Biological Activities and Bioavailability of Mangosteen Xanthones: A Critical Review of the Current Evidence

Abstract
:
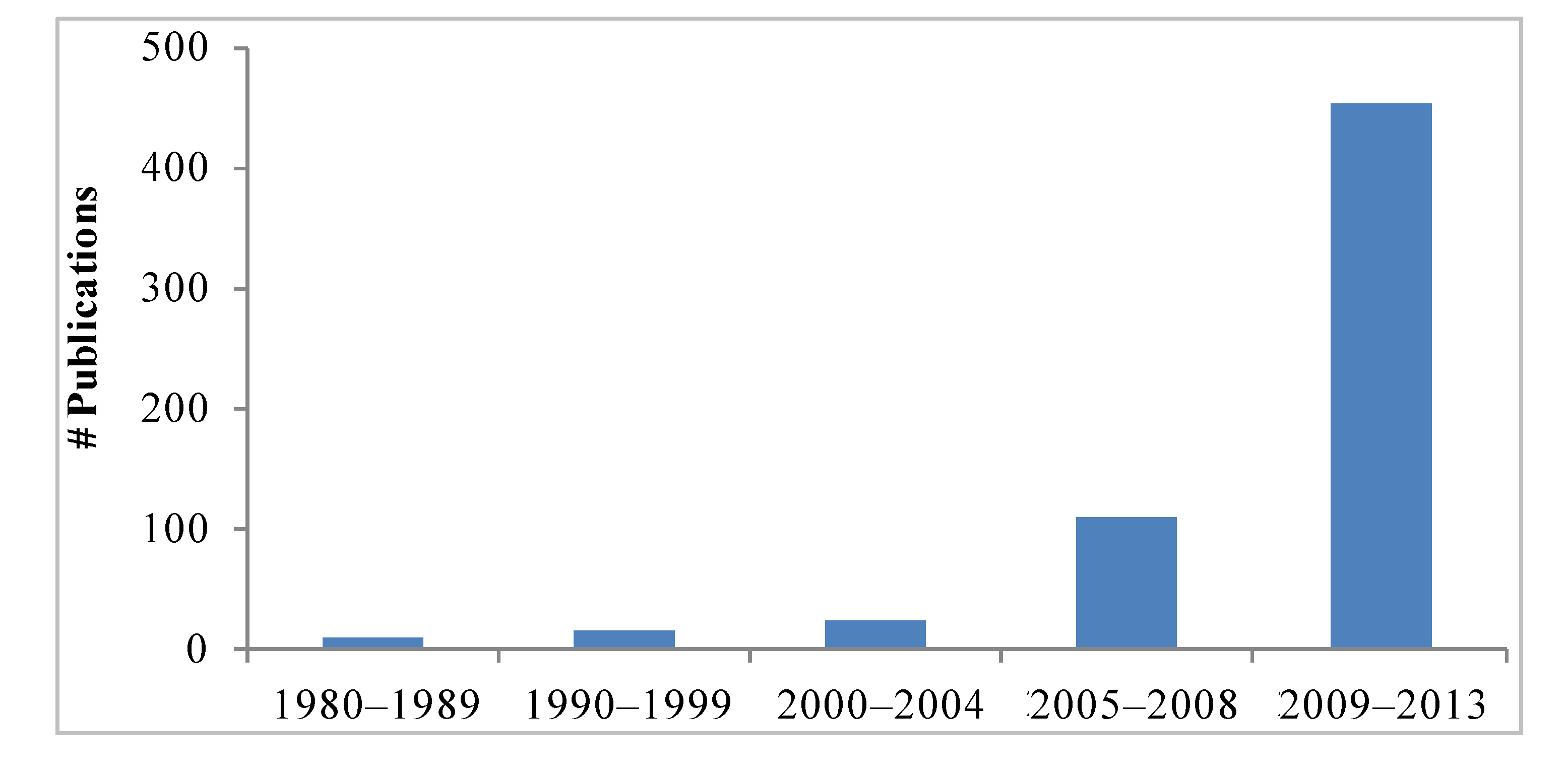
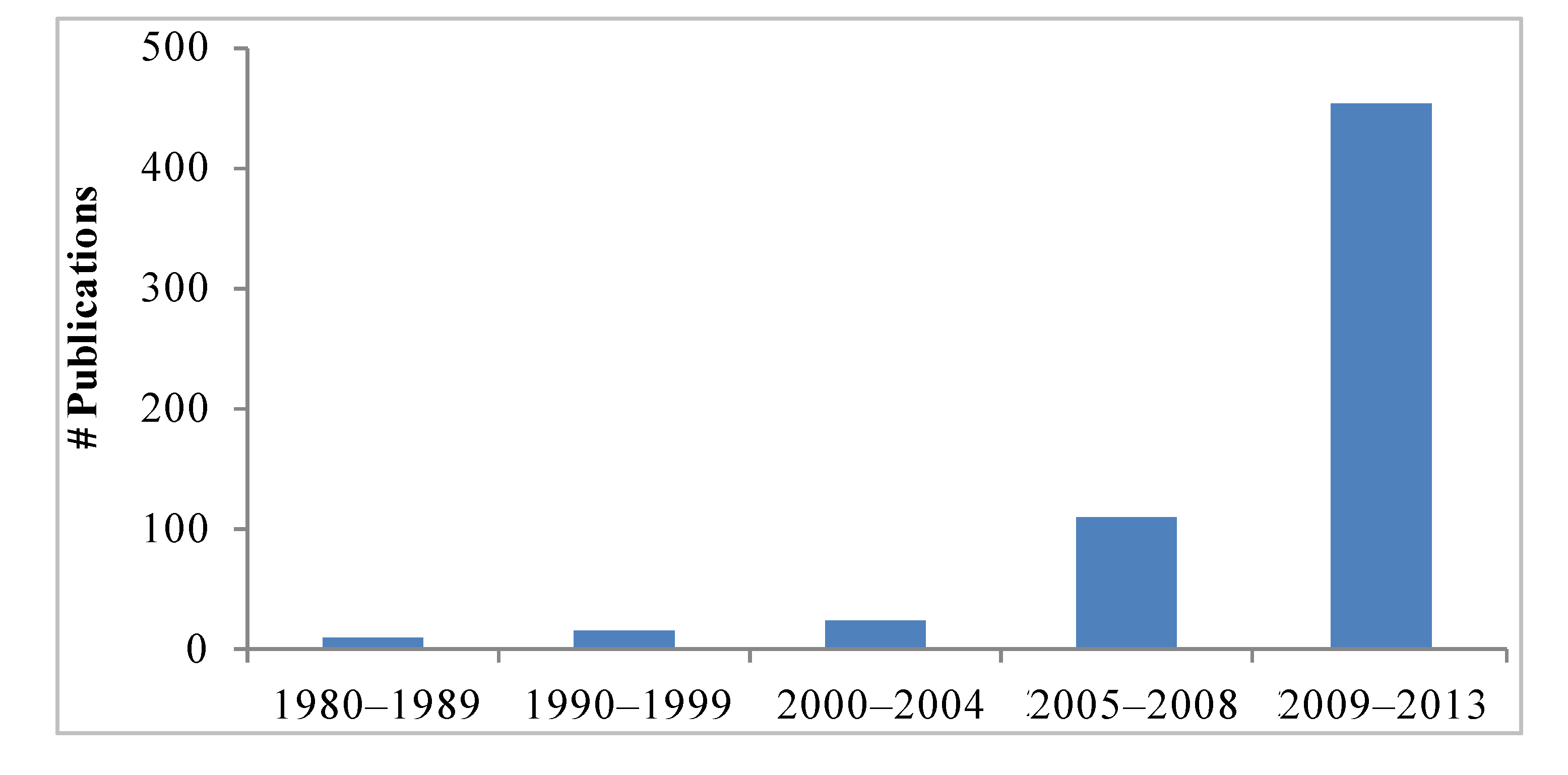
1. Introduction
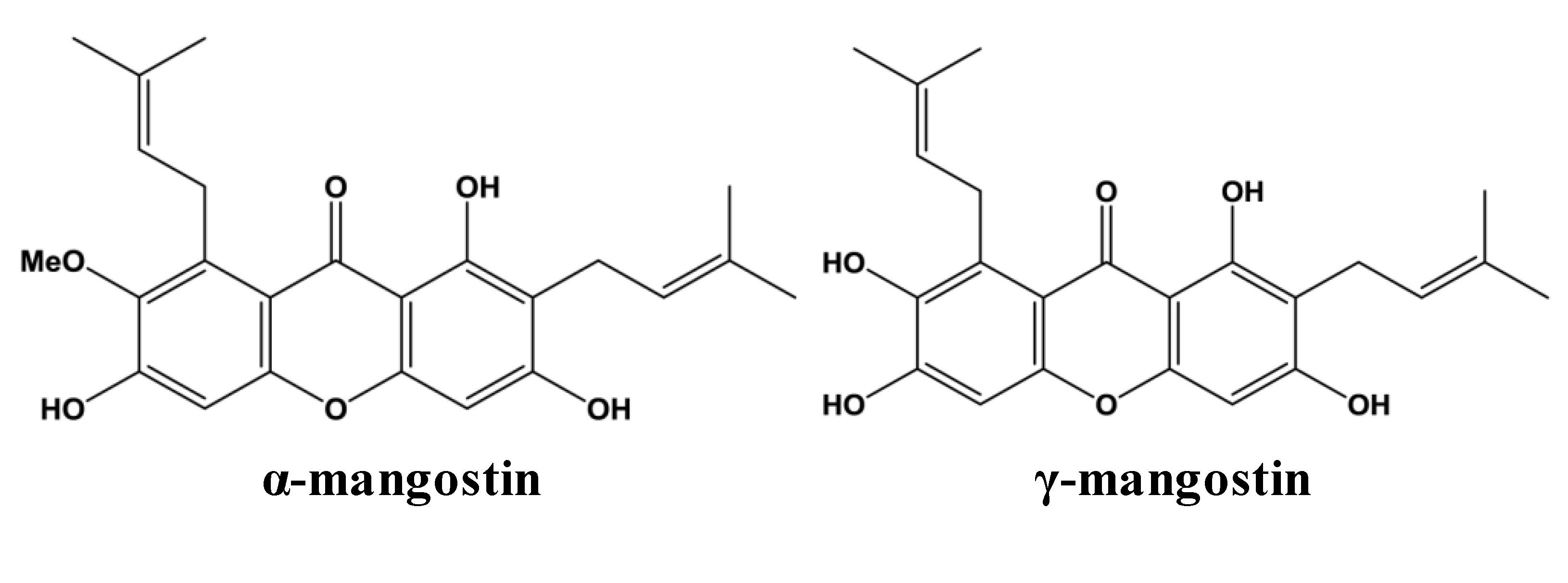
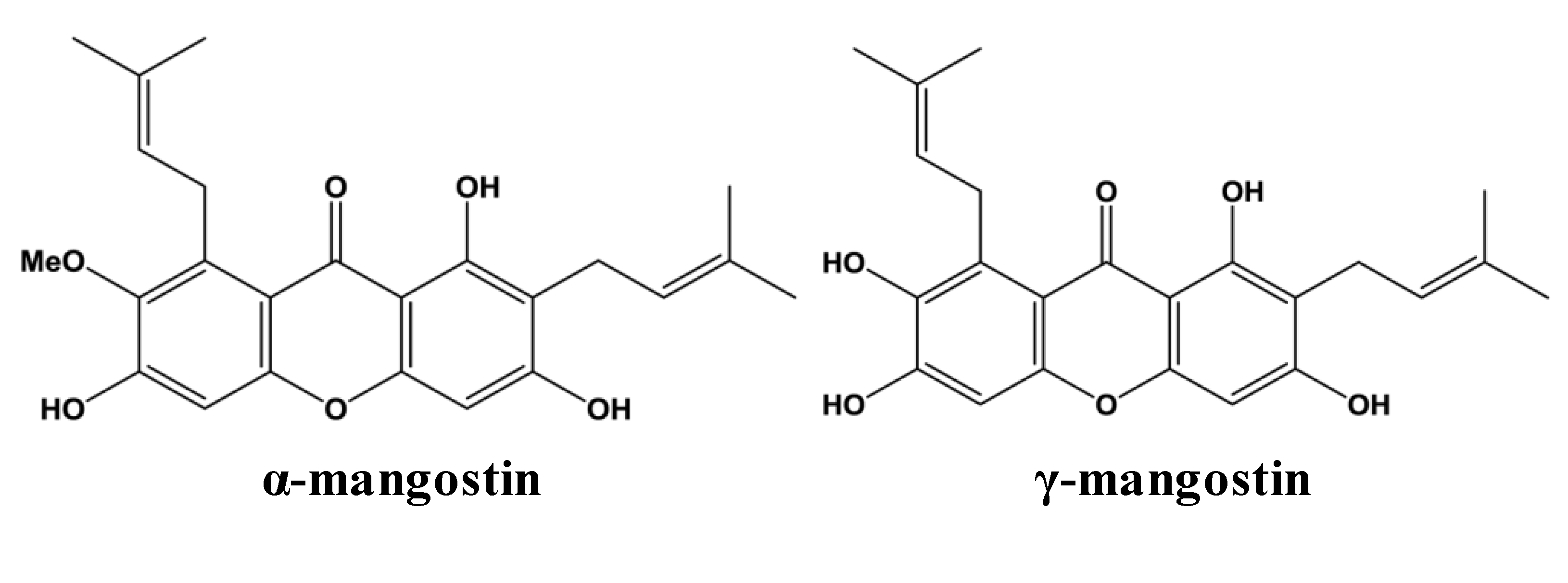
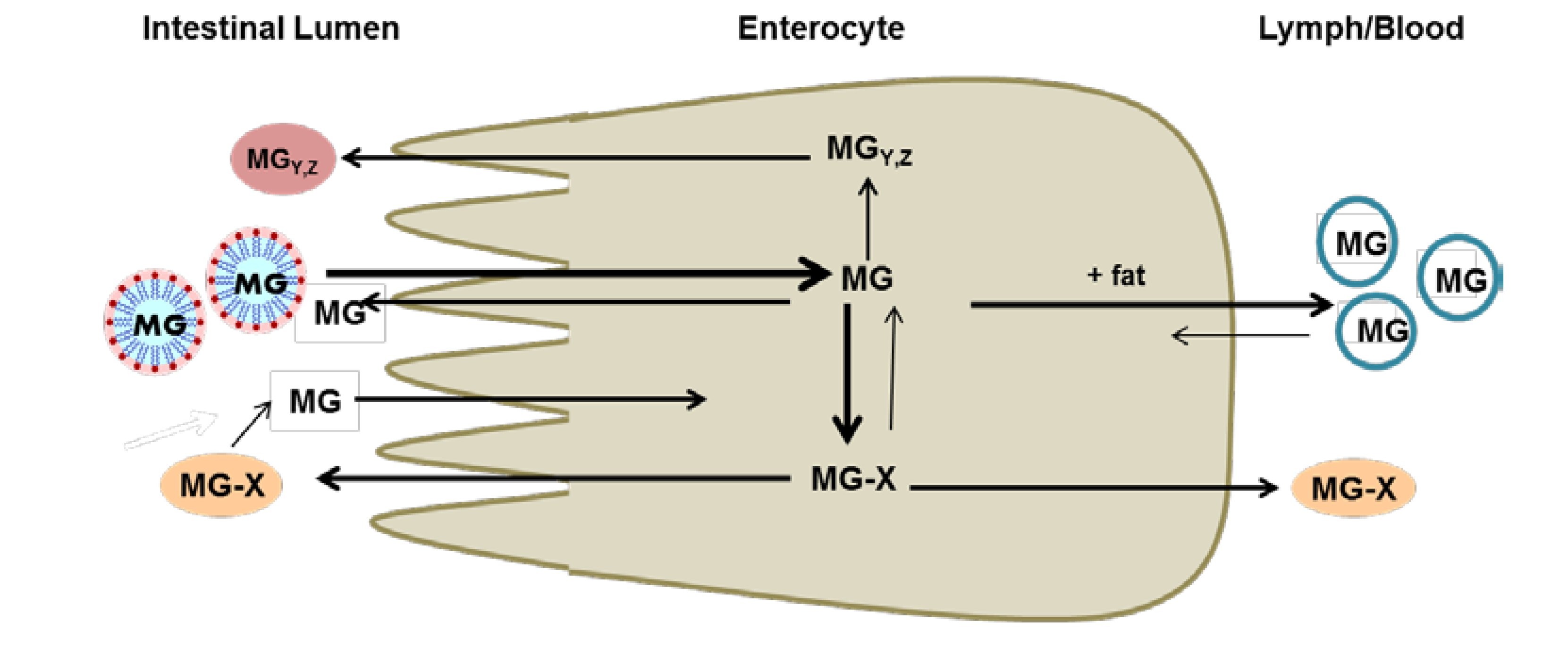
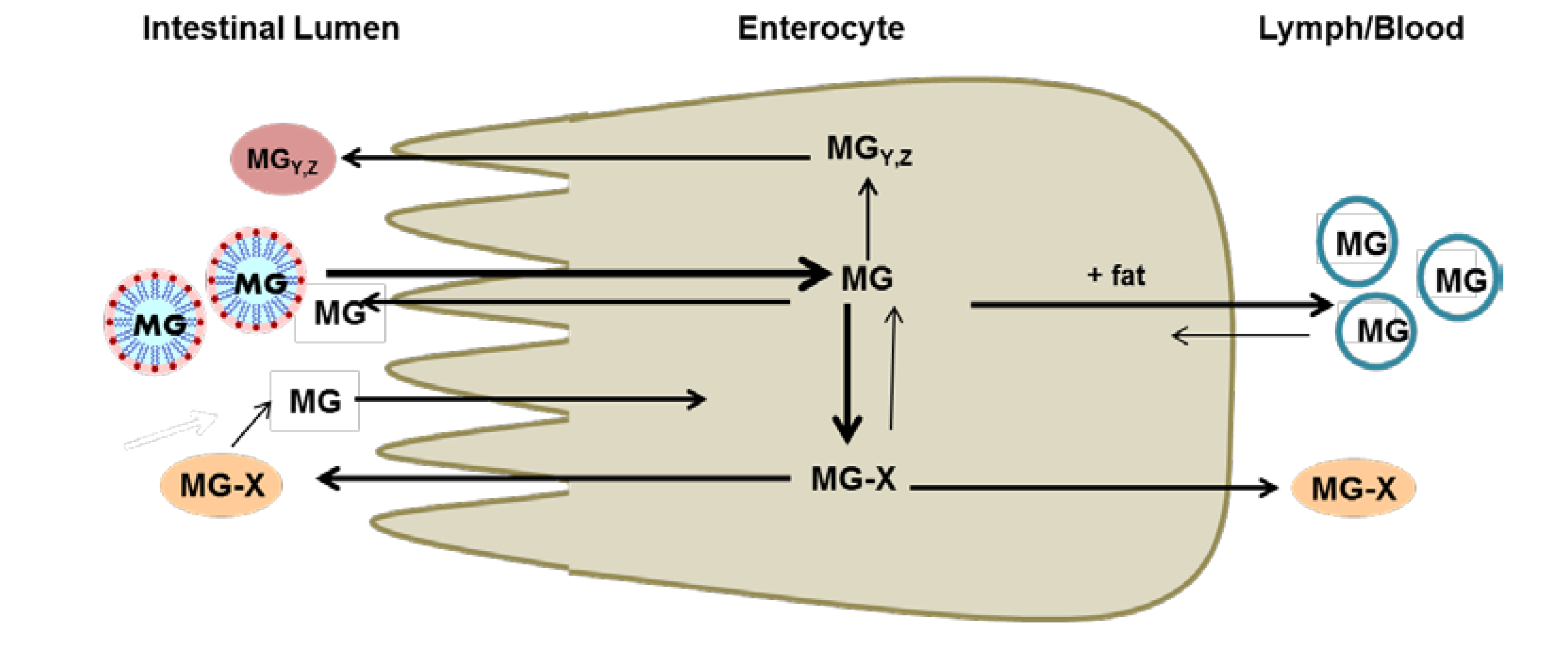
2. Metabolism and Bioavailability of Mangosteen Xanthones

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical properties | Biological activities | Reference |
|---|---|---|
| natural and synthetic derivatives of xanthone | enzyme modulation, anti-tumor activity, anti-microbial, central nervous system(CNS) depressants, CNS stimulants, neurological disorders, anti-convulsant, analgesic, anti-arrhythmic, anti-hypertensive, anti-inflammatory, anti-allergic and immunomodulatory activities | [9] |
| xanthones isolated from pericarp, whole fruit, trunk, leaves and branches | anti-oxidant, anti-tumor, anti-inflammatory, anti-allergic, anti-bacterial, anti-fungal, anti-viral and anti-malarial activities | [3] |
| structural characterization of mangosteen xanthones in whole fruit, stem, aril, seeds, heartwood, leaves | anti-oxidant, anti-bacterial, anti-fungal, anti-malarial, anti-HIV, cytotoxic, aromatase inhibitory, anti-cancer and anti-inflammatory activities | [1] |
| chemical constituents and methods of isolation from pericarp, whole fruit, stem, aril, seeds, heartwood, leaves | anti-oxidant, anti-fungal, anti-bacterial, cytotoxic, anti-histamine, anti-HIV, CNS-depressant, cardiovascular, anti-inflammatory and anti-ulcerative activities | [5] |
| xanthones from mangosteen extracts | anti-cancer, anti-inflammatory, pro-apoptotic, cell cycle arresting, anti-invasive and anti-metastatic activities | [10] |
3. Anti-Cancer Activities of Xanthones
| Cancer cell type | Animal model | Tested compound | Delivery route | Dose | Outcomes | Reference |
|---|---|---|---|---|---|---|
| BJMC3879 (murine mammary adenocarcinoma) | Balb/c | panaxanthone (75%–85% α-MG, 5%–15% γ-MG) | diet | 5000 ppm | suppression of tumor volume and lung metastasis; decreased microvessel density | [18] |
| BJMC3879 (murine mammary adenocarcinoma) | Balb/c | α-MG | subcutaneous | 20 mg/kg/day | decreased tumor growth and metastatic expansion; increased apoptosis; activation of caspase-3; decreased microvessel density; cytochrome c release from mitochondria; cell cycle arrest | [17] |
| GBM8401 (human malignant glioblastoma) | nude Balb/cA-ν (ν/ν) | α-MG | intraperitoneal | 2 mg/kg/day | inhibition of tumor growth by 50%; increased phosphorylation of AMPK; induction of autophagy | [19] |
| 22Rv1 (human prostate carcinoma) | Athymic nu/nu mice | α-MG | oral gavage | 100 mg/kg 5x/week | decreased tumor growth | [20] |
| COLO205 (human colorectal adenocarcinoma) | Athymic NCr nu/nu mice | mangosteen pericarp extract containing 48 mg α-MG and 6.4 mg γ-MG per gram of extract | intratumorally | 0.024–3.0 mg per tumor | complete suppression of tumor growth at 3 mg extract/tumor; apoptotic cells, nuclear fragmentation and chromatin condensation; activation of caspases-3 and -8 | [22] |
| HCT116 (human colorectal carcinoma) | Athymic NCR nu/nu nude mice | extract of mangosteen pericarp (81% α-MG and 16% γ-MG) | diet | 0.25% and 0.5% extract: food ratio (wt/wt) | inhibition of tumor growth; fewer blood vessels in tumor | [23] |
| NL-17 (murine colon adenocarcinoma) | Balb/c | pericarp methanolic extract (25% α-MG) | intraperitoneal | 100–200 mg/kg | reduced tumor mass by 50%–70% | [24] |
| Her2/CT26 cells (murine colon carcinoma) | Balb/c | α-MG | oral | 20 mg/kg | reduced subcutaneous growth | [25] |
| HT-29 (human colon adenocarcinoma) | Athymic Balb/c | α-MG | diet | 900 mg/kg | 40% reduction in tumor mass; decreased Bcl-2 and β-catenin | [13] |
| Chemically induced cancer model | ||||||
| Chemically-induced (1,2dimethylhydra-zine) colon cancer | F344 rats | α-MG | diet | 0.02% and 0.05% in CE-2 basal diet | inhibition of induction and development of ACF; decreased dysplastic foci and β-catenin accumulated crypts; lower proliferating cell nuclear antigen in colon | [21] |
| Cell type | Tested compound | Dose | Outcomes | Reference |
|---|---|---|---|---|
| BJMC3879 (murine mammary adenocarcinoma) | α-MG | 8 µM | induction of apoptosis; cell cycle arrest; activation of caspase-3 and -9; loss of mitochondrial potential | [18] |
| PC3, and 22Rv1 (human prostate carcinoma) | α-MG | 2.5–15 µM | suppressed cell viability and colony formation; cell cycle arrest; activation of caspase-3 | [20] |
| COLO205 (human colorectal adenocarcinoma) | mangosteen extract: 48 mg α-MG and 6.40 mg γ-MG/g extract | 30 µg/mL | induction of apoptosis; activation of caspase-3 and -8; release of mitochondrial cytochrome c | [22] |
| HCT116 (human colorectal carcinoma) | extract of mangosteen pericarp (81% α-MG and 16% γ-MG) | 10–20 µg/mL | reduced cell viability; increased activities of caspase-3/7 and-9; loss of mitochondrial potential; enhanced activity of MAPK/ERK, Myc/Max and p53 signaling; increased JNK; decreased NF-κB | [23] |
| NL-17 (murine colon adenocarcinoma) | pericarp methanol extract (25% α-MG) | >25 µg/mL | anti-proliferative activity | [24] |
| HT-29 (human colon adenocarcinoma) | α-MG | 6–12 µM | anti-proliferative activity; decreased Bcl2 and β-catenin | [13] |
4. Anti-Inflammatory Activity of Xanthones
5. Modulation of Pro-Apoptotic, Anti-Proliferative and Anti-Metastatic Signaling Pathways by Xanthones
| Cell type | Pro-inflammatory insult | Tested compound | Dose | Outcomes | Reference |
|---|---|---|---|---|---|
| Human U397 macrophage-likecells and primary adipocytes | LPS (100 µg/L) for 3 h | α- and γ-MG | α and γ-MG (2 h-pretreatment) with 10 or 30 µmol/L | α- and γ-MG decreased expression of IL-6, TNF-α, IFN-γ-inducible protein (IP)-10 in macrophage-like cells; decreased phosphorylation of MEK, JNK, ERK and p38; only γ-MG pretreatment attenuated LPS-mediated IκBα degradation; α- and γ-MG pretreatment decreased phosphorylation of c-Jun, Elk-1 and ATF-2; α- and γ-MG attenuated LPS-induced PPAR-γ suppression; γ-MG reduced inflammation and insulin resistance in adipocytes | [26] |
| Human primary adipocytes | LPS,10 µg/L for 3 h | α-MG and γ-MG | α- or γ-MG (24 h pretreatment with 3 µmol/L) | α- and γ-MG attenuated LPS-induced inflammatory gene expression of TNF-α, IL-1β, IL-6, IL-8, MCP-1, and Toll-like receptor-2; α- and γ-MG decreased MAPK activation by suppressing phosphorylation of JNK, p38, and ERK; γ-MG attenuated IκBα degradation and NF-κB activation induced by LPS; xanthones inhibited phosphorylation of c-jun and transcriptional activity of AP-1; γ-MG blocked LPS-induced suppression of PPARγ (peroxisome proliferator-activated receptor γ) and its target genes | [28] |
| Human U397 macrophage-like cells | LPS (0.1 ng/mL) for 4 h | α-MG | 6–12 nM for 30 min | α-MG attenuated LPS-stimulated TNF-α secretion by U937 macrophage-like cells and suppressed expression of genes related to immune responses and inflammatory processes such as cytokine production, Th1 and Th2 differentiation, and IL-1 signaling; α-MG decreased activation of p38, ERK1/2, JNK, STAT1, c-Fos and c-Jun | [27] |
| Human cells: primary monocyte-derived macrophages (MDM); macrophage-like THP-1, hepatic HepG2, enterocyte-like Caco-2, and HT-29 colon adenocarcinoma | LPS (100 ng/mL for MDM and HT-29, 0.1 ng/mL for THP-1); PMA (50 ng/mL for HepG2); IL-1β (5 ng/mL for Caco-2) | α-MG | 4.5–10 µM (pretreatment for various times) | inhibition of IL-8 secretion by Caco-2, HT-29 and THP-1 cells; inhibition of TNF-α by HepG2 cells; stimulation of TNF-α by primary MDM cells | [8] |
| Murine RAW 264.7 macrophage-like | LPS (100 µg/mL) | pericarp ethanol extract, α- and γ-MG | pericarp ethanol extract, (3–100 µg/mL), α- and γ-MG (3–100 µM) | α-MG and γ-MG inhibited NO and PGE2 production with moderate inhibitory effects on secretion of TNF-α and IL-4; expression of iNOS and COX-2 mRNA suppressed by α-MG; γ-MG inhibited transcription of iNOS | [29] |
| Murine RAW 264.7 macrophage-like | LPS (0.5–1 µg/mL) | α- and γ-MG | 3–25 µM | inhibition of NO and PGE2 production by α- and γ-MG; iNOS expression reduced by both compounds; COX-2 expression and iNOS enzymatic activity were not affected | [30] |
| Rat RBL-2H3 basophilic leukemia | bovine serum albumin | α-, β-, and γ-MG | 20 µM | α-MG significantly inhibited histamine release and blocked cytoplasmic Ca2+ elevation; γ-MG significantly reduced reactive oxygen species; suppressed phosphorylation of Syk, phospholipase C γ1 and γ2 by all mangostins; complete suppression of phosphorylation of Erk ½; JNK ½ and p38 MAPK signaling not altered; slight suppression of p-Akt; decreased phosphorylation of ERK and cytosolic phospholipase A2 | [31] |
| Rat C6 glioma cells | A23187 calcium ionophore (10 µM) | γ-MG | 1–30 µM | inhibition of COX-1 and -2 activities and PGE2 release by γ-MG; no effects on MAPK/ERK phosphorylation | [32] |
| Rat C6 glioma cells | LPS (10 µg/mL) | γ-MG | 1–30 µM | inhibition of LPS-induced PGE2 release, COX-2 mRNA and protein expression; no effect on COX-1; inhibition of IκB kinase activity; inhibition of IκB degradation; decreased NF-κB activation | [36] |
| Rat C6 glioma cells | A23187 calcium ionophore (10 µM) and LPS (1 µg/mL) | Garcinone B | 10–20 µM | inhibition of COX-1 and COX-2 activities and PGE2 release; inhibition of IKK activity and NF-κB-dependent transcription | [33] |
| Human Caco-2 enterocyte-like cells | IL-1β (25 µg/mL) | aqueous extract of mangosteen pericarp containing polyphenolic compounds | 50 µmol gallic acid equivalents/L | stimulation of basal PGE2 secretion; no effect on IL-8 secretion or activation of ERK, JNK, and NF-κB | [34] |
| Animal studies | |||||
|---|---|---|---|---|---|
| Model | Tested compound/product | Delivery route | Dose | Outcomes | Reference |
| rats: carrageenan-induced hind paw edema, cotton pellet implantation, granuloma pouch technique | α-MG, 1-isomangostin, mangostin triacetate | intra-peritoneal, oral | 50 mg/kg | reduction in paw edema volume, granuloma weight, and granuloma pouch exudate | [35] |
| rat carrageenan-induced hind paw edema | γ-MG | intra-peritoneal | 10 and 30 mg/kg | concentration dependent inhibition of edema formation | [36] |
| rat carrageenan-induced paw edema | α-MG isolated from Allanblackiamonticola | not specified | 9.4 mg/kg | inhibition of edema | [37] |
| mouse carrageenan-induced paw edema | α- and γ-MG | oral | 20 mg/kg | inhibition of paw edema formation by α-MG, but not by γ-MG | [30] |
| mouse OVA-induced allergic asthma | α- and γ-MG | oral | 10 and 30 mg/kg | both xanthones attenuated inflammatory cell recruitment into the airway; reduced airway hyper-responsiveness; lower levels of Th2 cytokines; attenuated PI3K activity, Akt phosphorylation, and NF-κB activation | [38] |
| Human studies | |||||
| human subjects with periodontal pockets | pericarp extract | topical | not specified | clinical improvement in periodontal inflammation; subgingival microbial composition altered from diseased to healthy state | [39] |
| healthy adults | mangosteen supplement containing mangosteen juice, vitamins, minerals, aloe vera, and green tea | oral | 59 mL/day for 30 days | decreased levels of serum CRP levels; increased ratio of T helper to cytotoxic T cells; elevated serum levels of IL-1α and IL-1β, and complement components C3 and C4 | [40] |
| obese subjects | mangosteen juice blend (mangosteen, apple, pear, grape, blueberry, raspberry, strawberry, cranberry and cherry) | oral | 6, 12, and 18 oz/day for 8 weeks | Decreased CRP levels in subjects consuming 18 oz of blended juice; increased levels of IP-10 in subjects consuming 6 and 18 oz of blended juice; no differences in F2 isoprostane and IL-12p70 levels; increased MIP-1 beta in subjects ingesting 18 oz blended juice | [41] |
| Biological activity | Target/messenger/process | Cell type | Reference |
|---|---|---|---|
| Apoptosis | *↓p-ERK1/2; ↓p-JNK1/2 | chondrosarcoma SW1353 | [42] |
| ↓Ψm | leukemia HL60; prostate cancer PC12; colorectal cancer DLD-1; melanoma SK-MEL-28; colorectal HCT116; malignant glioblastoma GBM 8401 | [23,43,44,45,46,47] | |
| ¥↑Caspase-3 | chondrosarcoma SW1353; colon cancer COLO205; leukemia HL60; prostate cancer PC12; melanoma SK-MEL-28; colorectal HCT116; breast cancer MDA-MB231 | [22,23,42,43,44,45,48] | |
| ↑Caspase-8 | chondrosarcoma SW1353; colon cancer COLO205; breast cancer MDA-MB231 | [22,42,48] | |
| ↓Bcl-2; ↑Bax | chondrosarcoma SW1353 | [42] | |
| ↑cytochrome c release | chondrosarcoma SW1353; colon cancer COLO205; leukemia HL60; prostate cancer PC12; breast cancer MDA-MB231 | [22,42,43,44,48] | |
| ↓Akt | chondrosarcoma SW1353; colorectal cancer DLD-1 | [42,46] | |
| ↑p-JNK1/2 | prostate cancer PC12; colorectal cancer DLD-1 | [44,46] | |
| ↑Endonuclease G | colorectal cancer DLD-1 | [46] | |
| ↑p-ERK1/2 | colorectal cancer DLD-1 and HCT116 | [23,46] | |
| ↑microRNA-143 | colorectal cancer DLD-1 | [46] | |
| ↓NF-κB | colorectal HCT116 | [23] | |
| ↑Myc, Max, p53 | colorectal HCT116 | [23] | |
| ↓β-catenin | colorectal HCT116 and SW480 | [49] | |
| Cell cycle | G1 arrest | colorectal cancer DLD-1; melanoma SK-MEL-28; breast cancer MDA-MB231 | [45,48,50] |
| ↓cyclins, cdc2 | colorectal cancer DLD-1; breast cancer MDA-MB231 | [48,50] | |
| Metastasis | ↓MMP-2, MMP-9 | prostate carcinoma PC-3; breast adenocarcinoma MCF-7; lung adenocarcinoma A549 | [51,52,53] |
| ↓u-PA+ | prostate carcinoma PC-3 | [51] | |
| ↓p-JNK1/2 | prostate carcinoma PC-3 | [51] | |
| ↓NF-κB | prostate carcinoma PC-3; breast adenocarcinoma MCF-7; lung adenocarcinoma A549 | [51,52,53] | |
| ↓AP-1 | prostate carcinoma PC-3; breast adenocarcinoma MCF-7 | [51,52,53] | |
| ↓p-ERK1/2 | breast adenocarcinoma MCF-7; lung adenocarcinoma A549 | [51,52,53] | |
| ↓ανβ3 integrin/FAK | lung adenocarcinoma A549 | [51,52,53] |
6. Future Research
7. Conclusions
Acknowledgements
Conflict of Interest
References
- Chin, Y.; Kinghorn, A.D. Structural characterization, biological effects, and synthetic studies on xanthones from mangosteen (Garcinia mangostana), a popular botanical dietary supplement. Mini Rev. Org. Chem. 2008, 5, 355–364. [Google Scholar] [CrossRef]
- Yapwattanaphun, C.; Subhadrabandhu, S.; Sugiura, A.; Yonemori, K.; Utsunomiya, N. Utilization of some Garcinia species in Thailand. Acta Hort. 2002, 575, 563–570. [Google Scholar]
- Pedraza-Chaverri, J.; Cárdenas-Rodríguez, N.; Orozco-Ibarra, M.; Pérez-Rojas, J.M. Medicinal properties of mangosteen (Garcinia mangostana). Food Chem. Toxicol. 2008, 46, 3227–3239. [Google Scholar] [CrossRef]
- Sloan, E.W. Getting ahead of the curve: Phytochemicals. Nutraceutical World 2010, 13, 16–17. [Google Scholar]
- Obolskiy, D.; Pischel, I.; Siriwatanametanon, N.; Heinrich, M. Garcinia mangostana L.: A phytochemical and pharmacological review. Phytother. Res. 2009, 23, 1047–1065. [Google Scholar] [CrossRef]
- Walker, E.B. HPLC analysis of selected xanthones in mangosteen fruit. J. Sep. Sci. 2007, 30, 1229–1234. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Kalpravidh, R.W.; Suksamrarn, S.; Chaivisuthangkura, A.; Chitchumroonchokchai, C.; Failla, M.L. Bioaccessibility, biotransformation, and transport of α-mangostin from Garcinia mangostana (mangosteen) using simulated digestion and Caco-2 human intestinal cells. Mol. Nutr. Food Res. 2009, 53 (Suppl. 1), 54–61. [Google Scholar]
- Gutierrez-Orozco, F.; Chitchumroonchokchai, C.; Lesinski, G.; Suksamrarn, S.; Failla, M. α-Mangostin: Anti-inflammatory activity and metabolism by human cells. J. Agric. Food Chem. 2013, 61, 3891–3900. [Google Scholar] [CrossRef]
- Pinto, M.; Sousa, M.; Nascimento, M.S. Xanthone derivatives: New insights in biological activities. Curr. Med. Chem. 2005, 12, 2517–2538. [Google Scholar] [CrossRef]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef]
- Li, L.; Brunner, I.; Han, A.R.; Hamburger, M.; Kinghorn, A.D.; Frye, R.; Butterweck, V. Pharmacokinetics of α-mangostin in rats after intravenous and oral application. Mol. Nutr. Food Res. 2011, 55 (Suppl. 1), 67–74. [Google Scholar] [CrossRef]
- Syamsudin, L.; Faizatun, L.; Rahayu, L. HPLC analysis and pharmacokinetic study of mangostin after orally administration in rats. T. Pharm. Res. 2009, 2, 43–49. [Google Scholar]
- Chitchumroonchokchai, C.; Thomas-Ahner, J.M.; Li, J.; Riedl, K.M.; Nontakham, J.; Suksumrarn, S.; Clinton, S.K.; Kinghorn, A.D.; Failla, M.L. Anti-tumorigenicity of dietary α-mangostin in an HT-29 colon cell xenograft model and the tissue distribution of xanthones and their phase II metabolites. Mol. Nutr. Food Res. 2013, 57, 203–211. [Google Scholar] [CrossRef]
- Ramaiya, A.; Li, G.; Petiwala, S.M.; Johnson, J.J. Single dose oral pharmacokinetic profile of α-mangostin in mice. Curr. Drug Targets 2012, 13, 1698–1704. [Google Scholar] [CrossRef]
- Kondo, M.; Zhang, L.; Ji, H.; Kou, Y.; Ou, B. Bioavailability and antioxidant effects of a xanthone-rich mangosteen (Garcinia mangostana) product in humans. J. Agric. Food Chem. 2009, 57, 8788–8792. [Google Scholar] [CrossRef]
- Chitchumroonchokchai, C.; Riedl, K.M.; Suksumrarn, S.; Clinton, S.K.; Kinghorn, A.D.; Failla, M.L. Xanthones in mangosteen juice are absorbed and partially conjugated by healthy adults. J. Nutr. 2012, 142, 675–680. [Google Scholar] [CrossRef]
- Shibata, M.; Iinuma, M.; Morimoto, J.; Kurose, H.; Akamatsu, K.; Okuno, Y.; Akao, Y.; Otsuki, Y. α-Mangostin extracted from the pericarp of the mangosteen (Garcinia mangostana Linn) reduces tumor growth and lymph node metastasis in an immunocompetent xenograft model of metastatic mammary cancer carrying a p53 mutation. BMC Med. 2011, 9, 69. [Google Scholar] [CrossRef]
- Doi, H.; Shibata, M.; Shibata, E.; Morimoto, J.; Akao, Y.; Iinuma, M.; Tanigawa, N.; Otsuki, Y. Panaxanthone isolated from pericarp of Garcinia mangostana L. suppresses tumor growth and metastasis of a mouse model of mammary cancer. Anticancer Res. 2009, 29, 2485–2495. [Google Scholar]
- Chao, A.C.; Hsu, Y.L.; Liu, C.K.; Kuo, P.L. α-Mangostin, a dietary xanthone, induces autophagic cell death by activating the AMP-activated protein kinase pathway in glioblastoma cells. J. Agric. Food Chem. 2011, 59, 2086–2096. [Google Scholar] [CrossRef]
- Johnson, J.; Petiwala, S.; Syed, D.; Rasmussen, J.; Adhami, V.; Siddiqui, I.; Kohl, A.; Mukhtar, H. α-Mangostin, a xanthone from mangosteen fruit, promotes cell cycle arrest in prostate cancer and decreases xenograft tumor growth. Carcinogenesis 2012, 33, 413–419. [Google Scholar] [CrossRef]
- Nabandith, V.; Suzui, M.; Morioka, T.; Kaneshiro, T.; Kinjo, T.; Matsumoto, K.; Akao, Y.; Iinuma, M.; Yoshimi, N. Inhibitory effects of crude α-mangostin, a xanthone derivative, on two different categories of colon preneoplastic lesions induced by 1,2-dimethylhydrazine in the rat. Asian Pac. J. Cancer Prev. 2004, 5, 433–438. [Google Scholar]
- Watanapokasin, R.; Jarinthanan, F.; Jerusalmi, A.; Suksamrarn, S.; Nakamura, Y.; Sukseree, S.; Uthaisang-Tanethpongtamb, W.; Ratananukul, P.; Sano, T. Potential of xanthones from tropical fruit mangosteen as anti-cancer agents: Caspase-dependent apoptosis induction in vitro and in mice. Appl. Biochem. Biotechnol. 2010, 162, 1080–1094. [Google Scholar] [CrossRef]
- Aisha, A.; Abu-Salah, K.; Ismail, Z.; Majid, A.M. In vitro and in vivo anti-colon cancer effects of Garcinia mangostana xanthones extract. BMC Complement. Altern. Med. 2012, 12, 104–113. [Google Scholar] [CrossRef]
- Kosem, N.; Ichikawa, K.; Utsumi, H.; Moongkarndi, P. In vivo toxicity and antitumor activity of mangosteen extract. J. Nat. Med. 2013, 67, 255–263. [Google Scholar] [CrossRef]
- Kim, S.J.; Hong, E.H.; Lee, B.R.; Park, M.H.; Kim, J.W.; Pyun, A.R.; Kim, Y.J.; Chang, S.Y.; Chin, Y.W.; Ko, H.J. α-Mangostin reduced ER stress-mediated tumor growth through autophagy activation. Immune Netw. 2012, 12, 253–260. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Kalpravidh, R.W.; Chuang, C.C.; Overman, A.; Martinez, K.; Kennedy, A.; McIntosh, M. Xanthones from mangosteen inhibit inflammation in human macrophages and in human adipocytes exposed to macrophage-conditioned media. J. Nutr. 2010, 140, 842–847. [Google Scholar] [CrossRef]
- Liu, S.H.; Lee, L.T.; Hu, N.Y.; Huange, K.K.; Shih, Y.C.; Munekazu, I.; Li, J.M.; Chou, T.Y.; Wang, W.H.; Chen, T.S. Effects of α-mangostin on the expression of anti-inflammatory genes in U937 cells. Chin. Med. 2012, 7, 19. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Kalpravidh, R.W.; Chitchumroonchokchai, C.; Chuang, C.C.; West, T.; Kennedy, A.; McIntosh, M. Xanthones from mangosteen prevent lipopolysaccharide-mediated inflammation and insulin resistance in primary cultures of human adipocytes. J. Nutr. 2009, 139, 1185–1191. [Google Scholar] [CrossRef]
- Tewtrakul, S.; Wattanapiromsakul, C.; Mahabusarakam, W. Effects of compounds from Garcinia mangostana on inflammatory mediators in RAW264.7 macrophage cells. J. Ethnopharmacol. 2009, 121, 379–382. [Google Scholar]
- Chen, L.G.; Yang, L.L.; Wang, C.C. Anti-inflammatory activity of mangostins from Garcinia mangostana. Food Chem. Toxicol. 2008, 46, 688–693. [Google Scholar] [CrossRef]
- Itoh, T.; Ohguchi, K.; Iinuma, M.; Nozawa, Y.; Akao, Y. Inhibitory effect of xanthones isolated from the pericarp of Garcinia mangostana L. on rat basophilic leukemia RBL-2H3 cell degranulation. Bioorg. Med. Chem. 2008, 16, 4500–4508. [Google Scholar] [CrossRef]
- Nakatani, K.; Nakahata, N.; Arakawa, T.; Yasuda, H.; Ohizumi, Y. Inhibition of cyclooxygenase and prostaglandin E2 synthesis by γ-mangostin, a xanthone derivative in mangosteen, in C6 rat glioma cells. Biochem. Pharmacol. 2002, 63, 73–79. [Google Scholar] [CrossRef]
- Yamakuni, T.; Aoki, K.; Nakatani, K.; Kondo, N.; Oku, H.; Ishiguro, K.; Ohizumi, Y. Garcinone B reduces prostaglandin E2 release and NF-κB-mediated transcription in C6 rat glioma cells. Neurosci. Lett. 2006, 394, 206–210. [Google Scholar] [CrossRef]
- Romier-Crouzet, B.; Walle, V.D.; During, A.; Joly, A.; Rousseau, C.; Henry, O.; Larondelle, Y.; Schneider, Y.J. Inhibition of inflammatory mediators by polyphenolic plant extracts in human intestinal Caco-2 cells. Food Chem. Toxicol. 2009, 47, 1221–1230. [Google Scholar] [CrossRef]
- Shankaranarayan, D.; Gopalakrishnan, C.; Kameswaran, L. Pharmacological profile of mangostin and its derivatives. Arch. Int. Pharmacodyn. Ther. 1979, 239, 257–269. [Google Scholar]
- Nakatani, K.; Yamakuni, T.; Kondo, N.; Arakawa, T.; Oosawa, K.; Shimura, S.; Inoue, H.; Ohizumi, Y. Gamma-mangostin inhibits inhibitor-κB kinase activity and decreases lipopolysaccharide-induced cyclooxygenase-2 gene expression in C6 rat glioma cells. Mol. Pharmacol. 2004, 66, 667–674. [Google Scholar] [CrossRef]
- Nguemfo, E.L.; Dimo, T.; Dongmo, A.B.; Azebaze, A.G.; Alaoui, K.; Asongalem, A.E.; Cherrah, Y.; Kamtchouing, P. Anti-oxidative and anti-inflammatory activities of some isolated constituents from the stem bark of Allanblackia monticola Staner L.C. (Guttiferae). Inflammopharmacology 2009, 17, 37–41. [Google Scholar] [CrossRef]
- Jang, H.Y.; Kwon, O.K.; Oh, S.R.; Lee, H.K.; Ahn, K.S.; Chin, Y.W. Mangosteen xanthones mitigate ovalbumin-induced airway inflammation in a mouse model of asthma. Food Chem. Toxicol. 2012, 50, 4042–4050. [Google Scholar] [CrossRef]
- Rassameemasmaung, S.; Sirikulsathean, A.; Amornchat, C.; Maungmingsook, P.; Rojanapanthu, P.; Gritsanaphan, W. Topical application of Garcinia mangostana L. pericarp gel as an adjunct to periodontal treatment. Complement. Ther. Med. 2008, 16, 262–267. [Google Scholar]
- Tang, Y.P.; Li, P.G.; Kondo, M.; Ji, H.P.; Kou, Y.; Ou, B. Effect of a mangosteen dietary supplement on human immune function: A randomized, double-blind, placebo-controlled trial. J. Med. Food 2009, 12, 755–763. [Google Scholar] [CrossRef]
- Udani, J.K.; Singh, B.B.; Barrett, M.L.; Singh, V.J. Evaluation of mangosteen juice blend on biomarkers of inflammation in obese subjects: A pilot, dose finding study. Nutr. J. 2009, 8, 48–54. [Google Scholar] [CrossRef]
- Krajarng, A.; Nakamura, Y.; Suksamrarn, S.; Watanapokasin, R. α-Mangostin induces apoptosis in human chondrosarcoma cells through downregulation of ERK/JNK and Akt signaling pathway. J. Agric. Food Chem. 2011, 59, 5746–5754. [Google Scholar] [CrossRef]
- Matsumoto, K.; Akao, Y.; Yi, H.; Ohguchi, K.; Ito, T.; Tanaka, T.; Kobayashi, E.; Iinuma, M.; Nozawa, Y. Preferential target is mitochondria in α-mangostin-induced apoptosis in human leukemia HL60 cells. Bioorg. Med. Chem. 2004, 12, 5799–5806. [Google Scholar] [CrossRef]
- Sato, A.; Fujiwara, H.; Oku, H.; Ishiguro, K.; Ohizumi, Y. α-Mangostin induces Ca2+-ATPase-dependent apoptosis via mitochondrial pathway in PC12 cells. J. Pharmacol. Sci. 2004, 95, 33–40. [Google Scholar] [CrossRef]
- Wang, J.; Sanderson, B.; Zhang, W. Cytotoxic effect of xanthones from pericarp of the tropical fruit mangosteen (Garcinia mangostana Linn.) on human melanoma cells. Food Chem. Toxicol. 2011, 49, 2385–2391. [Google Scholar]
- Nakagawa, Y.; Iinuma, M.; Naoe, T.; Nozawa, Y.; Akao, Y. Characterized mechanism of α-mangostin-induced cell death: Caspase-independent apoptosis with release of endonuclease-G from mitochondria and increased miR-143 expression in human colorectal cancer DLD-1 cells. Bioorg. Med. Chem. 2007, 15, 5620–5628. [Google Scholar] [CrossRef]
- Chang, H.; Huang, W.; Chen, H.; Yang, L.L. Apoptotic effects of γ-mangostin from the fruit hull of Garcinia mangostana on human malignant glioma cells. Molecules 2010, 15, 8953–8966. [Google Scholar] [CrossRef]
- Kurose, H.; Shibata, M.; Iinuma, M.; Otsuki, Y. Alterations in cell cycle and induction of apoptotic cell death in breast cancer cells treated with α-mangostin extracted from mangosteen pericarp. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Yoo, J.; Kang, K.; Jho, E.H.; Chin, Y.; Kim, J.; Nho, C.W. α- and γ-Mangostin inhibit the proliferation of colon cancer cells via β-catenin gene regulation in Wnt/cGMP signaling. Food Chem. 2011, 129, 1559–1566. [Google Scholar] [CrossRef]
- Matsumoto, K.; Akao, Y.; Ohguchi, K.; Ito, T.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar] [CrossRef]
- Hung, S.; Shen, K.; Wu, C.; Liu, C.; Shih, Y. α-Mangostin suppresses PC-3 human prostate carcinoma cell metastasis by inhibiting matrix metalloproteinase-2/9 and urokinase-plasminogen expression through the JNK signaling pathway. J. Agric. Food Chem. 2009, 57, 1291–1298. [Google Scholar] [CrossRef]
- Lee, Y.; Ko, K.; Shi, M.; Liao, Y.; Chiang, T.; Wu, P.; Shih, Y.; Shih, Y. α-Mangostin, a novel dietary xanthone, suppresses TPA-mediated MMP-2 and MMP-9 expressions through the ERK signaling pathway in MCF-7 human breast adenocarcinoma cells. J. Food Sci. 2010, 75, H13–H23. [Google Scholar] [CrossRef]
- Shih, Y.; Chien, S.; Chen, P.; Lee, J.; Wu, S.; Yin, L. α-Mangostin suppresses phorbol 12-myristate 13-acetate-induced MMP-2/MMP-9 expressions via αvβ3 integrin/FAK/ERK and NF-κB signaling pathway in human lung adenocarcinoma A549 cells. Cell Biochem. Biophys. 2010, 58, 31–44. [Google Scholar] [CrossRef]
- Williamson, G.; Barron, D.; Shimoi, K.; Terao, J. In vitro biological properties of flavonoid conjugates found in vivo. Free Radic. Res. 2005, 39, 457–469. [Google Scholar] [CrossRef]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated quercetin glucuronides as bioactive metabolites and precursors of aglycone in vivo. Food Funct. 2011, 2, 11–17. [Google Scholar] [CrossRef]
- Li, S.; Sang, S.; Pan, M.; Lai, C.; Lo, C.; Yang, C.; Ho, C. Anti-inflammatory property of the urinary metabolites of nobiletin in mouse. Bioorg. Med. Chem. Lett. 2007, 17, 5177–5181. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; Yáñez-Gascón, M.; Selma, M.; Azorín-Ortuño, M.; Toti, S.; Tomás-Barberán, F.; Dolara, P.; Espín, J. Anti-Inflammatory properties of a pomegranate extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on phenolic metabolism. J. Nutr. Biochem. 2010, 21, 717–725. [Google Scholar] [CrossRef]
- Halliwell, B. The wanderings of a free radical. Free Radic. Biol. Med. 2009, 46, 531–542. [Google Scholar] [CrossRef]
- Long, L.; Hoi, A.; Halliwell, B. Instability of, and generation of hydrogen peroxide by, phenolic compounds in cell culture media. Arch. Biochem. Biophys. 2010, 501, 162–169. [Google Scholar] [CrossRef]
- Speciale, A.; Chirafisi, J.; Saija, A.; Cimino, F. Nutritional antioxidants and adaptive cell responses: An update. Curr. Mol. Med. 2011, 11, 770–789. [Google Scholar] [CrossRef]
- Siow, R.; Mann, G.E. Dietary isoflavones and vascular protection: Activation of cellular antioxidant defenses by SERMs or hormesis? Mol. Aspects Med. 2010, 31, 468–477. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gutierrez-Orozco, F.; Failla, M.L. Biological Activities and Bioavailability of Mangosteen Xanthones: A Critical Review of the Current Evidence. Nutrients 2013, 5, 3163-3183. https://doi.org/10.3390/nu5083163
Gutierrez-Orozco F, Failla ML. Biological Activities and Bioavailability of Mangosteen Xanthones: A Critical Review of the Current Evidence. Nutrients. 2013; 5(8):3163-3183. https://doi.org/10.3390/nu5083163
Chicago/Turabian StyleGutierrez-Orozco, Fabiola, and Mark L. Failla. 2013. "Biological Activities and Bioavailability of Mangosteen Xanthones: A Critical Review of the Current Evidence" Nutrients 5, no. 8: 3163-3183. https://doi.org/10.3390/nu5083163
APA StyleGutierrez-Orozco, F., & Failla, M. L. (2013). Biological Activities and Bioavailability of Mangosteen Xanthones: A Critical Review of the Current Evidence. Nutrients, 5(8), 3163-3183. https://doi.org/10.3390/nu5083163