
Lactobacillus fermentum LF31 Supplementation Reversed Atrophy Fibers in a Model of Myopathy Through the Modulation of IL-6, TNF-α, and Hsp60 Levels Enhancing Muscle Regeneration

 ,
,  ,
,  , ,
, ,  ,
,  , ,
, ,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. The Model System: Animals and Diets
2.2. Histopathology
2.3. Immunofluorescence
2.4. Immunohistochemistry
2.5. Enzyme-Linked Immunoadsorbent Assay (ELISA)
2.6. Immunoblotting
2.7. Statistical Analysis
3. Results
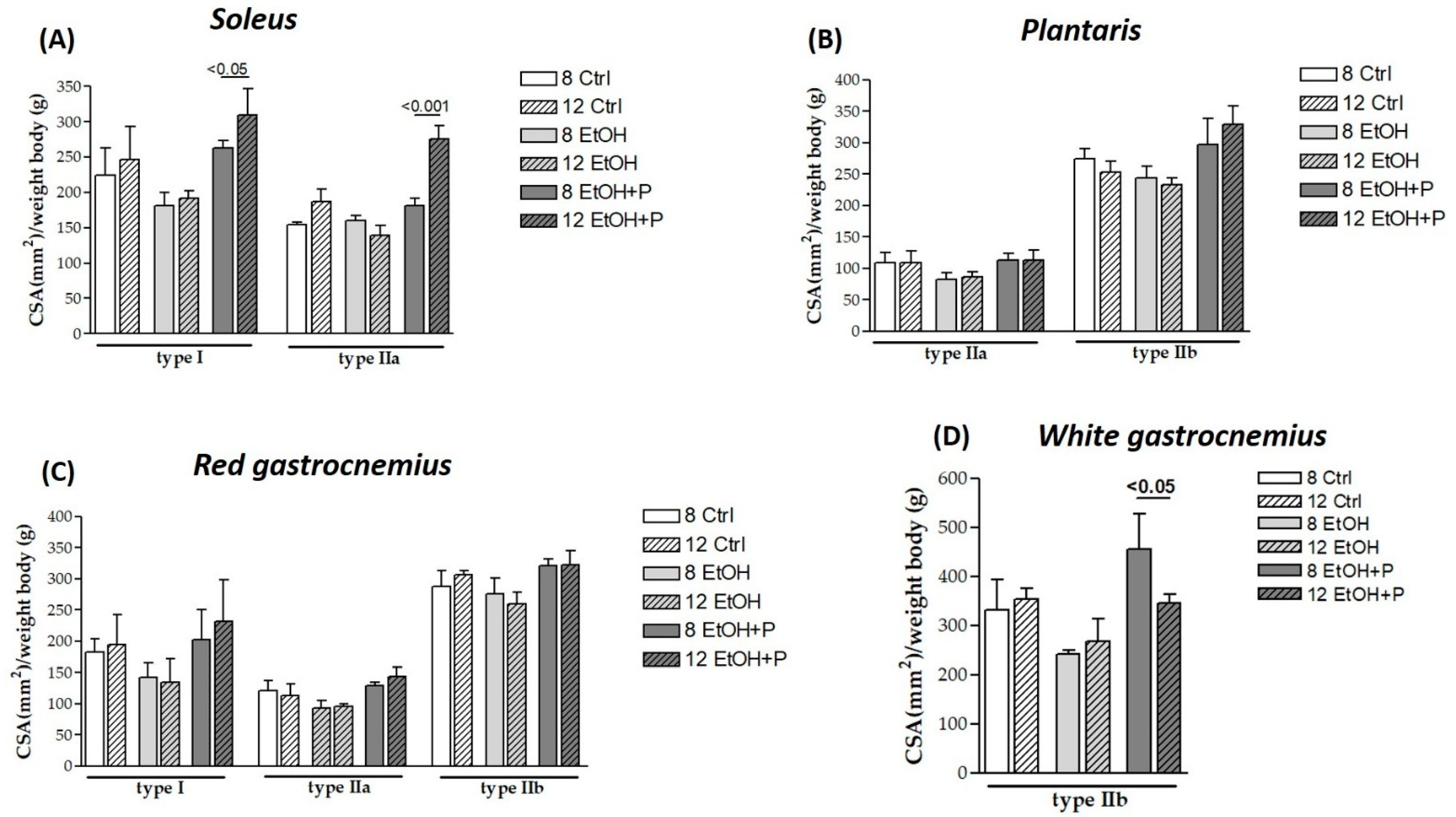
3.1. Effects of L. fermentum in Various Fiber Types
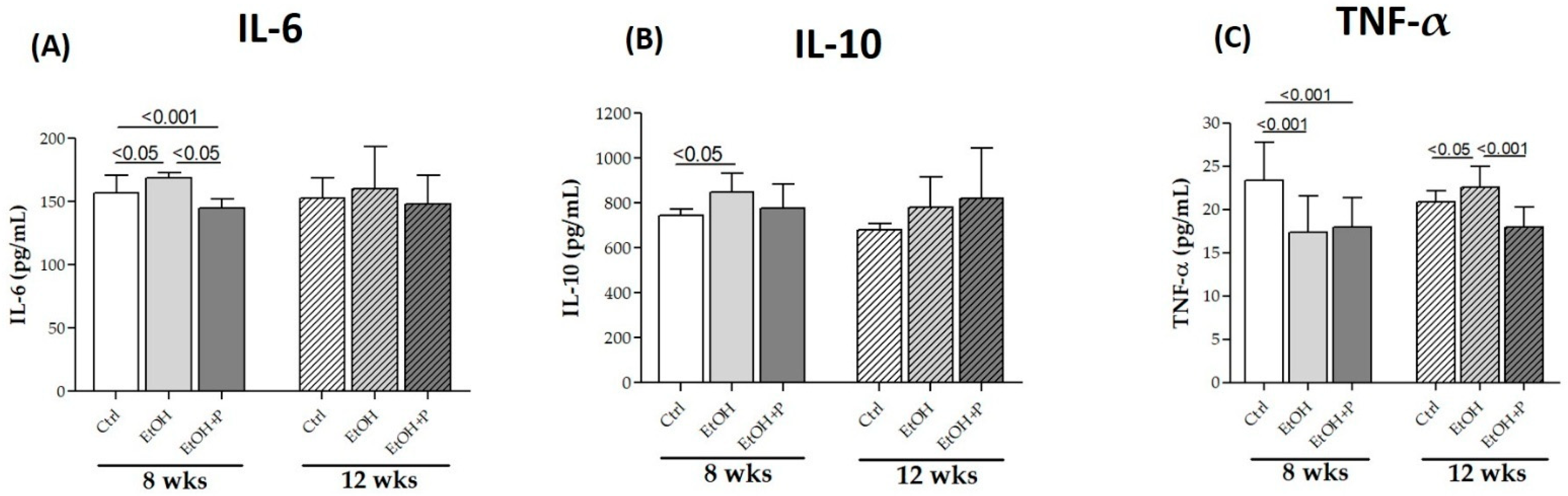
3.2. L. fermentum Attenuates Inflammation and Oxidative Stress Induced by Chronic Ethanol Consumption
3.3. Administration of L. fermentum Increases the Expression of Pax7 and MyoD in SCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SCs | Satellite cells |
| EtOH | Ethanol |
| MHC | Myosin heavy chain |
| IL | Interleukin |
| TNF-α | Tumor necrosis factor alpha |
| Hsp60 | Heat shock protein |
| LPSs | Lipopolysaccharides |
| ROS | Reactive oxygen species |
| E & E | Hematoxylin eosin |
| CSA | Cross-section area |
| IkB | IkappaB kinase |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| IGF-1 | Insulin-like growth factor 1 |
References
- Sausa, M.; Fucarino, A.; Paladino, L.; Zummo, F.P.; Fabbrizio, A.; Di Felice, V.; Rappa, F.; Barone, R.; Marino Gammazza, A.; Macaluso, F. Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis. Biomedicines 2024, 12, 382. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Bucchieri, F.; Cappello, F. The unexplored potential of exosomes in the muscle-brain axis. Proc. Natl. Acad. Sci. USA 2025, 7, e2420766121. [Google Scholar]
- Mostosi, D.; Molinaro, M.; Saccone, S.; Torrente, Y.; Villa, C.; Farini, A. Exploring the Gut Microbiota-Muscle Axis in Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2024, 21, 5589. [Google Scholar]
- Ticinesi, A.; Milani, C.; Lauretani, F.; Nouvenne, A.; Mancabelli, L.; Lugli, G.A.; Turroni, F.; Duranti, S.; Mangifesta, M.; Viappiani, A.; et al. Gut microbiota composition is associated with polypharmacy in elderly hospitalized patients. Sci. Rep. 2017, 11, 11102. [Google Scholar]
- Liu, C.; Cheung, W.H.; Li, J.; Chow, S.K.; Yu, J.; Wong, S.H.; Ip, M.; Sung, J.J.Y.; Wong, R.M.Y. Understanding the gut microbiota and sarcopenia: A systematic review. JCSM 2021, 12, 1393–1407. [Google Scholar]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and Gut-Derived Inflammation. Alcohol. Res. 2017, 38, 163–171. [Google Scholar] [PubMed]
- Engen, P.A.; Green, S.J.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. The Gastrointestinal Microbiome: Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol. Res. 2015, 37, 223–236. [Google Scholar]
- Leclercq, S.; Matamoros, S.; Cani, P.D.; Neyrinck, A.M.; Jamar, F.; Stärkel, P.; Windey, K.; Tremaroli, V.; Bäckhed, F.; Verbeke, K.; et al. Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity. Proc. Natl. Acad. Sci. USA 2014, 111, E4485–E4493. [Google Scholar]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif. Tissue Int. 2018, 102, 433–442. [Google Scholar]
- Wang, Y.; Li, Y.; Bo, L.; Zhou, E.; Chen, Y.; Naranmandakh, S.; Xie, W.; Ru, Q.; Chen, L.; Zhu, Z.; et al. Progress of linking gut microbiota and musculoskeletal health: Casualty, mechanisms, and translational values. Gut Microbes 2023, 15, 2263207. [Google Scholar]
- Jäger, R.; Purpura, M.; Farmer, S.; Cash, H.A.; Keller, D. Probiotic Bacillus coagulans GBI-30, 6086 Improves Protein Absorption and Utilization. Probiotics Antimicrob. Proteins 2018, 10, 611–615. [Google Scholar]
- Lapauw, L.; Rutten, A.; Dupont, J.; Amini, N.; Vercauteren, L.; Derrien, M.; Raes, J.; Gielen, E. Associations between gut microbiota and sarcopenia or its defining parameters in older adults: A systematic review. CJSM 2024, 15, 2190–2207. [Google Scholar]
- Lang, C.H.; Pruznak, A.M.; Nystrom, G.J.; Vary, T.C. Alcohol-induced decrease in muscle protein synthesis associated with increased binding of mTOR and raptor: Comparable effects in young and mature rats. Nutr. Metab. 2009, 20, 4. [Google Scholar]
- Steiner, J.L.; Lang, C.H. Dysregulation of skeletal muscle protein metabolism by alcohol. Am. J. Physiol. Endocrinol. Metab. 2015, 1, E699–E712. [Google Scholar]
- Shenkman, B.S.; Belova, S.P.; Zinovyeva, O.E.; Samkhaeva, N.D.; Mirzoev, T.M.; Vilchinskaya, N.A.; Altaeva, E.G.; Turtikova, O.V.; Kostrominova, T.Y.; Nemirovskaya, T.L. Effect of Chronic Alcohol Abuse on Anabolic and Catabolic Signaling Pathways in Human Skeletal Muscle. Alcohol. Clin. Exp. Res. 2018, 42, 41–52. [Google Scholar] [PubMed]
- Wu, D.; Cederbaum, A.I. Alcohol, oxidative stress, and free radical damage. Alcohol. Res. Health 2003, 27, 277–284. [Google Scholar] [PubMed]
- Clark, A.; Mach, N. The crosstalk between the gut microbiota and mitochondria during exercise. Front. Physiol. 2017, 8, 319. [Google Scholar]
- Levitt, D.E.; Yeh, A.Y.; Prendergast, M.J.; Jr, R.G.B.; Adler, K.A.; Cook, G.; Molina, P.E.; Simon, L. Chronic Alcohol Dysregulates Skeletal Muscle Myogenic Gene Expression after Hind Limb Immobilization in Female Rats. Biomolecules 2020, 12, 441. [Google Scholar]
- Carbone, J.W.; Pasiakos, S.M. Dietary Protein and Muscle Mass: Translating Science to Application and Health Benefit. Nutrients 2019, 22, 1136. [Google Scholar]
- Soccol, C.R.; Porto de Souza Vandenberghe, L.; Spier, M.R.; Medeiros, A.B.P.; Yamaguishi, C.T.; De Dea Lindner, J.; Pandey, A.; Soccol, V.T. The potential of Probiotics: A Review. Food Technol. Biotechnol. 2010, 48, 413–434. [Google Scholar]
- Plaza-Diaz, J. Beneficial effects of rosemary, sage and propolis on human health. Nutrients 2018, 10, 1629. [Google Scholar]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 1, 5804–5815. [Google Scholar]
- Paladino, L.; Rappa, F.; Barone, R.; Macaluso, F.; Zummo, F.P.; Davide, S.; Szychlinska, M.A.; Bucchieri, F.; Conway di Macario, E.; Macario, A.J.L.; et al. NF-κB Regulation and the Chaperone System Mediate Restorative Effects of the Probiotic Lactobacillus fermentum LF31 in the Small Intestine and Cerebellum of Mice with Ethanol-Induced Damage. Biology 2023, 12, 1394. [Google Scholar]
- Carr, R.M.; Dhir, R.; Yin, X.; Agarwal, B.; Ahima, R.S. Temporal effects of ethanol consumption on energy homeostasis, hepatic steatosis, and insulin sensitivity in mice. Alcohol. Clin. Exp. Res. 2013, 37, 1091–1099. [Google Scholar]
- Carr, R.M.; Peralta, G.; Yin, X.; Ahima, R.S. Absence of perilipin 2 prevents hepatic steatosis, glucose intolerance and ceramide accumulation in alcohol-fed mice. PLoS ONE 2014, 9, e97118. [Google Scholar]
- Barone, R.; Rappa, F.; Macaluso, F.; Caruso Bavisotto, C.; Sangiorgi, C.; Di Paola, G.; Tomasello, G.; Di Felice, V.; Marcianò, V.; Farina, F.; et al. Alcoholic Liver Disease: A Mouse Model Reveals Protection by Lactobacillus fermentum. Clin. Transl. Gastroenterol. 2016, 7, e138. [Google Scholar]
- Fernandez-Solà, J.; Preedy, V.R.; Lang, C.H.; Gonzalez-Reimers, E.; Arno, M.; Lin, J.C.; Wiseman, H.; Zhou, S.; Emery, P.W.; Nakahara, T.; et al. Molecular and cellular events in alcohol-induced muscle disease. Alcohol. Clin. Exp. Res. 2007, 31, 1953–1962. [Google Scholar] [PubMed]
- Urbano-Márquez, A.; Fernández-Solà, J. Effects of alcohol on skeletal and cardiac muscle. Muscle Nerve 2004, 30, 689–707. [Google Scholar]
- Stožer, A.; Vodopivc, P.; Križančić, B.L. Pathophysiology of exercise-induced muscle damage and its structural, functional, metabolic, and clinical consequences. Physiol. Res. 2020, 31, 565–598. [Google Scholar]
- Siparsky, P.N.; Kirkendall, D.T.; Garrett, W.E., Jr. Muscle changes in aging: Understanding sarcopenia. Sports Health 2014, 6, 36–40. [Google Scholar] [PubMed]
- Ørngreen, M.C.; Vissing, J. Treatment Opportunities in Patients with Metabolic Myopathies. Curr. Treat. Options Neurol. 2017, 21, 37. [Google Scholar]
- Fucarino, A.; Burgio, S.; Paladino, L.; Caruso Bavisotto, C.; Pitruzzella, A.; Bucchieri, F.; Cappello, F. The Microbiota Is Not an Organ: Introducing the Muco-Microbiotic Layer as a Novel Morphofunctional Structure. Anatomia 2022, 1, 186–203. [Google Scholar] [CrossRef]
- Cappello, F.; Saguto, D.; Burgio, S.; Paladino, L.; Bucchieri, F. Does Intestine Morphology Still Have Secrets to Reveal? A Proposal about the “Ghost” Layer of the Bowel. Appl. Biosci. 2022, 1, 95–100. [Google Scholar] [CrossRef]
- Cappello, F.; Gratie, M.I. Unveiling the muco-microbiotic layer: A three-component framework for mucosal health and disease. J. Biol. Res. Boll. Della Soc. Ital. Di Biol. Sper. 2025. [Google Scholar]
- Manna, O.M.; Caruso Bavisotto, C.; Gratie, M.I.; Damiani, P.; Bonaventura, G.; Cappello, F.; Tomasello, G.; D’Andrea, V. Targeting Helicobacter pylori Through the “Muco-Microbiotic Layer” Lens: The Challenge of Probiotics and Microbiota Nanovesicles. Nutrients 2025, 3, 569. [Google Scholar]
- Cappello, F.; Rappa, F.; Canepa, F.; Carini, F.; Mazzola, M.; Tomasello, G.; Bonaventura, G.; Giuliana, G.; Leone, A.; Saguto, D.; et al. Probiotics Can Cure Oral Aphthous-Like Ulcers in Inflammatory Bowel Disease Patients: A Review of the Literature and a Working Hypothesis. Int. J. Mol. Sci. 2019, 11, 5026. [Google Scholar]
- Steiner, J.L.; Lang, C.H. Alcohol impairs skeletal muscle protein synthesis and mTOR signaling in a time-dependent manner following electrically stimulated muscle contraction. J. Appl. Physiol. 2014, 15, 1170–1179. [Google Scholar]
- Bloemberg, D.; Quadrilatero, J. Rapid determination of myosin heavy chain expression in rat, mouse, and human skeletal muscle using multicolor immunofluorescence analysis. PLoS ONE 2012, 7, e35273. [Google Scholar]
- Walsh, M.E.; Bhattacharya, A.; Sataranatarajan, K.; Qaisar, R.; Sloane, L.; Rahman, M.M.; Kinter, M.; Van Remmen, H. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 2015, 14, 957–970. [Google Scholar]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [PubMed]
- Das, S.K.; Vasudevan, D.M. Alcohol-induced oxidative stress. Life Sci. 2007, 27, 177–187. [Google Scholar]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell Mol. Med. JCMM 2010, 14, 840–860. [Google Scholar] [PubMed]
- Reeg, S.; Jung, T.; Castro, J.P.; Davies, K.J.A.; Henze, A.; Grune, T. The molecular chaperone Hsp70 promotes the proteolytic removal of oxidatively damaged proteins by the proteasome. Free Radic. Biol. Med. 2016, 99, 153–166. [Google Scholar]
- Gething, M.J.; Sambrook, J. Protein folding in the cell. Nature 1992, 355, 33–45. [Google Scholar]
- Marino Gammazza, A.; Macaluso, F.; Di Felice, V.; Cappello, F.; Barone, R. Hsp60 in Skeletal Muscle Fiber Biogenesis and Homeostasis: From Physical Exercise to Skeletal Muscle Pathology. Cells 2018, 22, 224. [Google Scholar]
- Zhao, T.; Wang, H.; Liu, Z.; Liu, Y.; DeJi; Li, B.; Huang, X. Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease. Antioxidants 2023, 12, 769. [Google Scholar] [CrossRef]
- Samali, A.; Cai, J.; Zhivotovsky, B.; Jones, D.P.; Orrenius, S. Presence of a pre-apoptotic complex of pro-caspase-3, Hsp60 and Hsp10 in the mitochondrial fraction of jurkat cells. EMBO J. 1999, 18, 2040–2048. [Google Scholar]
- Pockley, A.G.; Muthana, M.; Calderwood, S.K. The dual immunoregulatory roles of stress proteins. Trends Biochem. Sci. 2008, 33, 71–79. [Google Scholar]
- Quintana, F.J.; Cohen, I.R. HSP60 speaks to the immune system in many voices. Novartis Found. Symp. 2008, 291, 101–111. [Google Scholar]
- Chun, J.N.; Choi, B.; Lee, K.W.; Lee, D.J.; Kang, D.H.; Lee, J.Y.; Song, I.S.; Kim, H.I.; Lee, S.H.; Kim, H.S.; et al. Cytosolic HSP60 is involved in the NF-kappaB-dependent survival of cancer cells via IKK regulation. PLoS ONE 2010, 5, e9422. [Google Scholar]
- Min, S.; Kim, J.Y.; Cho, H.M.; Park, S.; Hwang, J.M.; You, H.; Chae, Y.C.; Lee, W.J.; Sun, W.; Kang, W.; et al. Heat shock protein 60 couples an oxidative stress-responsive p38/MK2 signaling and NF-κB survival machinery in cancer cells. Redox Biol. 2022, 51, 102293. [Google Scholar]
- Blaine, S.K.; Ridner, C.; Mcampbell, B.R.; Crone, L.; Claus, E.D.; Wilson, J.R.; West, S.N.; McClanahan, A.J.; Siddiq, A.S.; Layman, I.M.P.; et al. IL-6, but not TNF-α, response to alcohol cues and acute consumption associated with neural cue reactivity, craving, and future drinking in binge drinkers. Brain Behav. Immun. Health 2023, 31, 100645. [Google Scholar] [PubMed]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar]
- Opp, M.R.; Smith, E.M.; Hughes, T.K., Jr. Interleukin-10 (cytokine synthesis inhibitory factor) acts in the central nervous system of rats to reduce sleep. J. Neuroimmunol. 1995, 60, 165–168. [Google Scholar]
- Carlini, V.; Noonan, D.M.; Abdalalem, E.; Goletti, D.; Sansone, C.; Calabrone, L.; Albini, A. The multifaceted nature of IL-10: Regulation, role in immunological homeostasis and its relevance to cancer, COVID-19 and post-COVID conditions. Front. Immunol. 2023, 8, 1161067. [Google Scholar]
- Chen, L.H.; Huang, S.Y.; Huang, K.C.; Hsu, C.C.; Yang, K.C.; Li, L.A.; Chan, C.H.; Huang, H.Y. Lactobacillus paracasei PS23 decelerated age-related muscle loss by ensuring mitochondrial function in SAMP8 mice. Aging 2019, 29, 756–770. [Google Scholar]
- Seo, E.; Truong, C.S.; Jun, H.S. Psoralea corylifolia L. seed extract attenuates dexamethasone-induced muscle atrophy in mice by inhibition of oxidative stress and inflammation. J. Ethnopharmacol. 2022, 296, 115490. [Google Scholar]
- Cáceres-Ayala, C.; Mira, R.G.; Acuña, M.J.; Brandan, E.; Cerpa, W.; Rebolledo, D.L. Episodic Binge-like Ethanol Reduces Skeletal Muscle Strength Associated with Atrophy, Fibrosis, and Inflammation in Young Rats. Int. J. Mol. Sci. 2023, 14, 1655. [Google Scholar]
- Levitt, D.E.; Chalapati, N.; Prendergast, M.J.; Simon, L.; Molina, P.E. Ethanol-Impaired Myogenic Differentiation is Associated with Decreased Myoblast Glycolytic Function. Alcohol. Clin. Exp. Res. 2020, 44, 2166–2176. [Google Scholar]
- Abdel-Halim, N.H.M.; Farrag, E.A.E.; Hammad, M.O.; Habotta, O.A.; Hassan, H.M. Probiotics Attenuate Myopathic Changes in Aging Rats via Activation of the Myogenic Stellate Cells. Probiotics Antimicrob. Proteins 2023, 19, 1–12. [Google Scholar]
- Park, J.-Y.; Lee, J.Y.; Kim, Y.; Kang, C.-H. Limosilactobacillus fermentum MG5091 and Lactococcus lactis MG4668 and MG5474 Suppress Muscle Atrophy by Regulating Apoptosis in C2C12 Cells. Fermentation 2023, 9, 659. [Google Scholar] [CrossRef]
- Lehtinen, S.K.; Rahkila, P.; Helenius, M.; Korhonen, P.; Salminen, A. Down-regulation of transcription factors AP-1, Sp-1, and NF-kappa B precedes myocyte differentiation. Biochem. Biophys. Res. Commun. 1996, 4, 36–43. [Google Scholar]
- Hor, Y.Y.; Ooi, C.H.; Lew, L.C.; Jaafar, M.H.; Lau, A.S.; Lee, B.K.; Azlan, A.; Choi, S.B.; Azzam, G.; Liong, M.T. The molecular mechanisms of probiotic strains in improving ageing bone and muscle of d-galactose-induced ageing rats. J. Appl. Microbiol. 2021, 130, 1307–1322. [Google Scholar] [PubMed]
- Guttridge, D.C.; Albanese, C.; Reuther, J.Y.; Pestell, R.G.; Baldwin, A.S., Jr. NF-kappaB controls cell growth and differentiation through transcriptional regulation of cyclin D1. Mol. Cell Biol. 1999, 19, 5785–5799. [Google Scholar]
- Varian, B.J.; Gourishetti, S.; Poutahidis, T.; Lakritz, J.R.; Levkovich, T.; Kwok, C.; Teliousis, K.; Ibrahim, Y.M.; Mirabal, S.; Erdman, S.E. Beneficial bacteria inhibit cachexia. Oncotarget 2016, 7, 11803–11816, Erratum in: Oncotarget 2018, 29, 29536. [Google Scholar]
- Van Krimpen, S.J.; Jansen, F.A.C.; Ottenheim, V.L.; Belzer, C.; van der Ende, M.; van Norren, K. The Effects of Pro-, Pre-, and Synbiotics on Muscle Wasting, a Systematic Review-Gut Permeability as Potential Treatment Target. Nutrients 2021, 29, 1115. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Features | IHC | IF | WB |
|---|---|---|---|---|
| Paired box 7 (Pax7) | 5081-MSM1-P1ABX Mouse Monoclonal NeoBiotecnhologies, Union City, CA, USA | - | 1:200 | 1:1000 |
| Myoblast determination protein 1 (MyoD) | MA1-41017 Mouse Monoclonal Invitrogen, Waltham, MA, USA | - | 1:300 | 1:1000 |
| Actin Recombinant (JJ09-29) | MA5-32479 Rabbit Monoclonal Invitrogen, Waltham, MA, USA | - | - | 1:1000 |
| Laminin | AB2034 Rabbit Polyclonal Millipore, Chicago, IL, USA | - | 1:50 | - |
| Heat Shock Protein 60 (Hsp60) | Sc-1722 Goat Polyclonal Santa Cruz, CA, USA | - | 1:50 | - |
| Type I Myosin Heavy Chain (MHC-I) | A4.951-s Mouse Monoclonal DSHB, Lowa City, IA, USA | 1:50 | - | |
| Type IIa Myosin Heavy Chain (MHC-IIa) | SC-71-s Mouse Monoclonal DSHB, Lowa City, IA, USA | 1:50 | - | |
| Type IIb Myosin Heavy Chain (MHC-IIb) | BF-F3-s Mouse Monoclonal DSHB, Lowa City, IA, USA | 1:50 | - |
| Target | Features | Sample Dilution | Limit of Detection | Precision |
|---|---|---|---|---|
| Mouse IL-6 | LS-F263 LS Bio, Lynnwood, WA, USA | No | 0.82–600 pg/mL | Intra-Assay: CV < 10% Inter-Assay: CV < 12% |
| Mouse IL-10 | LS-F253 LS Bio, Lynnwood, WA, USA | No | 45–5000 pg/mL | Intra-Assay: CV < 10% Inter-Assay: CV < 12% |
| Mouse HSPD1 | LS-F4239 LS Bio, Lynnwood, WA, USA | No | 4.56–100 ng/mL | Intra-Assay: CV < 10% Inter-Assay: CV < 12% |
| Mouse TNF alpha | LS-F5192 LS Bio, Lynnwood, WA, USA | No | 15.6–1000 pg/mL | Intra-Assay: CV < 10% Inter-Assay: CV < 12% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sausa, M.; Paladino, L.; Scalia, F.; Zummo, F.P.; Vergilio, G.; Rappa, F.; Cappello, F.; Gratie, M.I.; Proia, P.; Di Felice, V.; et al. Lactobacillus fermentum LF31 Supplementation Reversed Atrophy Fibers in a Model of Myopathy Through the Modulation of IL-6, TNF-α, and Hsp60 Levels Enhancing Muscle Regeneration. Nutrients 2025, 17, 1550. https://doi.org/10.3390/nu17091550
Sausa M, Paladino L, Scalia F, Zummo FP, Vergilio G, Rappa F, Cappello F, Gratie MI, Proia P, Di Felice V, et al. Lactobacillus fermentum LF31 Supplementation Reversed Atrophy Fibers in a Model of Myopathy Through the Modulation of IL-6, TNF-α, and Hsp60 Levels Enhancing Muscle Regeneration. Nutrients. 2025; 17(9):1550. https://doi.org/10.3390/nu17091550
Chicago/Turabian StyleSausa, Martina, Letizia Paladino, Federica Scalia, Francesco Paolo Zummo, Giuseppe Vergilio, Francesca Rappa, Francesco Cappello, Melania Ionelia Gratie, Patrizia Proia, Valentina Di Felice, and et al. 2025. "Lactobacillus fermentum LF31 Supplementation Reversed Atrophy Fibers in a Model of Myopathy Through the Modulation of IL-6, TNF-α, and Hsp60 Levels Enhancing Muscle Regeneration" Nutrients 17, no. 9: 1550. https://doi.org/10.3390/nu17091550
APA StyleSausa, M., Paladino, L., Scalia, F., Zummo, F. P., Vergilio, G., Rappa, F., Cappello, F., Gratie, M. I., Proia, P., Di Felice, V., Marino Gammazza, A., Macaluso, F., & Barone, R. (2025). Lactobacillus fermentum LF31 Supplementation Reversed Atrophy Fibers in a Model of Myopathy Through the Modulation of IL-6, TNF-α, and Hsp60 Levels Enhancing Muscle Regeneration. Nutrients, 17(9), 1550. https://doi.org/10.3390/nu17091550