
Dietary Fibre Modulates Gut Microbiota in Late Pregnancy Without Altering SCFA Levels, and Propionate Treatement Has No Effect on Placental Explant Function

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Microbiota—Composition
2.3. Serum—Short-Chain Fatty Acids
2.4. Short-Chain-Fatty-Acid Treatment of Placental Explants
2.5. Mitochondrial DNA Content and Gene Expression
2.6. Protein—Extraction and Western Blotting
2.7. Statistical Analysis
3. Results
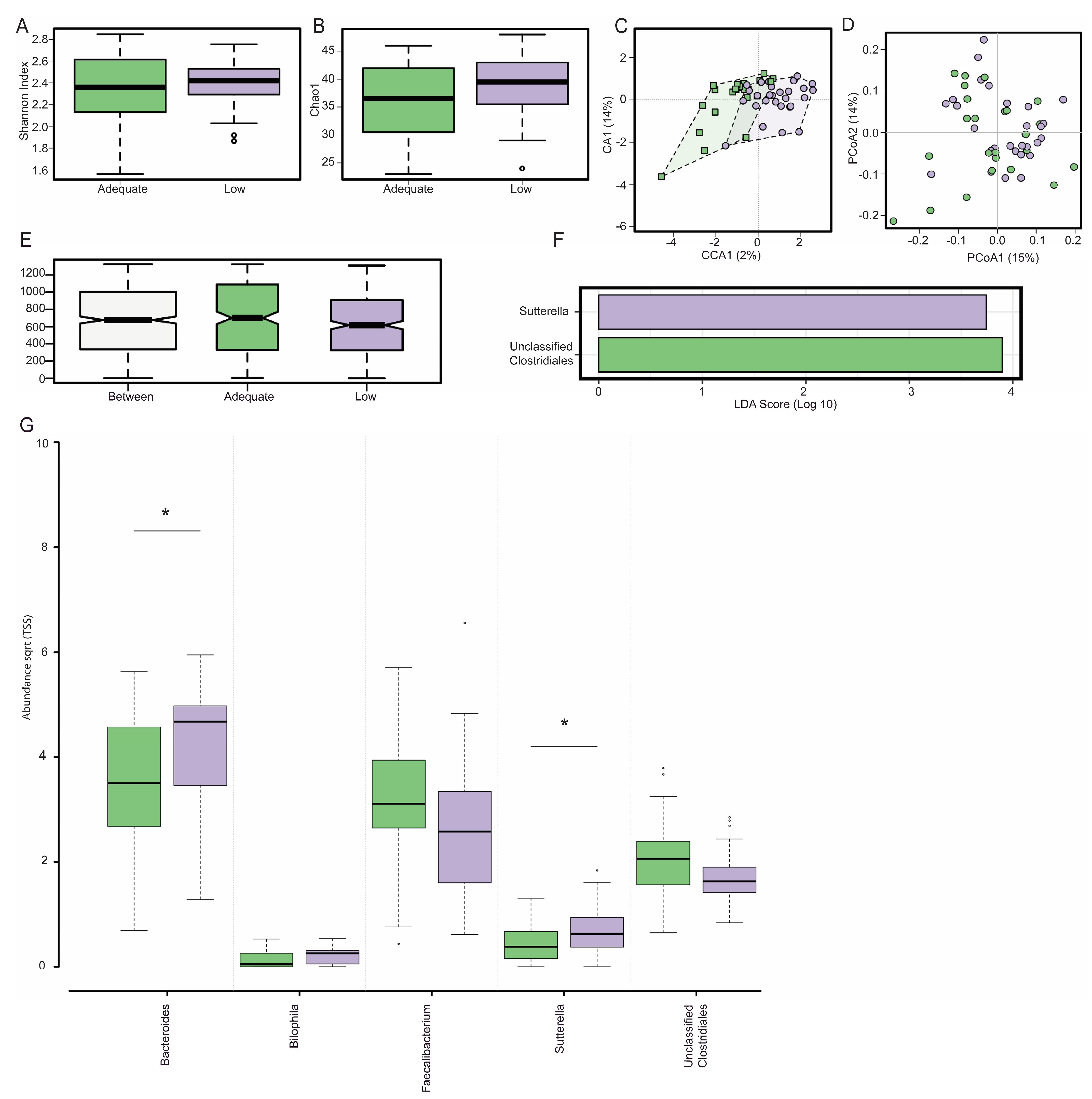
3.1. Gut Microbiota Composition
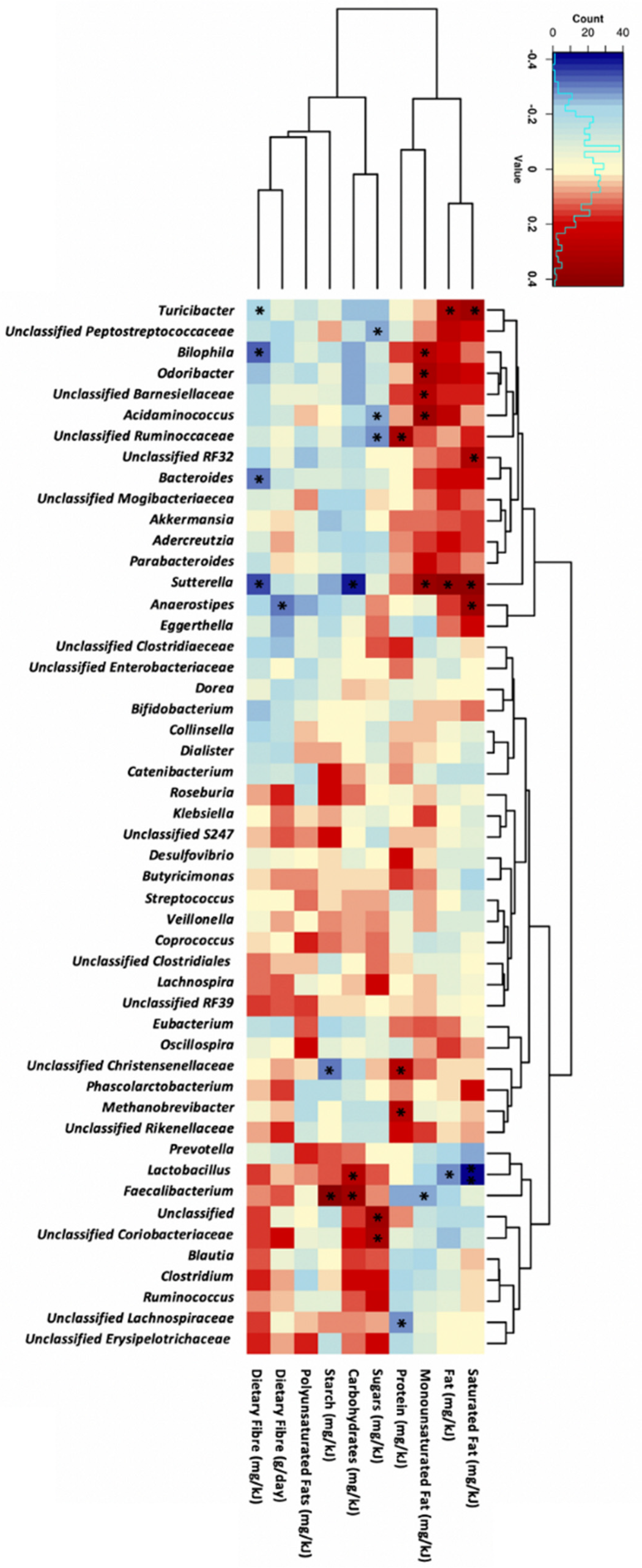
3.2. Dietary Fibre Intake and Genera Abundance
3.3. Macronutrient Intake and Genera Abundance
3.4. qPCR Validation
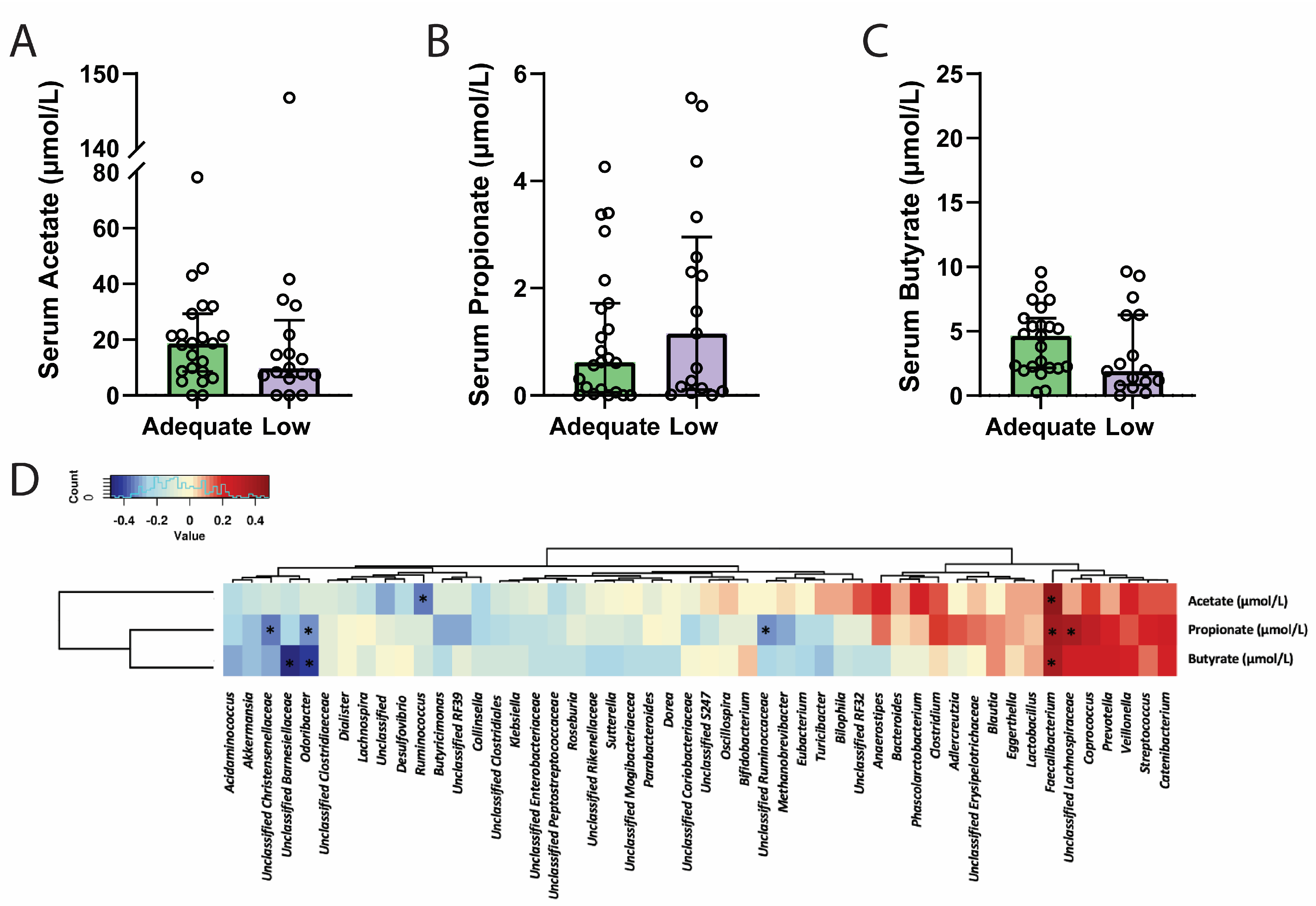
3.5. Dietary Fibre and Circulating Short-Chain Fatty Acid Levels
3.6. Circulating Short-Chain Fatty Acids and Genera Abundance
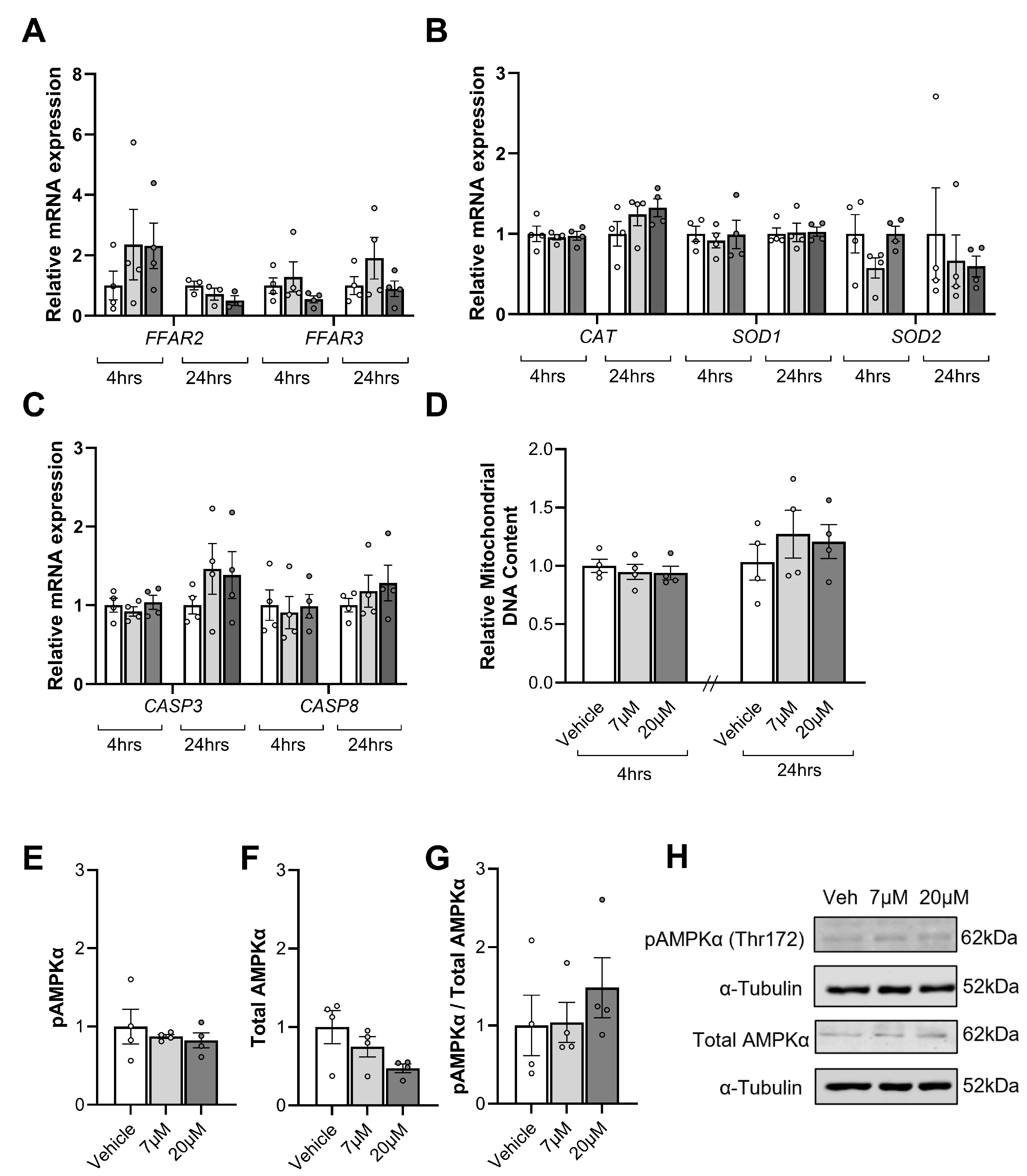
3.7. Impact of Short-Chain Fatty Acids on Placental Explants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopp, W. How Western Diet and Lifestyle Drive the Pandemic of Obesity and Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, S.; Solomon, C.G.; Hu, F.B. Dietary Fiber Intake, Dietary Glycemic Load, and the Risk for Gestational Diabetes Mellitus. Diabetes Care 2006, 29, 2223–2230. [Google Scholar] [CrossRef]
- Fall, C.H.D. Fetal malnutrition and long-term outcomes. Nestle Nutr. Inst. Workshop Ser. 2013, 74, 11–25. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The Health Benefits of Dietary Fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M. Connections Between the Gut Microbiome and Metabolic Hormones in Early Pregnancy in Overweight and Obese Women. Diabetes 2016, 65, 2214. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; Wilkinson, S.A.; Callaway, L.K.; McIntyre, H.D.; Morrison, M.; Dekker Nitert, M. Low dietary fiber intake increases Collinsella abundance in the gut microbiota of overweight and obese pregnant women. Gut Microbes 2018, 9, 189–201. [Google Scholar] [CrossRef]
- Barrett, H.L.; Gomez-Arango, L.F.; Wilkinson, S.A.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M. A Vegetarian Diet Is a Major Determinant of Gut Microbiota Composition in Early Pregnancy. Nutrients 2018, 10, 890. [Google Scholar] [CrossRef]
- O’Connor, H.; Li, S.; Hodge, A.; Callaway, L.; David Mclntyre, H.; Barrett, H.; Wilkinson, S.A.; Nitert, M.D. Gut microbiome composition is similar between pregnant women with excess body fat with healthy and less healthy dietary intake patterns. J. Hum. Nutr. Diet. 2023, 36, 1425–1437. [Google Scholar] [CrossRef]
- Myhrstad, M.C.W.; Tunsjø, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation—Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Chen, S.J.; Chen, C.C.; Liao, H.Y.; Lin, Y.T.; Wu, Y.W.; Liou, J.M.; Wu, M.S.; Kuo, C.H.; Lin, C.H. Association of Fecal and Plasma Levels of Short-Chain Fatty Acids with Gut Microbiota and Clinical Severity in Patients with Parkinson Disease. Neurology 2022, 98, e848–e858. [Google Scholar] [CrossRef]
- Fillier, T.A.; Shah, S.; Doody, K.M.; Pham, T.H.; Aubry, I.; Tremblay, M.L.; Cheema, S.K.; Blundell, J.; Thomas, R.H. Brief exposure of neuronal cells to levels of SCFAs observed in human systemic circulation impair lipid metabolism resulting in apoptosis. Sci. Rep. 2022, 12, 14355. [Google Scholar] [CrossRef]
- Kumagai, A.; Itakura, A.; Koya, D.; Kanasaki, K. AMP-Activated Protein (AMPK) in Pathophysiology of Pregnancy Complications. Int. J. Mol. Sci. 2018, 19, 3076. [Google Scholar] [CrossRef] [PubMed]
- Carey, E.A.K.; Albers, R.E.; Doliboa, S.R.; Hughes, M.; Wyatt, C.N.; Natale, D.R.C.; Brown, T.L. AMPK Knockdown in Placental Trophoblast Cells Results in Altered Morphology and Function. Stem Cells Dev. 2014, 23, 2921–2930. [Google Scholar] [CrossRef] [PubMed]
- Holland, O.J.; Cuffe, J.S.M.; Dekker Nitert, M.; Callaway, L.; Kwan Cheung, K.A.; Radenkovic, F.; Perkins, A.V. Placental mitochondrial adaptations in preeclampsia associated with progression to term delivery. Cell Death Dis. 2018, 9, 1150. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Y.; Qin, S.; Yang, H. Composition of Maternal Circulating Short-Chain Fatty Acids in Gestational Diabetes Mellitus and Their Associations with Placental Metabolism. Nutrients 2022, 14, 3727. [Google Scholar] [CrossRef] [PubMed]
- Nitert, M.D.; Barrett, H.L.; Foxcroft, K.; Tremellen, A.; Wilkinson, S.; Lingwood, B.; Tobin, J.M.; McSweeney, C.; O’Rourke, P.; McIntyre, H.D.; et al. SPRING: An RCT study of probiotics in the prevention of gestational diabetes mellitus in overweight and obese women. BMC Pregnancy Childbirth 2013, 13, 50. [Google Scholar] [CrossRef]
- Robinson, H.; Barrett, H.; Gomez-Arango, L.; McIntyre, H.D.; Callaway, L.; Dekker Nitert, M. Ketonuria Is Associated with Changes to the Abundance of Roseburia in the Gut Microbiota of Overweight and Obese Women at 16 Weeks Gestation: A Cross-Sectional Observational Study. Nutrients 2019, 11, 1836. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Tong, J.; Liu, C.; Summanen, P.; Xu, H.; Finegold, S.M. Application of quantitative real-time PCR for rapid identification of Bacteroides fragilis group and related organisms in human wound samples. Anaerobe 2011, 17, 64–68. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.-L.; Rainteau, D.; Bridonneau, C.; da Costa, G.; van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Fitzgerald, C.B.; Shkoporov, A.N.; Sutton, T.D.S.; Chaplin, A.V.; Velayudhan, V.; Ross, R.P.; Hill, C. Comparative analysis of Faecalibacterium prausnitzii genomes shows a high level of genome plasticity and warrants separation into new species-level taxa. BMC Genom. 2018, 19, 931. [Google Scholar] [CrossRef]
- Williams, B.L.; Hornig, M.; Parekh, T.; Lipkin, W.I. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. mBio 2012, 3, e00261-11. [Google Scholar] [CrossRef]
- Altemani, F.; Barrett, H.L.; Gomez-Arango, L.; Josh, P.; David McIntyre, H.; Callaway, L.K.; Morrison, M.; Tyson, G.W.; Dekker Nitert, M. Pregnant women who develop preeclampsia have lower abundance of the butyrate-producer Coprococcus in their gut microbiota. Pregnancy Hypertens. 2021, 23, 211–219. [Google Scholar] [CrossRef]
- Burton, G.J.; Sebire, N.J.; Myatt, L.; Tannetta, D.; Wang, Y.L.; Sadovsky, Y.; Staff, A.C.; Redman, C.W. Optimising sample collection for placental research. Placenta 2014, 35, 9–22. [Google Scholar] [CrossRef]
- Moylan, H.E.C.; Nguyen-Ngo, C.; Lim, R.; Lappas, M. The short-chain fatty acids butyrate and propionate protect against inflammation-induced activation of mediators involved in active labor: Implications for preterm birth. Mol. Hum. Reprod. 2020, 26, 452–468. [Google Scholar] [CrossRef]
- Wang, G.Y.; Qin, S.L.; Zheng, Y.N.; Geng, H.J.; Chen, L.; Yao, J.H.; Deng, L. Propionate promotes gluconeogenesis by regulating mechanistic target of rapamycin (mTOR) pathway in calf hepatocytes. Anim. Nutr. 2023, 15, 88–98. [Google Scholar] [CrossRef]
- Yoshida, H.; Ishii, M.; Akagawa, M. Propionate suppresses hepatic gluconeogenesis via GPR43/AMPK signaling pathway. Arch. Biochem. Biophys. 2019, 672, 108057. [Google Scholar] [CrossRef]
- Kent, N.L.; Atluri, S.C.; Moritz, K.M.; Cuffe, J.S.M. Maternal hypothyroidism in rats impairs placental nutrient transporter expression, increases labyrinth zone size, and impairs fetal growth. Placenta 2023, 139, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed]
- National Health and Medical Research Council. Australian Government Department of Health. In Nutrient Reference Values for Australia and New Zealand Including Recommended Dietary Intakes; Commonwealth of Australia: Canberra, Australia, 2017. [Google Scholar]
- Zhang, F.; Fan, D.; Huang, J.-l.; Zuo, T. The gut microbiome: Linking dietary fiber to inflammatory diseases. Med. Microecol. 2022, 14, 100070. [Google Scholar] [CrossRef]
- Roytio, H.; Mokkala, K.; Vahlberg, T.; Laitinen, K. Dietary intake of fat and fibre according to reference values relates to higher gut microbiota richness in overweight pregnant women. Br. J. Nutr. 2017, 118, 343–352. [Google Scholar] [CrossRef]
- Lin, A.; Bik, E.M.; Costello, E.K.; Dethlefsen, L.; Haque, R.; Relman, D.A.; Singh, U. Distinct Distal Gut Microbiome Diversity and Composition in Healthy Children from Bangladesh and the United States. PLoS ONE 2013, 8, e53838. [Google Scholar] [CrossRef]
- Ruengsomwong, S.; La-Ongkham, O.; Jiang, J.; Wannissorn, B.; Nakayama, J.; Nitisinprasert, S. Microbial Community of Healthy Thai Vegetarians and Non-Vegetarians, Their Core Gut Microbiota, and Pathogen Risk. J. Microbiol. Biotechnol. 2016, 26, 1723–1735. [Google Scholar] [CrossRef]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Breuninger, T.A.; Wawro, N.; Breuninger, J.; Reitmeier, S.; Clavel, T.; Six-Merker, J.; Pestoni, G.; Rohrmann, S.; Rathmann, W.; Peters, A.; et al. Associations between habitual diet, metabolic disease, and the gut microbiota using latent Dirichlet allocation. Microbiome 2021, 9, 61. [Google Scholar] [CrossRef]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhong, M.; Shen, Q.; Wu, Y.; Cao, M.; Ju, S.; Chen, L. Gut microbiota in early pregnancy among women with Hyperglycaemia vs. Normal blood glucose. BMC Pregnancy Childbirth 2020, 20, 284. [Google Scholar] [CrossRef]
- Ferrocino, I.; Ponzo, V.; Gambino, R.; Zarovska, A.; Leone, F.; Monzeglio, C.; Goitre, I.; Rosato, R.; Romano, A.; Grassi, G.; et al. Changes in the gut microbiota composition during pregnancy in patients with gestational diabetes mellitus (GDM). Sci. Rep. 2018, 8, 12216. [Google Scholar] [CrossRef]
- Hwang, N.; Eom, T.; Gupta, S.K.; Jeong, S.-Y.; Jeong, D.-Y.; Kim, Y.S.; Lee, J.-H.; Sadowsky, M.J.; Unno, T. Genes and Gut Bacteria Involved in Luminal Butyrate Reduction Caused by Diet and Loperamide. Genes 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Wiggins, H.S.; Jenkins, D.J.; Houston, H.; Jivraj, T.; Drasar, B.S.; Hill, M.J. Influence of diets high and low in animal fat on bowel habit, gastrointestinal transit time, fecal microflora, bile acid, and fat excretion. J. Clin. Investig. 1978, 61, 953–963. [Google Scholar] [CrossRef]
- Shrestha, N.; Sleep, S.L.; Cuffe, J.S.M.; Holland, O.J.; McAinch, A.J.; Dekker Nitert, M.; Hryciw, D.H. Pregnancy and diet-related changes in the maternal gut microbiota following exposure to an elevated linoleic acid diet. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E276–E285. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Rios-Covian, D.; Huillet, E.; Auger, S.; Khazaal, S.; Bermudez-Humaran, L.G.; Sokol, H.; Chatel, J.M.; Langella, P. Faecalibacterium: A bacterial genus with promising human health applications. FEMS Microbiol. Rev. 2023, 47, fuad039. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Yan, Y.; Ji, F.; Zhou, H. Maternal Obesity Increases Oxidative Stress in Placenta and It Is Associated with Intestinal Microbiota. Front. Cell Infect. Microbiol. 2021, 11, 671347. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Y.Y.; Li, S.P.; Zhao, H.M.; Jiang, F.J.; Wu, Y.X.; Tong, Y.; Pang, Q.F. Maternal propionate supplementation ameliorates glucose and lipid metabolic disturbance in hypoxia-induced fetal growth restriction. Food Funct. 2022, 13, 10724–10736. [Google Scholar] [CrossRef]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1897212. [Google Scholar] [CrossRef]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary short-chain fatty acid intake improves the hepatic metabolic condition via FFAR3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef]
- Wang, D.; Liu, C.D.; Tian, M.L.; Tan, C.Q.; Shu, G.; Jiang, Q.Y.; Zhang, L.; Yin, Y. Propionate promotes intestinal lipolysis and metabolic benefits via AMPK/LSD1 pathway in mice. J. Endocrinol. 2019, 243, 187–197. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, M.; Loor, J.J.; Jiang, Q.; Zhu, Y.; Li, W.; Du, X.; Song, Y.; Gao, W.; Lei, L.; et al. Propionate alleviates fatty acid-induced mitochondrial dysfunction, oxidative stress, and apoptosis by upregulating PPARG coactivator 1 alpha in hepatocytes. J. Dairy Sci. 2022, 105, 4581–4592. [Google Scholar] [CrossRef]
- Gesper, M.; Nonnast, A.B.H.; Kumowski, N.; Stoehr, R.; Schuett, K.; Marx, N.; Kappel, B.A. Gut-Derived Metabolite Indole-3-Propionic Acid Modulates Mitochondrial Function in Cardiomyocytes and Alters Cardiac Function. Front. Med. 2021, 8, 648259. [Google Scholar] [CrossRef]
- Wang, Y.; Christopher, B.A.; Wilson, K.A.; Muoio, D.; McGarrah, R.W.; Brunengraber, H.; Zhang, G.F. Propionate-induced changes in cardiac metabolism, notably CoA trapping, are not altered by l-carnitine. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E622–E633. [Google Scholar] [CrossRef] [PubMed]
- Corte-Iglesias, V.; Saiz, M.L.; Andrade-Lopez, A.C.; Salazar, N.; Bernet, C.R.; Martin-Martin, C.; Borra, J.M.; Lozano, J.J.; Aransay, A.M.; Diaz-Corte, C.; et al. Propionate and butyrate counteract renal damage and progression to chronic kidney disease. Nephrol. Dial. Transpl. 2024, 40, 133–150. [Google Scholar] [CrossRef]
- Jin, J.; Gao, L.; Zou, X.; Zhang, Y.; Zheng, Z.; Zhang, X.; Li, J.; Tian, Z.; Wang, X.; Gu, J.; et al. Gut Dysbiosis Promotes Preeclampsia by Regulating Macrophages and Trophoblasts. Circ. Res. 2022, 131, 492–506. [Google Scholar] [CrossRef]
- Hebden, L.; Kostan, E.; O’Leary, F.; Hodge, A.; Allman-Farinelli, M. Validity and reproducibility of a food frequency questionnaire as a measure of recent dietary intake in young adults. PLoS ONE 2013, 8, e75156. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adequate Fibre | Low Fibre | p | ||
|---|---|---|---|---|
| Clinical characteristics | SPRING Treatment (Placebo%) # | 56% | 52% | ns |
| Ethnicity (Caucasian%) # | 84% | 85% | ns | |
| Baseline BMI (kg/m2) ^ | 30.86 (28.31–37.15) | 30.88 (29.36–25.67) | ns | |
| 28-week BMI (kg/m2) ^ | 33.76 (30.33–38.57) | 33.75 (31.79–36.73) | ns | |
| Age (years) | 33 (31–36) | 30 (27–34) | * | |
| Fasting Blood Glucose (mmol/L) | 4.20 (3.95–4.50) | 4.20 ± (3.90–4.50) | ns | |
| C-Peptide (nmol/L) ^ | 0.80 (0.65–0.90) | 0.80 (0.60–1.20) | ns | |
| Insulin (mU/L) ^ | 9.30 (6.45–12.00) | 8.60 (6.00–16.00) | ns | |
| Cholesterol (mmol/L) | 6.40 (5.45–7.25) | 6.50 (6.20–7.70) | ns | |
| Triglycerides (mmol/L) ^ | 1.90 (1.50–2.70) | 2.20 (1.60–2.60) | ns | |
| HDL (mmol/L) | 1.90 (1.70–2.30) | 1.80 (1.50–2.10) | ns | |
| LDL (mmol/L) | 3.60 (3.03–4.25) | 3.90 (3.30–5.00) | ns | |
| VLDL (mmol/L) | 0.90 (0.70–1.10) | 1.00 (0.70–1.20) | ns | |
| Pregnancy | Gestational Age (weeks) | 40.36 (39.00–41.22) | 40.22 (38.57–41.18) | ns |
| Outcomes | Offspring Sex (Male%) # | 48% | 44% | ns |
| Birthweight (g) | 3606 (3413–3977) | 3833 (3437–4155) | ns | |
| Delivery (Vaginal%) # | 80% | 66% | ns |
| Dietary | Energy Intake (kJ/day)^ | 5872 (4998–7350) | 6102 (4586–6749) | ns |
|---|---|---|---|---|
| Data | Energy-adjusted fibre intake (mg/kJ) | 3.83 (3.30–4.12) | 2.09 (1.80–2.27) | **** |
| Gross fibre intake (g/day) ^ | 21.84 (16.86–30.05) | 11.82 (9.58–15.46) | **** | |
| Energy-adjusted carbohydrate intake (mg/kJ) | 25.80 (24.57–27.83) | 23.71 (20.99–24.84) | *** | |
| Gross carbohydrate intake (g/day) ^ | 164.3 (132.2–205.3) | 134.9 (108.7–167.6) | * | |
| Energy-adjusted fat intake (mg/kJ) | 13.20 (11.75–15.65) | 11.50 (10.40–13.20) | *** | |
| Gross fat intake (g/day) ^ | 62.82 (51.56–74.18) | 66.88 (51.97–73.75) | ns | |
| Energy-adjusted protein intake (mg/kJ) | 11.33 (10.12–12.45) | 11.93 (10.88–13.16) | ns | |
| Gross protein intake (g/day) ^ | 67.48 (54.48–83.52) | 72.80 (48.82–94.40) | ns | |
| Energy-adjusted starch intake (mg/kJ) | 13.15 (11.77–15.69) | 11.48 (10.40–13.24) | *** | |
| Gross starch intake (g/day) ^ | 80.01 (68.59–107.9) | 68.08 (51.32–80.18) | * | |
| Energy-adjusted sugar intake (mg/kJ) ^ | 11.77 (10.24–13.51) | 11.03 (8.22–13.13) | ns | |
| Gross sugar intake (g/day) | 81.21 (58.16–96.81) | 62.15 (44.54–89.16) | ns | |
| Energy-adjusted saturated fat intake (mg/kJ) | 4.15 (3.61–4.75) | 5.23 (4.77–5.59) | **** | |
| Gross saturated fat intake (g/day)^ | 26.34 (20.52–31.51) | 29.87 (24.72–33.87) | ns | |
| Energy-adjusted monounsaturated fat intake (mg/kJ) ^ | 3.51 (3.35–3.89) | 3.79 (3.60–4.09) | * | |
| Gross monosaturated fat intake (g/day) ^ | 22.23 (18.04–26.56) | 23.23 (16.94–26.68) | ns | |
| Energy-adjusted polyunsaturated fat intake (mg/kJ) ^ | 1.30 (1.15–1.75) | 1.20 (1.00–1.300) | * | |
| Gross polyunsaturated fat intake (g/day) ^ | 9.06 (6.46–12.09) | 6.59 (5.07–7.67) | ** | |
| Energy from fat (%) | 38.58 (36.34–40.84) | 42.75 (39.51–44.90) | ** | |
| Energy from carbohydrates (%) | 43.82 (41.03–46.46) | 39.59 (35.05–41.48) | **** | |
| Energy from protein (%) | 18.92 (16.89–20.79) | 20.09 ± 0.64% | ns |
| Macronutrient Intake (mg/kJ or g/Day) | Acetate | Propionate | Butyrate | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| Fibre (mg/kJ) | 0.15 | 0.13 | –0.08 | 0.76 | 0.17 | 0.28 |
| Fibre (g/day) | 0.09 | 0.55 | –0.04 | 0.80 | 0.24 | 0.13 |
| Carbohydrates (mg/kJ) | 0.01 | 0.94 | 0.14 | 0.37 | 0.35 | 0.03 |
| Starch (mg/kJ) | 0.04 | 0.81 | 0.09 | 0.57 | 0.26 | 0.10 |
| Sugars (mg/kJ) | 0.02 | 0.86 | 0.08 | 0.61 | 0.18 | 0.27 |
| Fat (mg/kJ) | 0.04 | 0.61 | 0.09 | 0.57 | 0.26 | 0.10 |
| Saturated fat (mg/kJ) | –0.08 | 0.61 | –0.15 | 0.33 | –0.38 | 0.01 |
| Monounsaturated fat (mg/kJ) | –0.24 | 0.14 | –0.26 | 0.10 | –0.32 | 0.04 |
| Polyunsaturated fat (mg/kJ) | 0.01 | 0.93 | 0.13 | 0.41 | 0.26 | 0.09 |
| Protein (mg/kJ) | 0.10 | 0.51 | –0.10 | 0.53 | –0.15 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanderpeet, C.L.; Dorey, E.S.; Neal, E.S.; Mullins, T.; McIntyre, D.H.; Callaway, L.K.; Barrett, H.L.; Dekker Nitert, M.; Cuffe, J.S.M. Dietary Fibre Modulates Gut Microbiota in Late Pregnancy Without Altering SCFA Levels, and Propionate Treatement Has No Effect on Placental Explant Function. Nutrients 2025, 17, 1234. https://doi.org/10.3390/nu17071234
Vanderpeet CL, Dorey ES, Neal ES, Mullins T, McIntyre DH, Callaway LK, Barrett HL, Dekker Nitert M, Cuffe JSM. Dietary Fibre Modulates Gut Microbiota in Late Pregnancy Without Altering SCFA Levels, and Propionate Treatement Has No Effect on Placental Explant Function. Nutrients. 2025; 17(7):1234. https://doi.org/10.3390/nu17071234
Chicago/Turabian StyleVanderpeet, Chelsea L., Emily S. Dorey, Elliott S. Neal, Thomas Mullins, David H. McIntyre, Leonie K. Callaway, Helen L. Barrett, Marloes Dekker Nitert, and James S. M. Cuffe. 2025. "Dietary Fibre Modulates Gut Microbiota in Late Pregnancy Without Altering SCFA Levels, and Propionate Treatement Has No Effect on Placental Explant Function" Nutrients 17, no. 7: 1234. https://doi.org/10.3390/nu17071234
APA StyleVanderpeet, C. L., Dorey, E. S., Neal, E. S., Mullins, T., McIntyre, D. H., Callaway, L. K., Barrett, H. L., Dekker Nitert, M., & Cuffe, J. S. M. (2025). Dietary Fibre Modulates Gut Microbiota in Late Pregnancy Without Altering SCFA Levels, and Propionate Treatement Has No Effect on Placental Explant Function. Nutrients, 17(7), 1234. https://doi.org/10.3390/nu17071234