
Anti-Hyperuricemic and Nephroprotective Effects of Hydrolysate Derived from Silkworm Pupae (Bombyx mori): In Vitro and In Vivo Study

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation and Enzymatic Hydrolysis of Silkworm Pupae Protein
2.3. Molecular Weight and Peptide Sequence Analysis of SPP
2.4. Assessment of XOD Inhibitory Activity
2.5. Animal Experiments
2.6. Biochemical Indexes Analysis
2.7. Detection of Inflammatory Cytokine Levels in Kidney
2.8. Histological Analysis
2.9. Western Blot Analysis
2.10. qRT-PCR Analysis
2.11. Statistical Analysis
3. Results
3.1. Preparation of SPP
3.2. Molecular Weight and Peptide Sequence of SPP
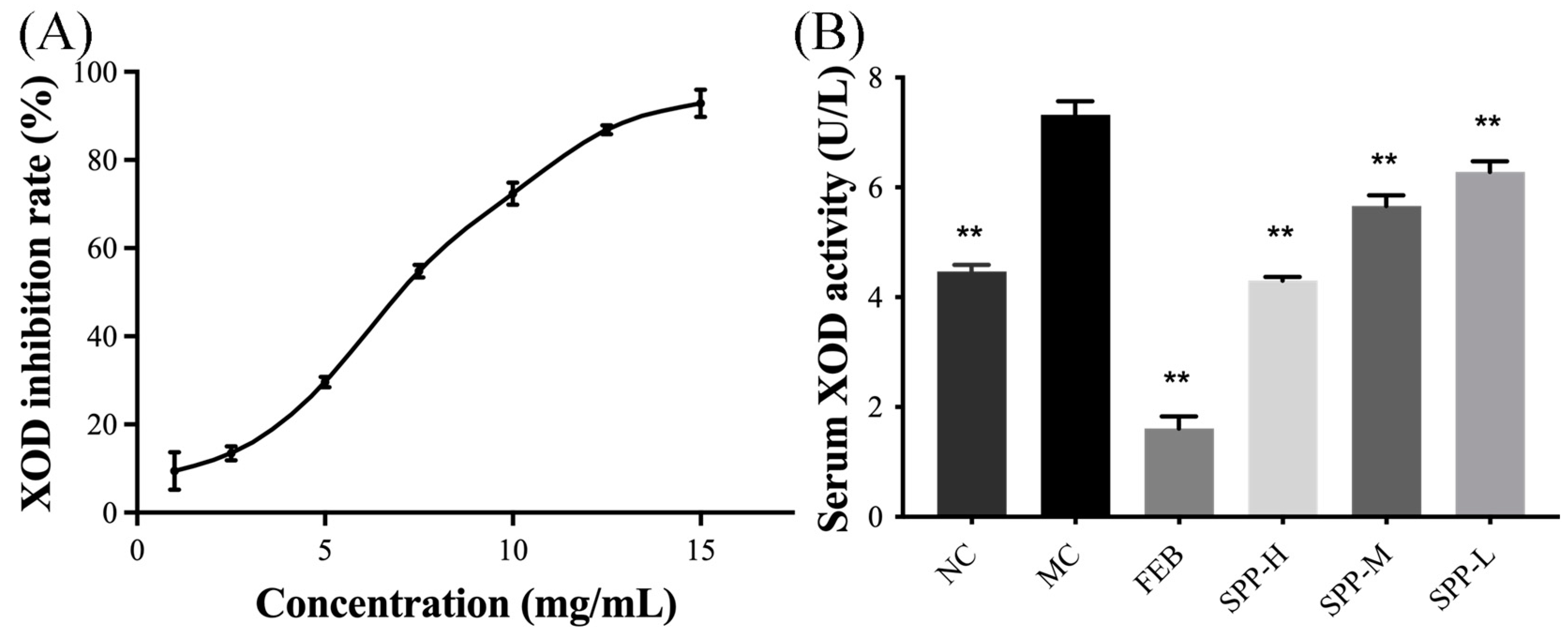
3.3. XOD Inhibitory Activity
3.4. SPP Alleviated Kidney Damage in Hyperuricemic Mice
3.5. Histopathological Examination
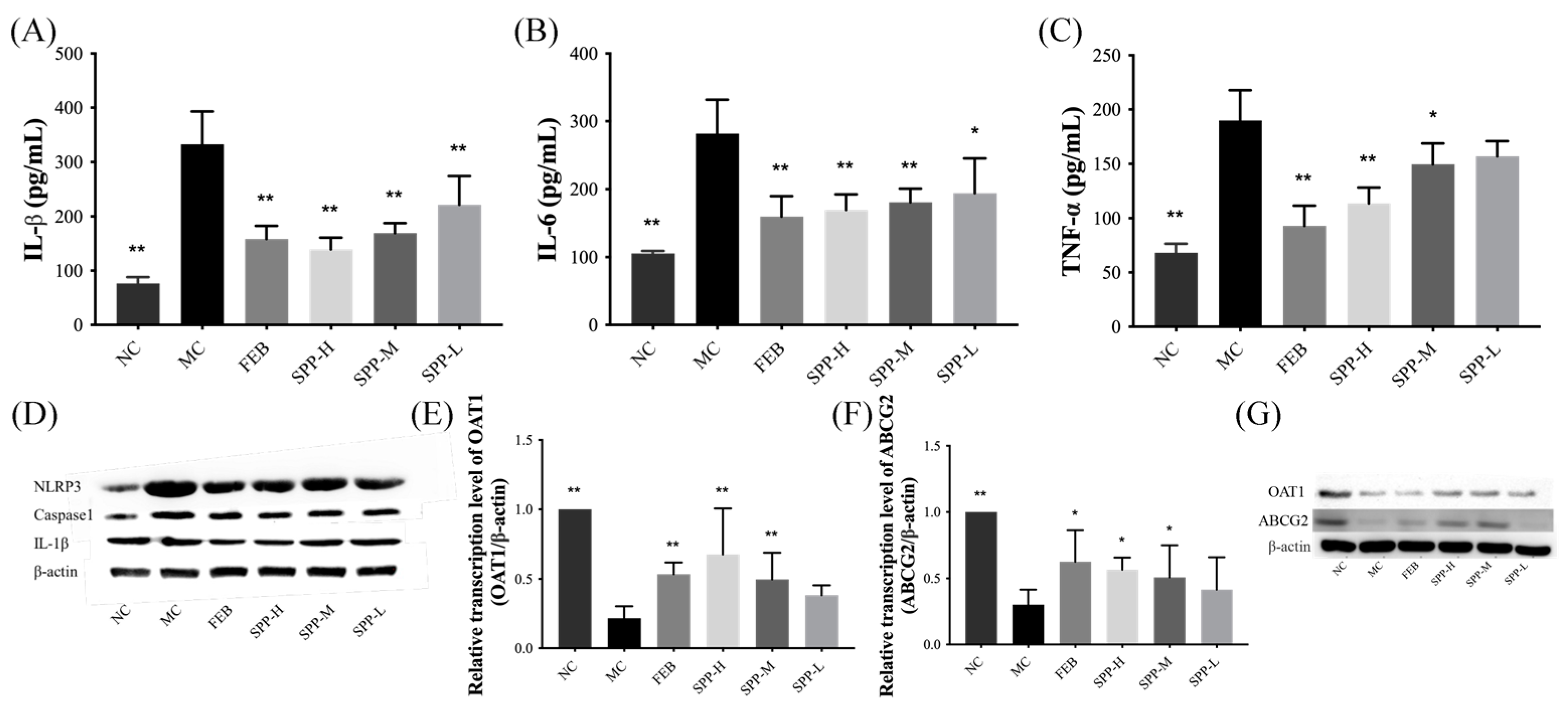
3.6. Nephrotic Inflammation in Mice
3.7. Expression of UA Transporter Proteins
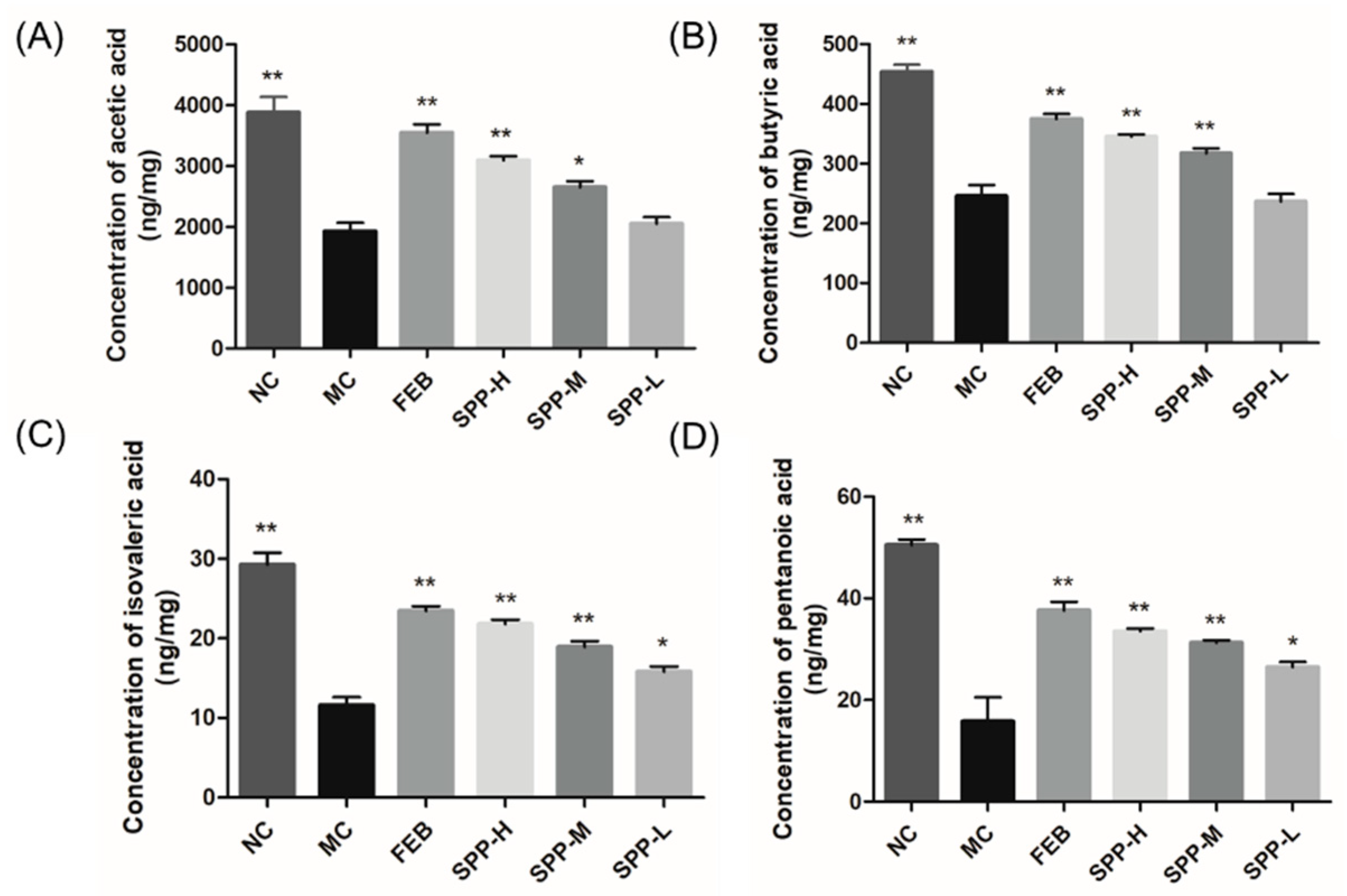
3.8. Short-Chain Fatty Acid Profiles in Hyperuricemic Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Li, C.; Duan, S.; Yuan, X.; Liang, J.; Hou, S. Curcumin Attenuates Potassium Oxonate-Induced Hyperuricemia and Kidney Inflammation in Mice. Biomed. Pharmacother. 2019, 118, 109195. [Google Scholar] [CrossRef]
- Du, Y.; Liu, Z.; Qiao, F.; Wang, S.; Chen, K.; Zhang, X. Computational Exploration of Reactive Fragment for Mechanism-Based Inhibition of Xanthine Oxidase. J. Organomet. Chem. 2018, 864, 58–67. [Google Scholar] [CrossRef]
- Yu, Q.; Sun, Z.; Wang, Y.; Du, X.; Huang, J.; Wang, L. Hyperuricemia Is Accompanied by Elevated Peripheral CD4+ T Cells. Sci. Rep. 2023, 13, 12537. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, X.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Li, Y.; Ming, T.; Wang, Z.J.; Su, X. Protective Effects of Tuna Meat Oligopeptides (TMOP) Supplementation on Hyperuricemia and Associated Renal Inflammation Mediated by Gut Microbiota. FASEB J. 2020, 34, 5061–5076. [Google Scholar] [CrossRef] [PubMed]
- Skoczyńska, M.; Chowaniec, M.; Szymczak, A.; Langner-Hetmańczuk, A.; Maciążek-Chyra, B.; Wiland, P. Pathophysiology of Hyperuricemia and Its Clinical Significance—A Narrative Review. Reumatologia 2020, 58, 312–323. [Google Scholar] [CrossRef]
- Kuwabara, M.; Kodama, T.; Ae, R.; Kanbay, M.; Andres-Hernando, A.; Borghi, C.; Hisatome, I.; Lanaspa, M.A. Update in Uric Acid, Hypertension, and cardiovascular diseases. Hypertens. Res. 2023, 46, 1714–1726. [Google Scholar] [CrossRef]
- Li, Y.J.; Chen, L.R.; Yang, Z.L.; Wang, P.; Jiang, F.F.; Guo, Y.; Qian, K.; Yang, M.; Yin, S.J.; He, G.H. Comparative Efficacy and Safety of Uricosuric Agents in the Treatment of Gout or Hyperuricemia: A Systematic Review and Network Meta-Analysis. Clin. Rheumatol. 2023, 42, 215–224. [Google Scholar] [CrossRef]
- Yan, F.; Xue, X.; Lu, J.; Dalbeth, N.; Qi, H.; Yu, Q.; Wang, C.; Sun, M.; Cui, L.; Liu, Z.; et al. Superiority of Low-Dose Benzbromarone to Low-Dose Febuxostat in a Prospective, Randomized Comparative Effectiveness Trial in Gout Patients with Renal Uric Acid Underexcretion. Arthritis Rheumatol. 2022, 74, 2015–2023. [Google Scholar] [CrossRef]
- Wan, H.; Han, J.; Tang, S.; Bao, W.; Lu, C.; Zhou, J.; Ming, T.; Li, Y.; Su, X. Comparisons of Protective Effects between Two Sea Cucumber Hydrolysates against Diet Induced Hyperuricemia and Renal Inflammation in Mice. Food Funct. 2020, 11, 1074–1086. [Google Scholar] [CrossRef]
- Badve, S.V.; Pascoe, E.M.; Tiku, A.; Boudville, N.; Brown, F.G.; Cass, A.; Clarke, P.; Dalbeth, N.; Day, R.O.; de Zoysa, J.R.; et al. Effects of Allopurinol on the Progression of Chronic Kidney Disease. N. Engl. J. Med. 2020, 382, 2504–2513. [Google Scholar] [CrossRef]
- Lin, L.; Yang, Q.; Zhao, K.; Zhao, M. Identification of the Free Phenolic Profile of Adlay Bran by UPLC-QTOF-MS/MS and Inhibitory Mechanisms of Phenolic Acids against Xanthine Oxidase. Food Chem. 2018, 253, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.A.; Cleveland, J.D. Hypersensitivity Reactions with Allopurinol and Febuxostat: A Study Using the Medicare Claims Data. Ann. Rheum. Dis. 2020, 79, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Chau, Y.T.; Chen, H.Y.; Lin, P.H.; Hsia, S.M. Preventive Effects of Fucoidan and Fucoxanthin on Hyperuricemic Rats Induced by Potassium Oxonate. Mar. Drugs 2019, 17, 343. [Google Scholar] [CrossRef]
- Coniglio, S.; Shumskaya, M.; Vassiliou, E. Unsaturated Fatty Acids and Their Immunomodulatory Properties. Biology 2023, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Jiang, H.; Mao, X. Identification and Anti-Hyperuricemic Activity of Xanthine Oxidase Inhibitory Peptides from Pacific White Shrimp and Swimming Crab Based on Molecular Docking Screening. J. Agric. Food Chem. 2023, 71, 1620–1627. [Google Scholar] [CrossRef]
- Wang, K.; Rong, G.; Gao, Y.; Wang, M.; Sun, J.; Sun, H.; Liao, X.; Wang, Y.; Li, Q.; Gao, W.; et al. Fluorous-Tagged Peptide Nanoparticles Ameliorate Acute Lung Injury via Lysosomal Stabilization and Inflammation Inhibition in Pulmonary Macrophages. Small 2022, 18, 2203432. [Google Scholar] [CrossRef]
- Li, M.; Lv, R.; Xu, X.; Ge, Q.; Lin, S. Tricholoma Matsutake-Derived Peptides Show Gastroprotective Effects against Ethanol-Induced Acute Gastric Injury. J. Agric. Food Chem. 2021, 69, 14985–14994. [Google Scholar] [CrossRef]
- Dalbeth, N.; Choi, H.K.; Joosten, L.A.B.; Khanna, P.P.; Matsuo, H.; Perez-Ruiz, F.; Stamp, L.K. Gout. Nat. Rev. Dis. Primers. 2019, 5, 69. [Google Scholar] [CrossRef]
- Cipolletta, E.; Tata, L.J.; Nakafero, G.; Avery, A.J.; Mamas, M.A.; Abhishek, A.J. Association Between Gout Flare and Subsequent Cardiovascular Events Among Patients with Gout. JAMA 2022, 328, 440–450. [Google Scholar] [CrossRef]
- Weaver, D.J., Jr. Uric acid and progression of chronic kidney disease. Pediatr. Nephrol. 2019, 34, 801–809. [Google Scholar] [CrossRef]
- Gherghina, M.E.; Peride, I.; Tiglis, M.; Neagu, T.P.; Niculae, A.; Checherita, I.A. Uric Acid and Oxidative Stress-Relationship with Cardiovascular, Metabolic, and Renal Impairment. Int. J. Mol. Sci. 2022, 23, 3188. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; Zeng, L.; Yin, S.; Hu, Y.; Li, S.; Fu, Y.; Zhang, X.; Xie, C.; Shu, L.; et al. RDP3, A Novel Antigout Peptide Derived from Water Extract of Rice. J. Agric. Food Chem. 2020, 68, 7143–7151. [Google Scholar] [CrossRef]
- Herman, R.A.; Yan, C.H.; Wang, J.Z.; Xun, X.M.; Wu, C.K.; Li, Z.N.; Ayepa, E.; You, S.; Gong, L.C.; Wang, J. Insight into the Silkworm Pupae: Modification Technologies and Functionality of the Protein and Lipids. Trends Food Sci. Technol. 2022, 129, 408–420. [Google Scholar] [CrossRef]
- Mahanta, D.K.; Komal, J.; Samal, I.; Bhoi, T.K.; Dubey, V.K.; Pradhan, K.; Nekkanti, A.; Gouda, M.N.R.; Saini, V.; Negi, N.; et al. Nutritional aspects and dietary benefits of “Silkworms”: Current scenario and future outlook. Front. Nutr. 2023, 10, 1121508. [Google Scholar] [CrossRef]
- Wu, X.; Yang, J.; Mumby, W.; Zhang, Y.; Zhang, Y.; Wang, C.; Chen, X.; Suo, H.; Song, J. Silkworm Pupa Protein and Its Peptides: Preparation, Biological Activity, Applications in Foods, and Advantages. Trends Food Sci. Technol. 2023, 139, 104129. [Google Scholar] [CrossRef]
- Choi, J.I.; Kweon, H.Y.; Lee, Y.L.; Lee, J.H.; Lee, S.Y. Efficacy of Silkworm Pupae Extract on Muscle Strength and Mass in Middle-Aged and Older Individuals: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Nutr. Health Aging. 2023, 27, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, S.; Xin, X.; Zhang, B.; Thomas, A.; Charles, A.; Lee, K.S.; Jin, B.R.; Gui, Z. Purification and Characterization of a Novel Immunomodulatory Hexapeptide from Alcalase Hydrolysate of Ultramicro-Pretreated Silkworm (Bombyx Mori) Pupa Protein. J. Asia-Pac. Entomol. 2019, 22, 633–637. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Zhu, Z.; Li, X.; Sun, S.; Wang, W.; Sadiq, F.A. Identification and Characterization of Two Novel Antioxidant Peptides from Silkworm Pupae Protein Hydrolysates. Eur. Food Res. Technol. 2021, 247, 343–352. [Google Scholar] [CrossRef]
- Ye, J.; Li, J.; Zhao, P. The Silkworm Carboxypeptidase Inhibitor Prevents Gastric Cancer Cells’ Proliferation through the EGF/EGFR Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 1078. [Google Scholar] [CrossRef]
- Altomare, A.A.; Baron, G.; Aldini, G.; Carini, M.; D’Amato, A. Silkworm pupae as source of high-value edible proteins and of bioactive peptides. Food. Sci. Nutr. 2020, 8, 2652–2661. [Google Scholar] [CrossRef]
- Lu, Y.; Li, S.; Xu, H.; Zhang, T.; Lin, X.; Wu, X. Effect of Covalent Interaction with Chlorogenic Acid on the Allergenic Capacity of Ovalbumin. J. Agric. Food Chem. 2018, 66, 9794–9800. [Google Scholar] [CrossRef] [PubMed]
- Takahama, U.; Koga, Y.; Hirota, S.; Yamauchi, R. Inhibition of Xanthine Oxidase Activity by an Oxathiolanone Derivative of Quercetin. Food Chem. 2011, 126, 1808–1811. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, D.; Liu, J.; Li, G.; Zhang, Z.; Chen, C.; Zhang, L.; Li, J. Characterization of Xanthine Oxidase Inhibitory Activities of Phenols from Pickled Radish with Molecular Simulation. Food Chem. X 2022, 14, 100343. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Yang, M.; Liu, N.; Liu, Y.; Sun, H.; Wang, S.; Zhang, Y.; Li, Y.; Yang, X.; Wang, Y. Short Hexapeptide Optimized from Rice-Derived Peptide 1 Shows Promising Anti-Hyperuricemia Activities. J. Agric. Food Chem. 2022, 70, 6679–6687. [Google Scholar] [CrossRef]
- Kimura, H.; Francisco, D.; Conway, M.; Martinez, F.D.; Vercelli, D.; Polverino, F.; Billheimer, D.; Kraft, M. Type 2 Inflammation Modulates ACE2 and TMPRSS2 in Airway Epithelial Cells. J. Allergy Clin. Immunol. 2020, 146, 80–88. [Google Scholar] [CrossRef]
- Qi, X.; Ma, Y.; Guan, K.; Liu, C.; Wang, R.; Ma, Y.; Niu, T. Whey Protein Peptide PEW Attenuates Hyperuricemia and Associated Renal Inflammation in Potassium Oxonate and Hypoxanthine-Induced Rat. Food Biosci. 2023, 51, 102311. [Google Scholar] [CrossRef]
- Zhao, Q.; Jiang, X.; Mao, Z.; Zhang, J.; Sun, J.; Mao, X. Exploration, Sequence Optimization and Mechanism Analysis of Novel Xanthine Oxidase Inhibitory Peptide from Ostrea Rivularis Gould. Food Chem. 2023, 404, 134537. [Google Scholar] [CrossRef]
- Hou, M.; Xiang, H.; Hu, X.; Chen, S.; Wu, Y.; Xu, J.; Yang, X. Novel Potential XOD Inhibitory Peptides Derived from Trachinotus Ovatus: Isolation, Identification and Structure-Function Analysis. Food Biosci. 2022, 47, 101639. [Google Scholar] [CrossRef]
- Hao, L.; Ding, Y.; Fan, Y.; Tian, Q.; Liu, Y.; Guo, Y.; Zhang, J.; Hou, H. Identification of Hyperuricemia Alleviating Peptides from Yellow Tuna Thunnus Albacares. J. Agric. Food Chem. 2024, 12, 12083–12099. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Li, C.; Zhao, Y.; Xu, L.; Ma, S.; Lin, F.; Xie, Y.; An, J.; Wang, S. Paeonia × suffruticosa Andrews Leaf Extract and Its Main Component Apigenin 7-O-Glucoside Ameliorate Hyperuricemia by Inhibiting Xanthine Oxidase Activity and Regulating Renal Urate Transporters. Phytomedicine 2023, 118, 154957. [Google Scholar] [CrossRef]
- Takir, M.; Kostek, O.; Ozkok, A.; Elcioglu, O.C.; Bakan, A.; Erek, A.; Mutlu, H.H.; Telci, O.; Semerci, A.; Odabas, A.R.; et al. Lowering Uric Acid with Allopurinol Improves Insulin Resistance and Systemic Inflammation in Asymptomatic Hyperuricemia. J. Investig. Med. 2015, 63, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Ro, K.S.; Zhao, L.; Hu, Y.; Ge, M.; Du, L.; Xie, J. Anti-Hyperuricemic Properties and Mechanism of Lactiplantibacillus Plantarum X7023. Process Biochem. 2024, 136, 26–37. [Google Scholar] [CrossRef]
- Xie, D.; Shen, Y.; Su, E.; Du, L.; Xie, J.; Wei, D. Anti-Hyperuricemic, Nephroprotective, and Gut Microbiota Regulative Effects of Separated Hydrolysate of α-Lactalbumin on Potassium Oxonate- and Hypoxanthine-Induced Hyperuricemic Mice. Mol. Nutr. Food Res. 2023, 67, 2200162. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.; Zhong, H.; Chen, F.; Regenstein, J.; Hu, X.; Cai, L.; Feng, F. The Gut Microbiota as a Target to Control Hyperuricemia Pathogenesis: Potential Mechanisms and Therapeutic Strategies. Crit. Rev. Food Sci. Nutr. 2022, 62, 3979–3989. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Mass | Length | Peptide Score | Relative Intensity (%) | Area |
|---|---|---|---|---|---|
| YHFGVPVG | 874.4337 | 8 | 875.4418 | 12.39 | 11,100,000 |
| FITPF | 623.3318 | 5 | 624.3403 | 16.80 | 7,740,000 |
| FLTPF | 623.3318 | 5 | 624.3403 | 16.80 | 7,740,000 |
| WFITPF | 809.4112 | 6 | 810.4222 | 21.32 | 7,280,000 |
| FFIYNREYNDALKLG | 1861.936 | 15 | 931.9765 | 14.91 | 6,790,000 |
| FNILVR | 760.4595 | 6 | 761.4681 | 12.27 | 4,550,000 |
| FNLLVR | 760.4595 | 6 | 761.4681 | 12.27 | 4,550,000 |
| FIPEYLS | 867.4378 | 7 | 868.4468 | 15.03 | 3,610,000 |
| GALFLQDNLVK | 1216.682 | 11 | 1217.689 | 14.83 | 3,320,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Yang, Z.; Lin, X.; Xu, Z.; Mu, L.; Li, Q.; Wu, X. Anti-Hyperuricemic and Nephroprotective Effects of Hydrolysate Derived from Silkworm Pupae (Bombyx mori): In Vitro and In Vivo Study. Nutrients 2025, 17, 1596. https://doi.org/10.3390/nu17091596
Fan Y, Yang Z, Lin X, Xu Z, Mu L, Li Q, Wu X. Anti-Hyperuricemic and Nephroprotective Effects of Hydrolysate Derived from Silkworm Pupae (Bombyx mori): In Vitro and In Vivo Study. Nutrients. 2025; 17(9):1596. https://doi.org/10.3390/nu17091596
Chicago/Turabian StyleFan, Yuting, Zhencong Yang, Xiao Lin, Zhoujin Xu, Lixia Mu, Qingrong Li, and Xuli Wu. 2025. "Anti-Hyperuricemic and Nephroprotective Effects of Hydrolysate Derived from Silkworm Pupae (Bombyx mori): In Vitro and In Vivo Study" Nutrients 17, no. 9: 1596. https://doi.org/10.3390/nu17091596
APA StyleFan, Y., Yang, Z., Lin, X., Xu, Z., Mu, L., Li, Q., & Wu, X. (2025). Anti-Hyperuricemic and Nephroprotective Effects of Hydrolysate Derived from Silkworm Pupae (Bombyx mori): In Vitro and In Vivo Study. Nutrients, 17(9), 1596. https://doi.org/10.3390/nu17091596