Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils

Abstract
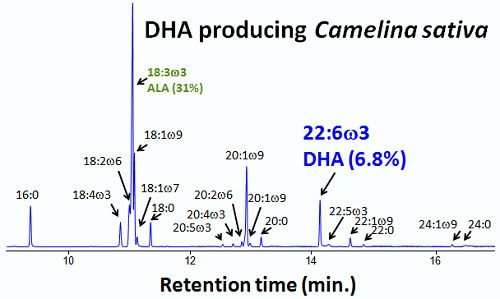
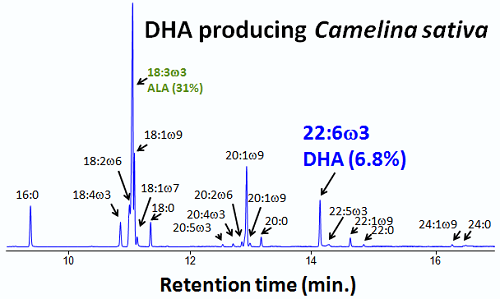
:
1. Introduction
2. Experimental Section
2.1. Lipid Extraction
2.2. Lipid Class Analysis
2.3. Separation of TAG, Glycolipid and Phospholipid Fractions from the CM Extracts
2.4. Fatty Acid Methyl Ester Preparation
2.5. Sterol and Fatty Alcohol Derivatization
2.6. GC and GC-MS Analysis
3. Results
3.1. Total Lipid Content

{kind=link}
| Extract | DHA camelina | Control camelina |
|---|---|---|
| hexane | 31.1 | 38.1 |
| chloroform-methanol 1 | 4.8 | 2.4 |
| residual lipid 2 | 0.3 | 0.4 |
| Total lipid | 36.2 | 40.9 |
3.2. Lipid Class Analysis
| Lipid class | DHA camelina | Control camelina | ||
|---|---|---|---|---|
| Hexane | Chloroform-methanol | Hexane | Chloroform-methanol | |
| SE/WE/HC 1 | 1.0 | 1.4 | 1.0 | 1.4 |
| TAG | 95.6 | 44.2 | 96.0 | 13.1 |
| FFA | 0.9 | 1.3 | 0.8 | 1.4 |
| UN 2 | 0.9 | 1.1 | 0.8 | 1.2 |
| ST | 0.5 | 0.7 | 0.4 | 0.4 |
| MAG | 0.7 | 1.1 | 0.8 | 6.2 |
| PL | 0.3 | 50.3 | 0.3 | 76.3 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 |
| Phospholipid | DHA camelina | Control camelina |
|---|---|---|
| PA | 2.1 | 4.7 |
| UN 1 | 5.7 | 2.2 |
| UN 2 | - | 1.1 |
| UN 3 | - | 0.6 |
| PE | 13.2 | 6.8 |
| PS | 1.2 | 1.4 |
| PC 1 + PI | 69.5 | 78.9 |
| UN 4 | 4.8 | 3.6 |
| UN 5 | 3.4 | 1.6 |
| Total | 100.0 | 100.0 |
| DHA camelina | Control camelina | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hexane | Chloroform-methanol | Meal | Hexane | Chloroform-methanol | Meal | |||||||
| Fatty acid | TAG 1 | Total 2 | TAG 3 | GL 3 | PL 3 | Residue 4 | TAG 1 | Total 2 | TAG 3 | GL 3 | PL 3 | Residue 4 |
| 16:1ω7 | 0.1 | 0.2 | 0.1 | 0.2 | 0.1 | 0.2 | 0.1 | 0.2 | 0.2 | - | - | 0.3 |
| 16:0 | 6.2 | 12.8 | 6.8 | 21.3 | 19.4 | 10.4 | 6.7 | 12.8 | 7.8 | 29.6 | 13.7 | 10.3 |
| 18:4ω3 | 3.7 | 3.3 | 3.4 | 2.1 | 2.9 | 3.6 | - | - | - | - | - | - |
| 18:2ω6 | 7.1 | 3.9 | 8.8 | 7.2 | 3.7 | 8.8 | 22.2 | 28.4 | 29.4 | 20.8 | 29.3 | 27.9 |
| 18:3ω3 | 41.9 | 50.3 | 39.9 | 38.6 | 54.1 | 38.9 | 32.0 | 20.6 | 19.7 | 13.0 | 12.3 | 20.0 |
| 18:1ω9 | 11.1 | 4.7 | 9.6 | 7.2 | 2.8 | 8.1 | 14.0 | 25.4 | 13.3 | 14.7 | 35.7 | 14.3 |
| 18:1ω7 | 1.4 | 2.3 | 2.1 | 3.7 | 3.4 | 2.8 | 1.0 | 1.5 | 2.2 | 4.0 | 2.8 | 2.2 |
| 18:0 | 3.2 | 4.0 | 3.0 | 4.5 | 5.7 | 3.1 | 3.0 | 2.7 | 2.9 | 5.7 | 3.6 | 2.7 |
| 20:5ω3 | 0.4 | 0.2 | 0.3 | - | - | 0.3 | - | - | - | - | - | - |
| 20:4ω3 | 0.4 | 0.4 | 0.4 | - | 0.2 | 0.3 | - | - | - | - | - | - |
| 20:2ω6 | 0.7 | 0.7 | 0.8 | 0.6 | 0.4 | 0.7 | 1.8 | 0.8 | 2.1 | 1.2 | - | 1.8 |
| 20:3ω3 | 0.8 | 1.2 | 0.9 | 0.6 | 1.3 | 0.5 | 0.9 | 0.3 | - | - | - | 0.4 |
| 20:1ω9/11 | 11.6 | 6.1 | 10.9 | 5.1 | 1.3 | 8.4 | 12.5 | 3.0 | 11.1 | 4.2 | 1.7 | 9.4 |
| 20:1ω7 | 0.6 | 0.8 | 1.4 | 0.6 | 0.2 | 1.1 | 0.6 | 0.6 | 2.6 | 1.3 | - | 1.8 |
| 20:0 | 1.3 | 0.8 | 1.4 | 0.6 | 0.1 | 1.4 | 1.5 | 0.7 | 2.3 | 1.4 | - | 1.8 |
| 22:6ω3 | 6.8 | 4.2 | 6.1 | 3.0 | 1.6 | 5.4 | - | - | - | - | - | - |
| 22:5ω3 | 0.3 | 1.1 | 0.4 | 0.6 | 1.4 | 0.3 | - | - | - | - | - | - |
| 22:1ω9 | 1.3 | 1.0 | 1.8 | 0.6 | 0.1 | 1.5 | 2.7 | 0.7 | 3.6 | 0.9 | - | 2.9 |
| 22:0 | 0.3 | 0.2 | 0.3 | 0.6 | 0.1 | 0.7 | 0.3 | 0.2 | 0.7 | 0.8 | - | 0.8 |
| 24:1ω9 | 0.3 | 0.4 | 0.4 | 0.6 | 0.3 | 0.6 | 0.3 | 0.6 | 0.7 | 0.9 | 0.5 | 1.0 |
| 24:0 | 0.1 | 0.4 | 0.2 | 0.9 | 0.4 | 1.1 | 0.1 | 0.4 | 0.5 | 1.4 | 0.4 | 1.3 |
| Others 5 | 0.4 | 1.0 | 1.0 | 1.4 | 0.5 | 1.8 | 0.3 | 1.1 | 0.9 | 0.1 | - | 1.1 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
3.3. Fatty Acid Composition
3.4. Sterol Composition
| DHA camelina | Control camelina | |||
|---|---|---|---|---|
| Sterols | Hexane | CM 1 | Hexane | CM 1 |
| 24-dehydrocholesterol | 0.8 | 1.8 | 0.5 | 1.4 |
| cholesterol | 5.7 | 7.6 | 4.7 | 7.2 |
| brassicasterol | 4.4 | 6.5 | 1.9 | 4.2 |
| cholest-7-en-3β-ol | 0.2 | 0.5 | 0.3 | 0.4 |
| campesterol | 24.5 | 20.8 | 25.7 | 21.7 |
| campestanol | 0.4 | 1.1 | 0.4 | 0.9 |
| stigmasterol | 1.0 | 2.6 | 0.5 | 1.6 |
| sitosterol | 54.3 | 43.7 | 53.8 | 42.9 |
| ∆5-avenasterol (isofucosterol) | 4.2 | 5.2 | 4.7 | 5.5 |
| Sum | 95.5 | 89.6 | 92.6 | 85.9 |
| Others | ||||
| UN1 C28 1db | 0.6 | 1.2 | 0.7 | 1.2 |
| UN2 C29 1db | 1.2 | 2.0 | 1.2 | 2.4 |
| UN3 C29 2db | 0.9 | 1.8 | 1.3 | 2.4 |
| UN4 C28 1db | 0.3 | 0.9 | 0.6 | 1.1 |
| UN5 C30 2db | 1.2 | 1.8 | 1.4 | 1.8 |
| UN6 C29 1db + C30 2db | 0.3 | 2.7 | 2.2 | 5.2 |
| Sum | 4.5 | 10.4 | 7.4 | 14.1 |
| Total | 100 | 100 | 100 | 100 |
3.5. Fatty Alcohol Composition
| Fatty alcohols | DHA camelina | Control camelina | ||
|---|---|---|---|---|
| Hexane | CM 1 | Hexane | CM 1 | |
| iso-16:0 | 0.8 | 1.9 | 1.5 | 1.7 |
| 16:0 | 5.7 | 13.8 | 6.1 | 12.2 |
| iso-17:0 | 16.4 | 37.1 | 23.5 | 38.6 |
| 17:0 | 3.8 | 0.3 | 5.7 | 1.0 |
| iso-18:0 | 6.6 | 11.8 | 8.3 | 13.5 |
| 18:1 | 2.8 | 8.0 | 2.5 | 5.1 |
| 18:0 | 9.3 | 7.9 | 7.4 | 6.4 |
| Phytol | 47.1 | 9.1 | 37.4 | 11.7 |
| 19:0 | 4.5 | 5.6 | 5.1 | 6.7 |
| iso-20:0 | 0.0 | 0.2 | 0.0 | 0.2 |
| 20:1 | 1.0 | 0.9 | 0.5 | 0.6 |
| 20:0 | 1.3 | 1.7 | 1.2 | 1.1 |
| iso-22:0 | 0.0 | 0.2 | 0.0 | 0.2 |
| 22:1 | 0.0 | 0.5 | 0.0 | 0.3 |
| 22:0 | 0.8 | 1.0 | 0.7 | 0.5 |
| Sum | 100 | 100 | 100 | 100 |
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Galli, C.; Calder, P.C. Effects of fat and fatty acid intake on inflammatory and immune responses: A critical review. Ann. Nutr. Metab. 2009, 55, 123–139. [Google Scholar] [CrossRef]
- Giles, G.E.; Mahoney, C.R.; Kanarek, R.B. Omega-3 fatty acids influence mood in healthy and depressed individuals. Nutr. Rev. 2013, 71, 727–741. [Google Scholar] [CrossRef]
- Weichselbaum, E.; Coe, S.; Buttriss, J.; Stanner, S. Fish in the diet: A review. Nutr. Bull. 2013, 38, 128–177. [Google Scholar] [CrossRef]
- Ryan, J.M.; Rice, G.E.; Mitchell, M.D. The role of gangliosides in brain development and the potential benefits of perinatal supplementation. Nutr. Res. 2013, 33, 877–887. [Google Scholar] [CrossRef]
- Simon, E.; Bardet, B.; Gregoire, S.; Acar, N.; Bron, A.M.; Creuzot-Garcher, C.R.; Bretillon, L. Decreasing dietary linoleic acid promotes long chain omega-3 fatty acid incorporation into rat retina and modifies gene expression. Exp. Eye Res. 2011, 93, 628–635. [Google Scholar] [CrossRef]
- Lund, I.; Steenfeldt, S.J. The effects of dietary long-chain essential fatty acids on growth and stress tolerance in pikeperch larvae (Sander lucioperca L.). Aquacul. Nutr. 2011, 17, 191–199. [Google Scholar] [CrossRef]
- Tani, S.; Nagao, K.; Matsumoto, M.; Hirayama, A. Highly purified eicosapentaenoic acid may increase low-density lipoprotein particle size by improving triglyceride metabolism in patients with hypertriglyceridemia. Circul. J. 2013, 77, 2349–2357. [Google Scholar] [CrossRef]
- Rechenberg, K.; Humphries, D. Nutritional interventions in depression and perinatal depression. Yale J. Biol. Med. 2013, 86, 127–137. [Google Scholar]
- Weintraub, H. Update on marine omega-3 fatty acids: Management of dyslipidemia and current omega-3 treatment options. Atherosclerosis 2013, 230, 381–389. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Lim, D.K.Y.; Timmins, M.; Vernen, F.; Yan, L.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11. [Google Scholar] [CrossRef]
- Monroig, O.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs 2013, 11, 3998–4018. [Google Scholar] [CrossRef]
- Mansour, M.P.; Frampton, D.M.F.; Nichols, P.D.; Volkman, J.K.; Blackburn, S.I. Lipid and fatty acid yield of nine stationary-phase microalgae: Applications and unusual C24–C28 polyunsaturated fatty acids. J. Appl. Phycol. 2005, 17, 287–300. [Google Scholar] [CrossRef]
- Mansour, M.P.; Volkman, J.K.; Blackburn, S.I. The effect of growth phase on the lipid class, fatty acid and sterol composition in the marine dinoflagellate, Gymnodinium sp. in batch culture. Phytochemistry 2003, 63, 145–153. [Google Scholar] [CrossRef]
- Mansour, M.P.; Volkman, J.K.; Holdsworth, D.G.; Jackson, A.E.; Blackburn, S.I. Very-long-chain (C28) highly unsaturated fatty acids in marine dinoflagellates. Phytochemistry 1999, 50, 541–548. [Google Scholar] [CrossRef]
- Mansour, M.P.; Volkman, J.K.; Jackson, A.E.; Blackburn, S.I. The fatty acid and sterol composition of five marine dinoflagellates. J. Phycol. 1999, 35, 710–720. [Google Scholar]
- Wood, C.C.; Petrie, J.R.; Shrestha, P.; Mansour, M.P.; Nichols, P.D.; Green, A.G.; Singh, S.P. A leaf-based assay using interchangeable design principles to rapidly assemble multistep recombinant pathways. Plant Biotechnol. J. 2009, 7, 914–924. [Google Scholar] [CrossRef]
- Petrie, J.R.; Shrestha, P.; Zhou, X.-R.; Mansour, M.P.; Liu, Q.; Belide, S.; Nichols, P.D.; Singh, S.P. Metabolic engineering plant seeds with fish oil-like levels of DHA. PLoS One 2012, 7, e49165. [Google Scholar] [CrossRef]
- Petrie, J.R.; Nichols, P.D.; Devine, M.; Singh, S.P. Engineered oilseed crops with fish oil DHA levels. Int. News Fats Oils Relat. Mater. 2013, 24, 648–652. [Google Scholar]
- Budin, J.T.; Breene, W.M.; Putnam, D.H. Some compositional properties of camelina (Camelina sativa L. Crantz) seeds and oils. J. Am. Oil Chem. Soc. 1995, 72, 309–315. [Google Scholar] [CrossRef]
- Petrie, J.R.; Shrestha, P.; Belide, S.; Kennedy, Y.; Lester, G.; Liu, Q.; Divi, U.K.; Mulder, R.J.; Mansour, M.P.; Nichols, P.D.; et al. Metabolic engineering Camelina sativa with fish oil-like levels of DHA. PLoS One 2014, 9, e85061. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Avalli, A.; Contarini, G. Determination of phospholipids in dairy products by SPE/HPLU/ELSD. J. Chromatogr. A 2005, 1071, 185–190. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar]
- Angelini, L.G.; Moscheni, E.; Colonna, G.; Belloni, P.; Bonari, E. Variation in agronomic characteristics and seed oil composition of new oilseed crops in central Italy. Ind. Crops Prod. 1997, 6, 313–323. [Google Scholar] [CrossRef]
- Zubr, J. Qualitative variation of Camelina sativa seed from different locations. Ind. Crops Prod. 2003, 17, 161–169. [Google Scholar] [CrossRef]
- Vollmann, J.; Moritz, T.; Kargl, C.; Baumgartner, S.; Wagentristl, H. Agronomic evaluation of camelina genotypes selected for seed quality characteristics. Ind. Crops Prod. 2007, 26, 270–277. [Google Scholar] [CrossRef]
- Abramovič, H.; Butinar, B.; Nikolič, V. Changes occurring in phenolic content, tocopherol composition and oxidative stability of Camelina sativa oil during storage. Food Chem. 2007, 104, 903–909. [Google Scholar] [CrossRef]
- Shukla, V.K.S.; Dutta, P.C.; Artz, W.E. Camelina oil and its unusual cholesterol content. J. Am. Oil Chem. Soc. 2002, 79, 965–969. [Google Scholar] [CrossRef]
- Schwartz, H.; Ilainen, V.; Pfironen, V.; Lampi, A.-M. Tocopherol, tocotrienol and plant sterol contents of vegetable oils and industrial fats. J. Food Compos. Anal. 2008, 21, 152–161. [Google Scholar] [CrossRef]
- Steinke, G.; Weitkamp, P.; Klein, E.; Mukherjee, K.D. High-yield preparation of wax esters via lipase-catalyzed esterification using fatty acids and alcohols from crambe and camelina oils. J. Agric. Food Chem. 2001, 49, 647–651. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mansour, M.P.; Shrestha, P.; Belide, S.; Petrie, J.R.; Nichols, P.D.; Singh, S.P. Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils. Nutrients 2014, 6, 776-789. https://doi.org/10.3390/nu6020776
Mansour MP, Shrestha P, Belide S, Petrie JR, Nichols PD, Singh SP. Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils. Nutrients. 2014; 6(2):776-789. https://doi.org/10.3390/nu6020776
Chicago/Turabian StyleMansour, Maged P., Pushkar Shrestha, Srinivas Belide, James R. Petrie, Peter D. Nichols, and Surinder P. Singh. 2014. "Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils" Nutrients 6, no. 2: 776-789. https://doi.org/10.3390/nu6020776
APA StyleMansour, M. P., Shrestha, P., Belide, S., Petrie, J. R., Nichols, P. D., & Singh, S. P. (2014). Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils. Nutrients, 6(2), 776-789. https://doi.org/10.3390/nu6020776