Pathology of Bursae of Fabricius in Methionine-Deficient Broiler Chickens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chickens and Diets
2.2. Clinical Signs and Relative Bursa Weights
2.3. Pathological Observation
2.4. Cell Cycle of Bursa of Fabricius
2.5. Annexin-V Apoptosis Detection by Flow Cytometry
2.6. Statistical Analysis
3. Results
3.1. Clinical Observation

{kind=link}
{kind=link}
| Groups | 7 days | 14 days | 21 days | 28 days | 35 days | 42 days |
|---|---|---|---|---|---|---|
| Control group | 148.40 ± 3.05 | 354.27 ± 9.24 | 657.73 ± 20.11 | 999.80 ± 28.16 | 1347.40 ± 54.42 | 1781.33 ± 78.18 |
| Met Deficiency | 138.40 ± 2.38 | 313.73 ± 5.52 ** | 543.60 ± 10.69 ** | 808.20 ± 16.60 ** | 1182.00 ± 28.93 * | 1562.93 ± 37.13 * |
3.2. Changes of Weight and Relative Weight of Bursa
| Item | Groups | 7 days | 14 days | 21 days | 28 days | 35 days | 42 days |
|---|---|---|---|---|---|---|---|
| Weight of bursae (g) | Control group | 0.19 ± 0.02 | 0.58 ± 0.05 | 1.41 ± 0.16 | 1.88 ± 0.14 | 2.84 ± 0.42 | 3.90 ± 0.46 |
| Met deficiency | 019 ± 0.02 | 0.53 ± 0.05 | 1.05 ± 0.11 * | 1.12 ± 0.24 ** | 1.29 ± 0.28 ** | 2.17 ± 0.43 ** | |
| Relative weight of bursae (g/kg) | Control group | 1.57 ± 0.01 | 1.69 ± 0.01 | 2.08 ± 0.01 | 1.79 ± 0.01 | 1.87 ± 0.01 | 1.93 ± 0.01 |
| Met deficiency | 1.50 ± 0.02 | 1.56 ± 0.01 | 1.69 ± 0.01 ** | 1.40 ± 0.01 ** | 1.53 ± 0.01 ** | 1.38 ± 0.01 ** |
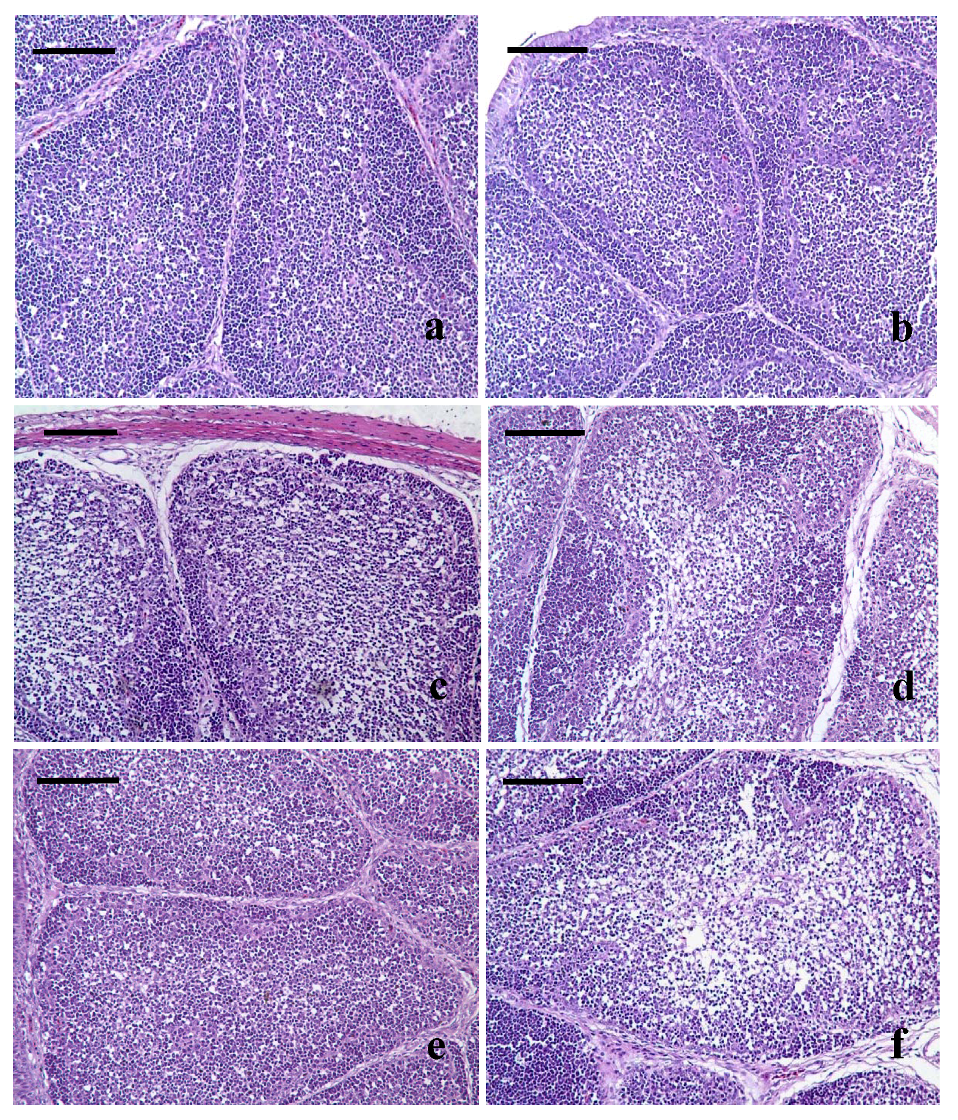
3.3. Pathological Lesions of Bursae of Fabricius
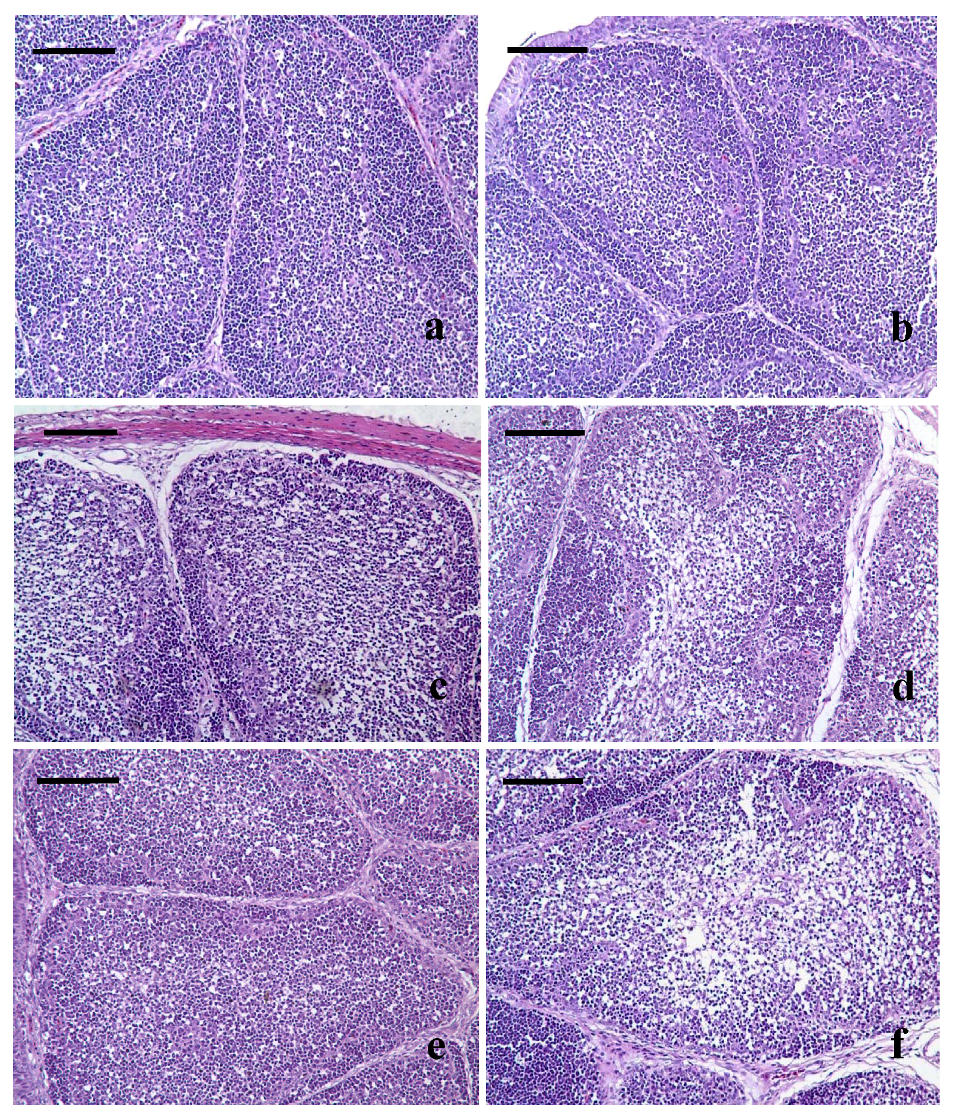
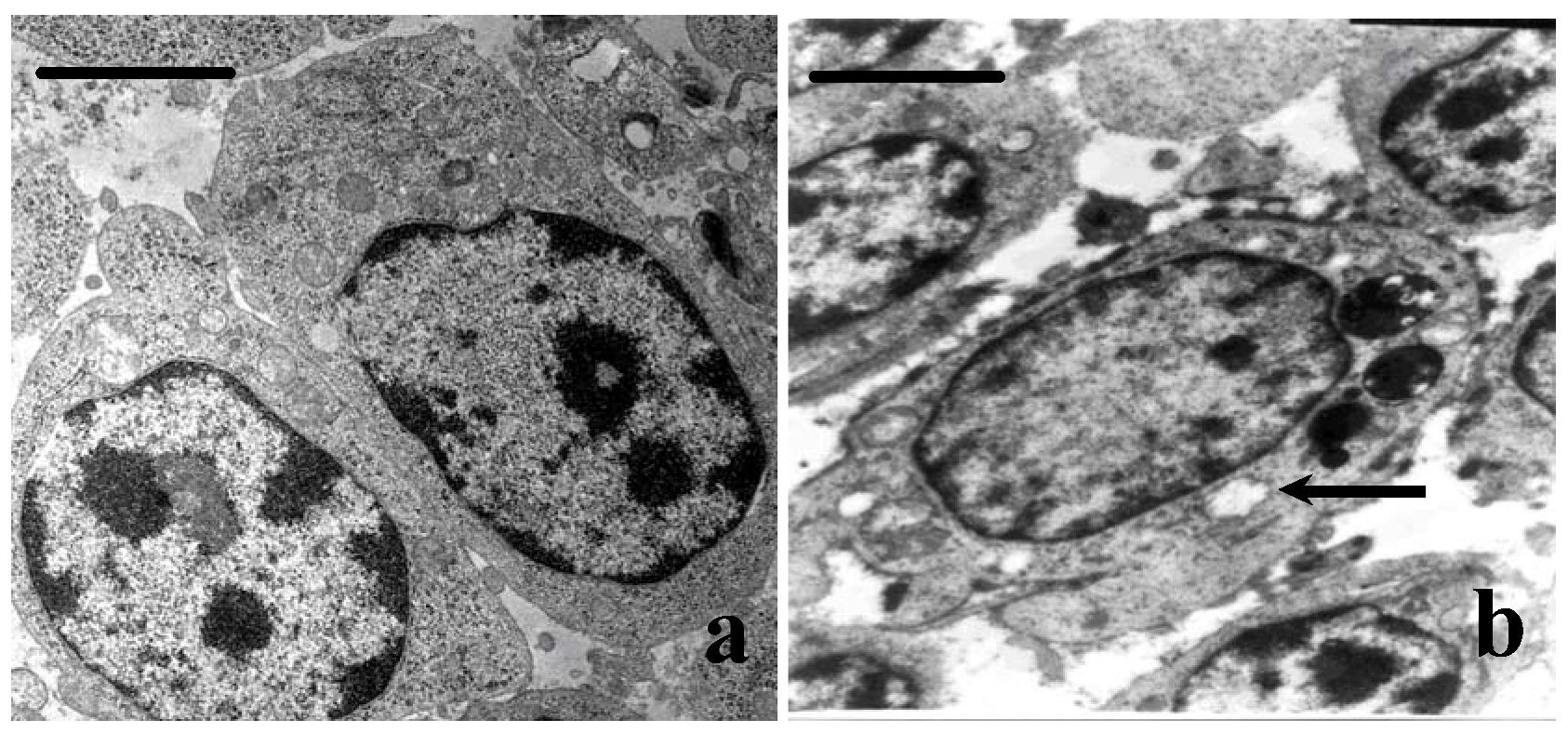
3.4. Ultrastructural Changes
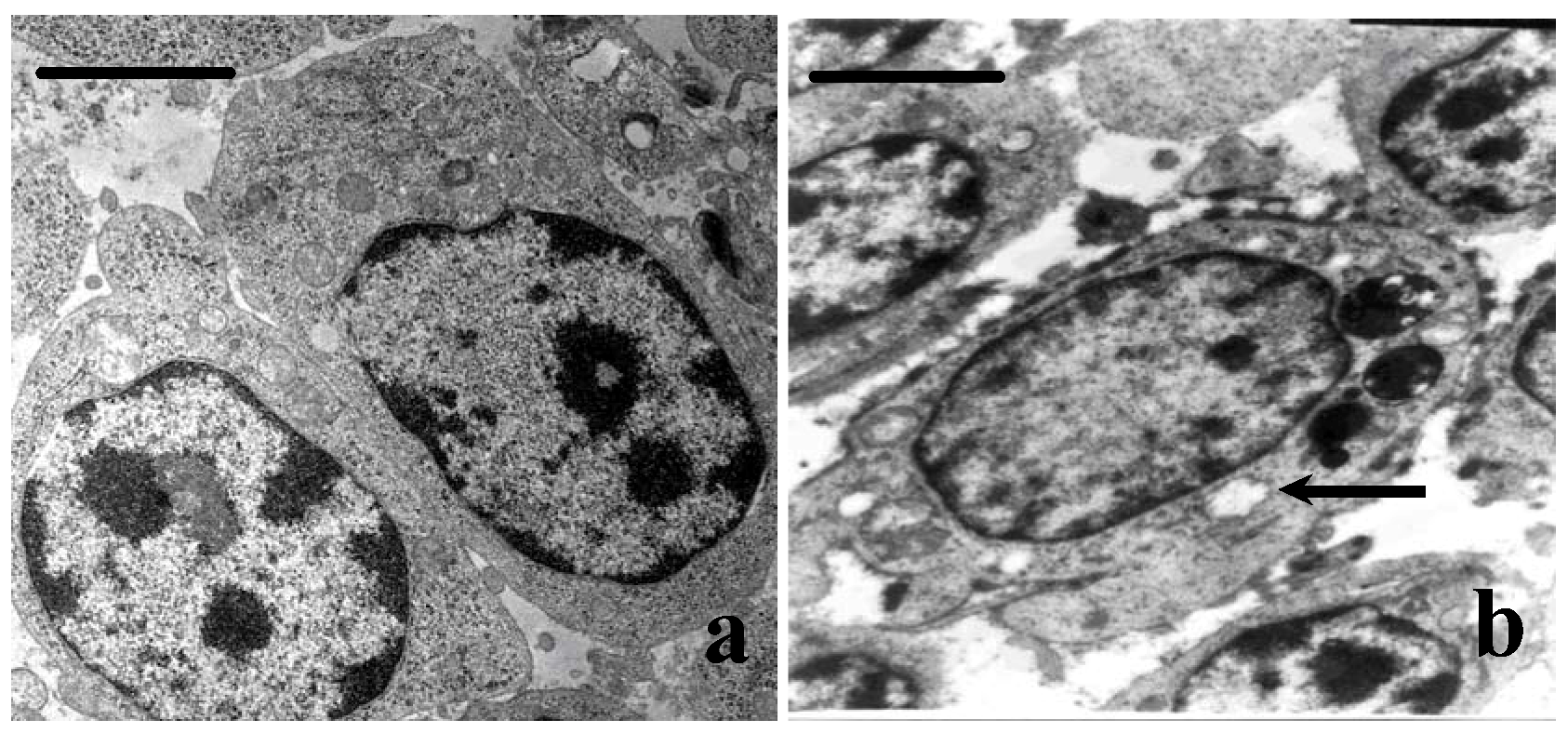
3.5. Cell Cycle Phase Distribution of Bursa Cells
| Groups | Item | 14 days | 28 days | 42 days |
|---|---|---|---|---|
| Control group | G0G1 | 82.93 ± 0.36 | 83.84 ± 0.57 | 87.95 ± 0.31 |
| S | 8.55 ± 0.05 | 7.97 ± 0.57 | 5.83 ± 0.23 | |
| G2M | 8.65 ± 0.24 | 8.18 ± 0.67 | 6.22 ± 0.08 | |
| PI a | 17.14 ± 0.39 | 16.16 ± 0.56 | 11.93 ± 0.35 | |
| Met deficiency | G0G1 | 85.91 ± 0.42 ** | 88.24 ± 0.21 ** | 93.66 ± 0.40 ** |
| S | 6.96 ± 0.31 ** | 5.44 ± 0.11 ** | 2.84 ± 0.06 ** | |
| G2M | 7.33 ± 0.15 ** | 6.32 ± 0.21 ** | 3.38 ± 0.44 ** | |
| PI a | 14.09 ± 0.43 ** | 11.76 ± 0.21 ** | 6.22 ± 0.40 ** |
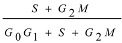 × 100%; Data are the means ± standard deviation (n = 5); * P < 0.05, compared with the control group, ** P < 0.01, compared with the control group; Data were analyzed by Independent-Samples T test using SPSS 11.0 software.
× 100%; Data are the means ± standard deviation (n = 5); * P < 0.05, compared with the control group, ** P < 0.01, compared with the control group; Data were analyzed by Independent-Samples T test using SPSS 11.0 software.3.6. Annexin-V-FITC Staining Assay by Flow Cytometry
| Group | 14 days | 28 days | 42 days |
|---|---|---|---|
| Control group | 1.99 ± 0.03 | 2.48 ± 0.29 | 3.19 ± 0.34 |
| Met deficiency | 2.13 ± 0.10 | 3.20 ± 0.27 * | 5.74 ± 0.54 ** |
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Carew, L.B.; McMurtry, J.P.; Alster, F.A. Effects of methionine deficiencies on plasma levels of thyroid hormones, insulin-like growth factors-I and -II, liver and body weights and feed intake in growing chickens. Poult. Sci. 2003, 82, 1932–1938. [Google Scholar]
- Barnes, D.M.; Calvert, C.C.; Klasing, K.C. Methionine deficiency decreases protein accretion and synthesis but not tRNA acylation in muscles of chicks. J. Nutr. 1995, 125, 2623–2630. [Google Scholar]
- Xie, M.; Hou, S.S.; Huang, W.; Fan, H.P. Effect of excess methionine and methionine hydroxy analogue on growth performance and plasma homocysteine of growing Pekin ducks. Poult. Sci. 2007, 86, 1995–1999. [Google Scholar]
- Sekiz, S.S.; Scott, M.L.; Nesheim, M.C. The effect of methionine deficiency on body weight, food and energy utilization in the chick. Poult. Sci. 1975, 54, 1184–1188. [Google Scholar] [CrossRef]
- Troen, A.M.; Lutgens, E.; Smith, D.E.; Rosenberg, I.H.; Selhub, J. The atherogenic effect of excess methionine intake. Proc. Natl. Acad. Sci. USA 2003, 100, 15089–15094. [Google Scholar]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar]
- Dunlevy, L.P.E.; Chitty, L.S.; Burren, K.A.; Doudney, K.; Mikic, T.S.; Stanier, P.; Scott, R.; Copp, A.J.; Greene, N.D.E. Excess methionine suppresses the methylation cycle and inhibits neural tube closure in mouse embryos. FEBS Lett. 2006, 580, 2803–2807. [Google Scholar] [CrossRef]
- Kano, Y.; Sakamoto, S.; Kasahara, T.; Kusumoto, K.; Hida, K.; Suda, K.; Ozawa, K.; Miura, Y.; Takaku, F. Methionine dependency of cell growth in normal and malignant hematopoietic cells. Cancer Res. 1982, 42, 3090–3092. [Google Scholar]
- Mori, N.; Hirayama, K. Long-term consumption of a methionine-supplemented diet increases iron and lipid peroxide levels in rat liver. J. Nutr. 2000, 130, 2349–2355. [Google Scholar]
- Feo, F.; Pascale, R.; Garcea, R.; Daino, L.; Pirisi, L.; Frassetto, S.; Ruggiu, M.E.; di Padova, C.; Stramentinoli, G. Effect of the variations of s-adenosyl-l-methionine liver content on fat accumulation and ethanol metabolism in ethanol-intoxicated rats. Toxicol. Appl. Pharmacol. 1986, 83, 331–341. [Google Scholar] [CrossRef]
- Swain, B.K.; Johri, T.S. Effect of supplemental methionine, choline and their combinations on the performance and immune response of broilers. Br. Poult. Sci. 2000, 41, 83–88. [Google Scholar] [CrossRef]
- Tsiagbe, V.K.; Cook, M.E.; Harper, A.E.; Sunde, M.L. Enhanced immune responses in broiler chicks fed methionine-supplemented diets. Poult. Sci. 1987, 66, 1147–1154. [Google Scholar] [CrossRef]
- Tsiagbe, V.K.; Cook, M.E.; Harper, A.E.; Sunde, M.L. Efficacy of cysteine in replacing methionine in the immune responses of broiler chicks. Poult. Sci. 1987, 66, 1138–1146. [Google Scholar] [CrossRef]
- Klasing, K.C.; Barnes, D.M. Decreased amino acid requirements of growing chicks due to immunologic stress. J. Nutr. 1988, 118, 1158–1164. [Google Scholar]
- Konashi, S.; Takahashi, K.; Akiba, Y. Effects of dietary essential amino acid deficiencies on immunological variables in broiler chickens. Br. J. Nutr. 2000, 83, 449–456. [Google Scholar]
- Zhang, L.B.; Guo, Y.M. Effects of liquid DL-2-hydroxy-4-methylthio butanoic acid on growth performance and immune responses in broiler chickens. Poult. Sci. 2008, 87, 1370–1376. [Google Scholar] [CrossRef]
- Yen, C.L.E.; Mar, M.H.; Craciunescu, C.N.; Edwards, L.J.; Steven, H.Z. Deficiency in methionine, tryptophan, isoleucine, or choline induces apoptosis in cultured cells. J. Nutr. 2002, 132, 1840–1847. [Google Scholar]
- Thompson, J.N.; Scott, M.L. Impaired lipid and vitamin E absorption related to atrophy of the pancreas in selenium-deficient chicks. J. Nutr. 1970, 100, 797–809. [Google Scholar]
- Cui, H.M. The histopathologic study of selenium deficiency in chicks. Acta Vet. Zootech. Sin. 1988, 19, 52–56. [Google Scholar]
- National Research Council, Nutrient Requirements of Domestic Animals. In Nutrient Requirements of Poultry, 9th ed; National Academy of Science: Washington, DC, USA, 1994.
- Peng, X.; Cui, Y.; Cui, W.; Deng, J.L.; Cui, H.M. The decrease of relative weight, lesions, and apoptosis of Bursa of Fabricius induced by excess dietary selenium in chickens. Biol. Trace Elem. Res. 2009, 131, 33–42. [Google Scholar] [CrossRef]
- Cui, H.M.; Fang, J.; Peng, X. Pathology of the thymus, spleen and bursa of Fabricius in zinc-deficient ducklings. Avian Pathol. 2003, 32, 259–263. [Google Scholar] [CrossRef]
- Cooper, M.D.; Peterson, R.D.A.; South, M.A.; Good, R.A. The functions of the thymus system and the bursa system in the chicken. J. Exp. Med. 1966, 123, 75–102. [Google Scholar]
- Toivanen, P.; Toivanen, A.; Good, R.A. Ontogeny of bursal function in chicken I. Embryonic stem cell for humoral immunity. J. Immunol. 1972, 109, 1058–1070. [Google Scholar]
- Pines, J. Cyclins, CDKs and cancer. Semin. Cancer Biol. 1995, 6, 63–72. [Google Scholar] [CrossRef]
- King, K.L.; Cidlowski, J.A. Cell cycle and apoptosis: Common pathways to life and death. J. Cell. Biochem. 2004, 58, 175–180. [Google Scholar]
- Yen, C.L.E.; Mar, M.H.; Steven, H.Z. Choline deficiency-induced apoptosis in PC12 cells is associated with diminished membrane phosphatidylcholine and sphingomyelin, accumulation of ceramide and diacylglycerol, and activation of a caspase. FASEB J. 1999, 13, 135–142. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, B.; Cui, H.; Peng, X.; Fang, J.; Cui, W.; Liu, X. Pathology of Bursae of Fabricius in Methionine-Deficient Broiler Chickens. Nutrients 2013, 5, 877-886. https://doi.org/10.3390/nu5030877
Wu B, Cui H, Peng X, Fang J, Cui W, Liu X. Pathology of Bursae of Fabricius in Methionine-Deficient Broiler Chickens. Nutrients. 2013; 5(3):877-886. https://doi.org/10.3390/nu5030877
Chicago/Turabian StyleWu, Bangyuan, Hengmin Cui, Xi Peng, Jing Fang, Wei Cui, and Xiaodong Liu. 2013. "Pathology of Bursae of Fabricius in Methionine-Deficient Broiler Chickens" Nutrients 5, no. 3: 877-886. https://doi.org/10.3390/nu5030877
APA StyleWu, B., Cui, H., Peng, X., Fang, J., Cui, W., & Liu, X. (2013). Pathology of Bursae of Fabricius in Methionine-Deficient Broiler Chickens. Nutrients, 5(3), 877-886. https://doi.org/10.3390/nu5030877