Scholar] [CrossRef]
- Barakoti, L.; Bains, K. Effect of household processing on the in vitro bioavailability of iron in mungbean (Vigna radiata). Food Nutr. Bull. 2007, 28, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Hemalatha, S.; Platel, K.; Srinivasan, K. Influence of germination and fermentation on bioaccessibility of zinc and iron from food grains. Eur. J. Clin. Nutr. 2007, 61, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Nielsen, S.S.; Rodriguez-Burger, A.P.; Mason, A.C. Iron and zinc bioavailability in rats fed intrinsically labeled bean and bean-rice infant weaning food products. J. Agric. Food Chem. 2001, 49, 5063–5069. [Google Scholar] [CrossRef] [PubMed]
- Urizar Hernández, A.L.; Bressani, R. The effect of lime cooking of corn on phytic acid, calcium, total and ionizable iron content. Arch. Latinoam. Nutr. 1997, 47, 217–223. [Google Scholar] [PubMed]
- Sotelo, A.; González-Osnaya, L.; Sánchez-Chinchillas, A.; Trejo, A. Role of oxate, phytate, tannins and cooking on iron bioavailability from foods commonly consumed in Mexico. Int. J. Food Sci. Nutr. 2010, 61, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Glahn, R.P.; Miller, D.D. Bioaccessibility of phenols in common beans (Phaseolus vulgaris L.) and iron (Fe) availability to Caco-2 cells. J. Agric. Food Chem. 2008, 56, 10999–11005. [Google Scholar] [CrossRef] [PubMed]
- García-Casal, M.N.; Layrisse, M.; Solano, L.; Barón, M.A.; Arguello, F.; Llovera, D.; Ramírez, J.; Leets, I.; Tropper, E. Vitamin A and beta-carotene can improve nonheme iron absorption from rice, wheat and corn by humans. J. Nutr. 1998, 128, 646–650. [Google Scholar] [PubMed]
- El-Hawary, Z.; El-Shobaki, F.A.; Saleh, N.; Morcos, S.R. Intestinal absorption of iron alone and in combination with authentic or natural vitamin C and carotene. Z. Für Ernährungswissenschaft 1976, 15, 327–332. [Google Scholar] [CrossRef]
- Kim, E.-Y.; Ham, S.-K.; Bradke, D.; Ma, Q.; Han, O. Ascorbic acid offsets the inhibitory effect of bioactive dietary polyphenolic compounds on transepithelial iron transport in Caco-2 intestinal cells. J. Nutr. 2011, 141, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, L.; Brune, M.; Rossander, L. Iron absorption in man: Ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr. 1989, 49, 140–144. [Google Scholar] [PubMed]
- Reddy, M.B.; Hurrell, R.F.; Cook, J.D. Estimation of nonheme-iron bioavailability from meal composition. Am. J. Clin. Nutr. 2000, 71, 937–943. [Google Scholar] [PubMed]
- Péneau, S.; Dauchet, L.; Vergnaud, A.-C.; Estaquio, C.; Kesse-Guyot, E.; Bertrais, S.; Latino-Martel, P.; Hercberg, S.; Galan, P. Relationship between iron status and dietary fruit and vegetables based on their vitamin C and fiber content. Am. J. Clin. Nutr. 2008, 87, 1298–1305. [Google Scholar] [PubMed]
- Ball, M.J.; Bartlett, M.A. Dietary intake and iron status of Australian vegetarian women. Am. J. Clin. Nutr. 1999, 70, 353–358. [Google Scholar] [PubMed]
- Pirzad, A.; Darvishzadeh, R.; Bernousi, I.; Hassani, A.; Sivritepe, N. Influence of Water Deficit on Iron and Zinc Uptake by Matricaria chamomilla L. Chil. J. Agric. Res. 2012, 72, 232–236. [Google Scholar] [CrossRef]
- Rebey, I.B.; Zakhama, N.; Karoui, I.J.; Marzouk, B. Polyphenol composition and antioxidant activity of cumin (Cuminum cyminum L.) seed extract under drought. J. Food Sci. 2012, 77, C734–C739. [Google Scholar] [CrossRef] [PubMed]
- Germanò, M.P.; Cacciola, F.; Donato, P.; Dugo, P.; Certo, G.; D’Angelo, V.; Mondello, L.; Rapisarda, A. Betula pendula leaves: Polyphenolic characterization and potential innovative use in skin whitening products. Fitoterapia 2012, 83, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Rabiei, K.; Bekhradnia, S.; Nabavi, S.M.; Nabavi, S.F.; Ebrahimzadeh, M.A. Antioxidant activity of polyphenol and ultrasonic extracts from fruits of Crataegus pentagyna subsp. elburensis. Nat. Prod. Res. 2012, 26, 2353–2357. [Google Scholar] [CrossRef] [PubMed]
- Beevi, S.S.; Mangamoori, L.N.; Gowda, B.B. Polyphenolics profile and antioxidant properties of Raphanus sativus L. Nat. Prod. Res. 2012, 26, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Beevi, S.S.; Narasu, M.L.; Gowda, B.B. Polyphenolics profile, antioxidant and radical scavenging activity of leaves and stem of Raphanus sativus L. Plant Foods Hum. Nutr. Dordr. Neth. 2010, 65, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; McClung, A.M.; Chen, M.-H. Phytochemicals and antioxidant capacities in rice brans of different color. J. Food Sci. 2011, 76, C117–C126. [Google Scholar] [CrossRef] [PubMed]
- Vladimir-Knežević, S.; Blažeković, B.; Štefan, M.B.; Alegro, A.; Koszegi, T.; Petrik, J. Antioxidant activities and polyphenolic contents of three selected Micromeria species from Croatia. Mol. Basel Switz. 2011, 16, 1454–1470. [Google Scholar] [CrossRef] [PubMed]
- Blazeković, B.; Vladimir-Knezević, S.; Brantner, A.; Stefan, M.B. Evaluation of antioxidant potential of Lavandula x intermedia Emeric ex Loisel. “Budrovka”: A comparative study with L. angustifolia Mill. Mol. Basel Switz. 2010, 15, 5971–5987. [Google Scholar] [CrossRef] [PubMed]
- Tuntipopipat, S.; Zeder, C.; Siriprapa, P.; Charoenkiatkul, S. Inhibitory effects of spices and herbs on iron availability. Int. J. Food Sci. Nutr. 2009, 60 (Suppl. 1), 43–55. [Google Scholar] [CrossRef] [PubMed]
- Nnam, N.M.; Onyeke, N.G. Chemical composition of two varieties of sorrel (Hibiscus sabdariffa L.), calyces and the drinks made from them. Plant Foods Hum. Nutr. 2003, 58, 1–7. [Google Scholar] [CrossRef]
- García-Casal, M.N.; Ramírez, J.; Leets, I.; Pereira, A.C.; Quiroga, M.F. Antioxidant capacity, polyphenol content and iron bioavailability from algae (Ulva sp., Sargassum sp. and Porphyra sp.) in human subjects. Br. J. Nutr. 2009, 101, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Ariza-Nieto, M.; Blair, M.W.; Welch, R.M.; Glahn, R.P. Screening of iron bioavailability patterns in eight bean (Phaseolus vulgaris L.) genotypes using the Caco-2 cell in vitro model. J. Agric. Food Chem. 2007, 55, 7950–7956. [Google Scholar] [CrossRef] [PubMed]
- Gillooly, M.; Bothwell, T.H.; Charlton, R.W.; Torrance, J.D.; Bezwoda, W.R.; MacPhail, A.P.; Derman, D.P.; Novelli, L.; Morrall, P.; Mayet, F. Factors affecting the absorption of iron from cereals. Br. J. Nutr. 1984, 51, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Glahn, R.P.; Cheng, Z.; Welch, R.M.; Gregorio, G.B. Comparison of iron bioavailability from 15 rice genotypes: Studies using an in vitro digestion/caco-2 cell culture model. J. Agric. Food Chem. 2002, 50, 3586–3591. [Google Scholar] [CrossRef] [PubMed]
- Matuschek, E.; Towo, E.; Svanberg, U. Oxidation of polyphenols in phytate-reduced high-tannin cereals: Effect on different phenolic groups and on in vitro accessible iron. J. Agric. Food Chem. 2001, 49, 5630–5638. [Google Scholar] [CrossRef] [PubMed]
- Jood, S.; Kalra, S. Chemical composition and nutritional characteristics of some hull less and hulled barley cultivars grown in India. Nahrung 2001, 45, 35–39. [Google Scholar] [CrossRef]
- Macfarlane, B.J.; Bezwoda, W.R.; Bothwell, T.H.; Baynes, R.D.; Bothwell, J.E.; MacPhail, A.P.; Lamparelli, R.D.; Mayet, F. Inhibitory effect of nuts on iron absorption. Am. J. Clin. Nutr. 1988, 47, 270–274. [Google Scholar] [PubMed]
- Zaida, F.; Bureau, F.; Guyot, S.; Sedki, A.; Lekouch, N.; Arhan, P.; Bouglé, D. Iron availability and consumption of tea, vervain and mint during weaning in Morocco. Ann. Nutr. Metab. 2006, 50, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Sehgal, S. Effect of domestic processing and cooking methods on total, hcl extractable iron and in vitro availability of iron in spinach and amaranth leaves. Nutr. Health 2002, 16, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, D.R.; Salim, K.M.; Srivastava, A.; Singh, L.B.; Srivastava, R.C. Estimation of proximate composition, micronutrients and phytochemical compounds in traditional vegetables from Andaman and Nicobar Islands. Int. J. Food Sci. Nutr. 2011, 62, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Adedapo, A.; Jimoh, F.; Afolayan, A. Comparison of the nutritive value and biological activities of the acetone, methanol and water extracts of the leaves of Bidens pilosa and Chenopodium album. Acta Pol. Pharm. 2011, 68, 83–92. [Google Scholar] [PubMed]
- Jimoh, F.; Adedapo, A.; Aliero, A.; Afolayan, A. Polyphenolic and biological activities of leaves extracts of Argemone subfusiformis (Papaveraceae) and Urtica urens (Urticaceae). Rev. Biol. Trop. 2010, 58, 1517–1531. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Lee, M.-K.; Heo, B.-G.; Ham, K.-S.; Kang, S.-G.; Cho, J.-Y.; Gorinstein, S. Comparison of the nutrient and chemical contents of traditional Korean Chungtaejeon and green teas. Plant Foods Hum. Nutr. Dordr. Neth. 2010, 65, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Jimoh, F.O.; Adedapo, A.A.; Afolayan, A.J. Comparison of the nutritional value and biological activities of the acetone, methanol and water extracts of the leaves of Solanum nigrum and Leonotis leonorus. Food Chem. Toxicol. 2010, 48, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Butnariu, M.; Bostan, C.; Samfira, I. Determination of mineral contents and antioxidant activity in some plants that contain allelochemicals of Banat region. Stud. Univ. “Vasile Goldiş” Ser. Ştiinţ. Vieţii 2012, 22, 95–100. [Google Scholar]
- Ebrahimzadeh, M.A.; Nabavi, S.M.; Nabavi, S.F. Correlation between the in vitro iron chelating activity and poly phenol and flavonoid contents of some medicinal plants. Pak. J. Biol. Sci. PJBS 2009, 12, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Schricker, B.R.; Miller, D.D.; Rasmussen, R.R.; van Campen, D. A comparison of in vivo and in vitro methods for determining availability of iron from meals. Am. J. Clin. Nutr. 1981, 34, 2257–2263. [Google Scholar] [PubMed]
- Lynch, S. The precision of in vitro methods and algorithms for predicting the bioavailability of dietary iron. Int. J. Vitam. Nutr. Res. 2005, 75, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, A.; Chenoweth, W.A.; Kelly, J.F.; Agee, K.M. Iron bioavailability fromAmaranthus species: 2—Evaluation using haemoglobin repletion in anaemic rats. J. Sci. Food Agric. 1998, 78, 274–280. [Google Scholar] [CrossRef]
- Martinez Meyer, M.R.; Rojas, A.; Santanen, A.; Stoddard, F.L. Content of zinc, iron and their absorption inhibitors in Nicaraguan common beans (Phaseolus vulgaris L.). Food Chem. 2013, 136, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, R.P.; Singh, N.P. Analysis of macro and micro nutrients in some Indian medicinal herbs grown in Jaunpur (u.p.) soil. Nat. Sci. 2011, 3, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Cercamondi, C.I.; Icard-Verniere, C.; Egli, I.M.; Vernay, M.; Hama, F.; Brouwer, I.D.; Zeder, C.; Berger, J.; Hurrell, R.F.; Mouquet-Rivier, C. A Higher Proportion of Iron-Rich Leafy Vegetables in a Typical Burkinabe Maize Meal Does Not Increase the Amount of Iron Absorbed in Young Women. J. Nutr. 2014, 144, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Alpers, D.H.; Stenson, W.F.; Taylor, B.E.; Bier, D.M. Manual of Nutritional Therapeutics, Spiral Manua, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008; p. 63. [Google Scholar]
- Nelson, R.E.; Ferruzzi, M.G. Synthesis and bioaccessibility of Fe-pheophytin derivatives from crude spinach extract. J. Food Sci. 2008, 73, H86–H91. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancuceanu, R.; Dinu, M.; Hovaneţ, M.V.; Anghel, A.I.; Popescu, C.V.; Negreş, S. A Survey of Plant Iron Content—A Semi-Systematic Review. Nutrients 2015, 7, 10320-10351. https://doi.org/10.3390/nu7125535
Ancuceanu R, Dinu M, Hovaneţ MV, Anghel AI, Popescu CV, Negreş S. A Survey of Plant Iron Content—A Semi-Systematic Review. Nutrients. 2015; 7(12):10320-10351. https://doi.org/10.3390/nu7125535
Chicago/Turabian StyleAncuceanu, Robert, Mihaela Dinu, Marilena Viorica Hovaneţ, Adriana Iuliana Anghel, Carmen Violeta Popescu, and Simona Negreş. 2015. "A Survey of Plant Iron Content—A Semi-Systematic Review" Nutrients 7, no. 12: 10320-10351. https://doi.org/10.3390/nu7125535
APA StyleAncuceanu, R., Dinu, M., Hovaneţ, M. V., Anghel, A. I., Popescu, C. V., & Negreş, S. (2015). A Survey of Plant Iron Content—A Semi-Systematic Review. Nutrients, 7(12), 10320-10351. https://doi.org/10.3390/nu7125535