
Low Dietary Folate Increases Developmental Delays in the Litters of Mthfr677TT Mice

 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Diets
2.2. Metabolite Measurement
2.3. Western Blots
2.4. Statistics
3. Results
3.1. Plasma MethylTHF Decreases and Homocysteine Increases Due to TT Genotype and Low Folate Diet
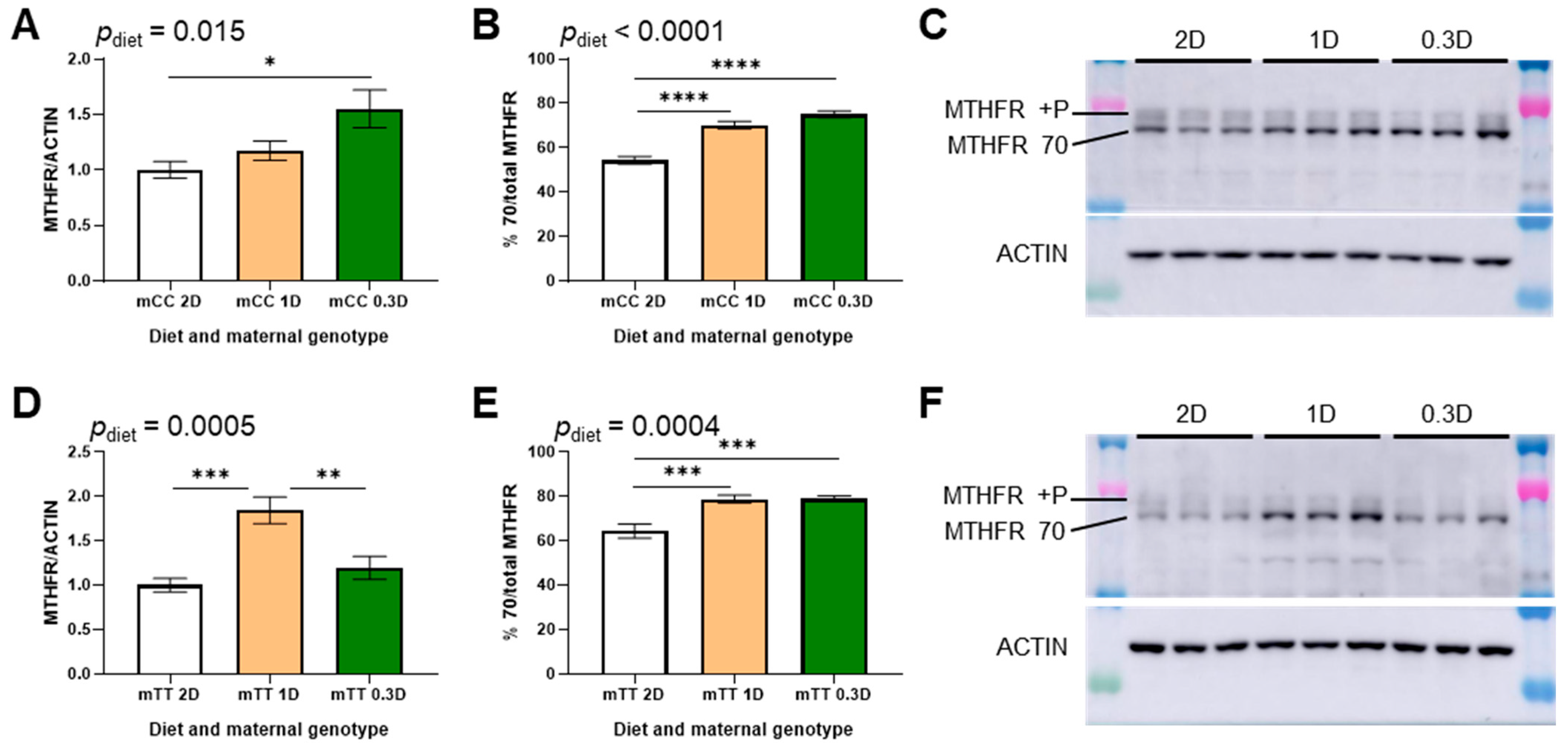
3.2. Effects of Low Dietary Folate on Hepatic MTHFR Expression Differ Between CC and TT Mothers
3.3. Low Dietary Folate Increased Developmental Delays in the Litters of TT Mothers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2D | 2 mg folic acid/kg diet |
| 1D | 1 mg folic acid/kg diet |
| 0.3D | 0.3 mg folic acid/kg diet |
| CC | Mthfr677CC |
| CT | Mthfr677CT |
| e | embryonic |
| E | embryonic day |
| Hcy | homocysteine |
| m | maternal |
| MTHFR | methylenetetrahydrofolate reductase |
| methylTHF | methyltetrahydrofolate |
| SAM | S-adenosylmethionine |
| tHcy | total homocysteine |
| TT | Mthfr677TT |
References
- Bailey, L.B.; Stover, P.J.; McNulty, H.; Fenech, M.F.; Gregory, J.F.; Mills, J.L.; Pfeiffer, C.M.; Fazili, Z.; Zhang, M.; Ueland, P.M.; et al. Biomarkers of Nutrition for Development—Folate Review. J. Nutr. 2015, 145, 1636S–1680S. [Google Scholar] [CrossRef]
- Crider, K.S.; Qi, Y.P.; Yeung, L.F.; Mai, C.T.; Head Zauche, L.; Wang, A.; Daniels, K.; Williams, J.L. Folic Acid and the Prevention of Birth Defects: 30 Years of Opportunity and Controversies. Annu. Rev. Nutr. 2022, 42, 423–452. [Google Scholar] [CrossRef]
- Graafland, N.; Rousian, M.; de Zwart, M.L.; Steegers-Theunissen, R.P.M.; Steegers, E.A.P.; Posthumus, A.G. Parental conditions, modifiable lifestyle factors, and first trimester growth and development: A systematic review. Hum. Reprod. Update 2025, 31, 166–182. [Google Scholar] [CrossRef]
- Zhou, Y.; Crider, K.S.; Yeung, L.F.; Rose, C.E.; Li, Z.; Berry, R.J.; Li, S.; Moore, C.A. Periconceptional folic acid use prevents both rare and common neural tube defects in China. Birth Defects Res. 2022, 114, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Lin, S.; Zhuang, J.; Bloom, M.S.; Smith, M.; Nie, Z.; Mai, J.; Ou, Y.; Wu, Y.; Gao, X.; et al. First-Trimester Maternal Folic Acid Supplementation Reduced Risks of Severe and Most Congenital Heart Diseases in Offspring: A Large Case-Control Study. J. Am. Heart Assoc. 2020, 9, e015652. [Google Scholar] [CrossRef]
- Yang, L.; Wang, W.; Mao, B.; Qiu, J.; Guo, H.; Yi, B.; He, X.; Lin, X.; Lv, L.; Xu, X.; et al. Maternal Folic Acid Supplementation, Dietary Folate Intake, and Low Birth Weight: A Birth Cohort Study. Front. Public Health 2022, 10, 844150. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.; Lopes, C.; Pinto, E. Folate and folic acid in the periconceptional period: Recommendations from official health organizations in thirty-six countries worldwide and WHO. Public Health Nutr. 2016, 19, 176–189. [Google Scholar] [CrossRef]
- Barry, M.J.; Nicholson, W.K.; Silverstein, M.; Chelmow, D.; Coker, T.R.; Davis, E.M.; Donahue, K.E.; Jaén, C.R.; Li, L.; Ogedegbe, G.; et al. Folic Acid Supplementation to Prevent Neural Tube Defects: US Preventive Services Task Force Reaffirmation Recommendation Statement. JAMA 2023, 330, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, A.; Yeung, L.F.; Qi, Y.P.; Pfeiffer, C.M.; Crider, K.S. Folate and vitamin B12 usual intake and biomarker status by intake source in United States adults aged ≥19 y: NHANES 2007–2018. Am. J. Clin. Nutr. 2023, 118, 241–254. [Google Scholar] [CrossRef]
- Cui, M.; Lu, X.L.; Lyu, Y.Y.; Wang, F.; Xie, X.L.; Cheng, X.Y.; Zhang, T. Knowledge and intake of folic acid to prevent neural tube defects among pregnant women in urban China: A cross-sectional study. BMC Pregnancy Childbirth 2021, 21, 432. [Google Scholar] [CrossRef]
- Iglesias-Vázquez, L.; Serrat, N.; Bedmar, C.; Pallejà-Millán, M.; Arija, V. Prenatal folic acid supplementation and folate status in early pregnancy: ECLIPSES study. Br. J. Nutr. 2022, 128, 1938–1945. [Google Scholar] [CrossRef]
- Camier, A.; Kadawathagedara, M.; Lioret, S.; Bois, C.; Cheminat, M.; Dufourg, M.N.; Charles, M.A.; de Lauzon-Guillain, B. Social Inequalities in Prenatal Folic Acid Supplementation: Results from the ELFE Cohort. Nutrients 2019, 11, 1108. [Google Scholar] [CrossRef]
- Wegner, C.; Kancherla, V.; Lux, A.; Köhn, A.; Bretschneider, D.; Freese, K.; Heiduk, M.; Redlich, A.; Schleef, D.; Jorch, G.; et al. Periconceptional folic acid supplement use among women of reproductive age and its determinants in central rural Germany: Results from a cross sectional study. Birth Defects Res. 2020, 112, 1057–1066. [Google Scholar] [CrossRef]
- Bhide, P.; Kar, A. Prevalence and determinants of folate deficiency among urban Indian women in the periconception period. Eur. J. Clin. Nutr. 2019, 73, 1639–1641. [Google Scholar] [CrossRef]
- Aweke, M.N.; Fentie, E.A.; Agimas, M.C.; Baffa, L.D.; Shewarega, E.S.; Belew, A.K.; Muhammad, E.A.; Mengistu, B. Folic acid supplementation during preconception period in sub-Saharan African countries: A systematic review and meta-analysis. PLoS ONE 2025, 20, e0318422. [Google Scholar] [CrossRef] [PubMed]
- Frosst, P.; Blom, H.J.; Milos, R.; Goyette, P.; Sheppard, C.A.; Matthews, R.G.; Boers, G.J.; den Heijer, M.; Kluijtmans, L.A.; van den Heuvel, L.P.; et al. A candidate genetic risk factor for vascular disease: A common mutation in methylenetetrahydrofolate reductase. Nat. Genet. 1995, 10, 111–113. [Google Scholar] [CrossRef]
- Tabatabaei, R.S.; Fatahi-Meibodi, N.; Meibodi, B.; Javaheri, A.; Abbasi, H.; Hadadan, A.; Bahrami, R.; Mirjalili, S.R.; Karimi-Zarchi, M.; Neamatzadeh, H. Association of Fetal MTHFR C677T Polymorphism with Susceptibility to Neural Tube Defects: A Systematic Review and Update Meta-Analysis. Fetal Pediatr. Pathol. 2022, 41, 225–241. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, J.; Wang, B.; Ding, C.; Liu, H. Association between MTHFR C677T polymorphism and neural tube defect risks: A comprehensive evaluation in three groups of NTD patients, mothers, and fathers. Birth Defects Res. A Clin. Mol. Teratol. 2015, 103, 488–500. [Google Scholar] [CrossRef]
- Chen, H.; Chen, X.; Yao, Q.; Xin, J.; Zhang, Y.; Huang, X.; Wang, D.; Li, M.; Zhang, T.; Tillmann, T.; et al. Appraising the causal relevance of maternal red blood cell folate and congenital heart disease in offspring: 2-sample Mendelian randomization. Ann. Epidemiol. 2025, 106, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Vafapour, M.; Talebi, H.; Danaei, M.; Yeganegi, M.; Azizi, S.; Dastgheib, S.A.; Bahrami, R.; Pourkazemi, M.; Jayervand, F.; Shahbazi, A.; et al. Global and population-specific association of MTHFR polymorphisms with preterm birth risk: A consolidated analysis of 44 studies. BMC Pregnancy Childbirth 2025, 25, 230. [Google Scholar] [CrossRef] [PubMed]
- Guenther, B.D.; Sheppard, C.A.; Tran, P.; Rozen, R.; Matthews, R.G.; Ludwig, M.L. The structure and properties of methylenetetrahydrofolate reductase from Escherichia coli suggest how folate ameliorates human hyperhomocysteinemia. Nat. Struct. Biol. 1999, 6, 359–365. [Google Scholar]
- Fredriksen, A.; Meyer, K.; Ueland, P.M.; Vollset, S.E.; Grotmol, T.; Schneede, J. Large-scale population-based metabolic phenotyping of thirteen genetic polymorphisms related to one-carbon metabolism. Hum. Mutat. 2007, 28, 856–865. [Google Scholar] [CrossRef]
- Yang, Q.H.; Botto, L.D.; Gallagher, M.; Friedman, J.M.; Sanders, C.L.; Koontz, D.; Nikolova, S.; Erickson, J.D.; Steinberg, K. Prevalence and effects of gene-gene and gene-nutrient interactions on serum folate and serum total homocysteine concentrations in the United States: Findings from the third National Health and Nutrition Examination Survey DNA Bank. Am. J. Clin. Nutr. 2008, 88, 232–246. [Google Scholar] [CrossRef]
- Colson, N.J.; Naug, H.L.; Nikbakht, E.; Zhang, P.; McCormack, J. The impact of MTHFR 677 C/T genotypes on folate status markers: A meta-analysis of folic acid intervention studies. Eur. J. Nutr. 2017, 56, 247–260. [Google Scholar] [CrossRef]
- Tsang, B.L.; Devine, O.J.; Cordero, A.M.; Marchetta, C.M.; Mulinare, J.; Mersereau, P.; Guo, J.; Qi, Y.P.; Berry, R.J.; Rosenthal, J.; et al. Assessing the association between the methylenetetrahydrofolate reductase (MTHFR) 677C>T polymorphism and blood folate concentrations: A systematic review and meta-analysis of trials and observational studies. Am. J. Clin. Nutr. 2015, 101, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.E.; Faquette, M.L.; Leclerc, D.; Keser, V.; Luan, Y.; Bennett-Firmin, J.L.; Malysheva, O.V.; Reagan, A.M.; Howell, G.R.; Caudill, M.A.; et al. Folic Acid and Methyltetrahydrofolate Supplementation in the Mthfr(677C>T) Mouse Model with Hepatic Steatosis. Nutrients 2024, 17, 82. [Google Scholar] [CrossRef] [PubMed]
- Ulvik, A.; Ueland, P.M.; Fredriksen, A.; Meyer, K.; Vollset, S.E.; Hoff, G.; Schneede, J. Functional inference of the methylenetetrahydrofolate reductase 677C > T and 1298A > C polymorphisms from a large-scale epidemiological study. Hum. Genet. 2007, 121, 57–64. [Google Scholar] [CrossRef] [PubMed]
- van Beynum, I.M.; Kapusta, L.; den Heijer, M.; Vermeulen, S.H.; Kouwenberg, M.; Daniels, O.; Blom, H.J. Maternal MTHFR 677C>T is a risk factor for congenital heart defects: Effect modification by periconceptional folate supplementation. Eur. Heart J. 2006, 27, 981–987. [Google Scholar] [CrossRef]
- Trimmer, E.E. Methylenetetrahydrofolate reductase: Biochemical characterization and medical significance. Curr. Pharm. Des. 2013, 19, 2574–2593. [Google Scholar] [CrossRef]
- Yamada, K.; Strahler, J.R.; Andrews, P.C.; Matthews, R.G. Regulation of human methylenetetrahydrofolate reductase by phosphorylation. Proc. Natl. Acad. Sci. USA 2005, 102, 10454–10459. [Google Scholar] [CrossRef]
- Christensen, K.E.; Bahous, R.H.; Hou, W.; Deng, L.; Malysheva, O.V.; Arning, E.; Bottiglieri, T.; Caudill, M.A.; Jerome-Majewska, L.A.; Rozen, R. Low Dietary Folate Interacts with MTHFD1 Synthetase Deficiency in Mice, a Model for the R653Q Variant, to Increase Incidence of Developmental Delays and Defects. J. Nutr. 2018, 148, 501–509. [Google Scholar] [CrossRef]
- Froese, D.S.; Kopec, J.; Rembeza, E.; Bezerra, G.A.; Oberholzer, A.E.; Suormala, T.; Lutz, S.; Chalk, R.; Borkowska, O.; Baumgartner, M.R.; et al. Structural basis for the regulation of human 5,10-methylenetetrahydrofolate reductase by phosphorylation and S-adenosylmethionine inhibition. Nat. Commun. 2018, 9, 2261. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ramsamooj, S.; Li, Q.; Johnson, J.L.; Yaron, T.M.; Sharra, K.; Cantley, L.C. Regulation of folate and methionine metabolism by multisite phosphorylation of human methylenetetrahydrofolate reductase. Sci. Rep. 2019, 9, 4190. [Google Scholar] [CrossRef]
- Yamada, K.; Mendoza, J.; Koutmos, M. Structural basis of S-adenosylmethionine-dependent allosteric transition from active to inactive states in methylenetetrahydrofolate reductase. Nat. Commun. 2024, 15, 5167. [Google Scholar] [CrossRef]
- Blomgren, L.K.M.; Huber, M.; Mackinnon, S.R.; Bürer, C.; Baslé, A.; Yue, W.W.; Froese, D.S.; McCorvie, T.J. Dynamic inter-domain transformations mediate the allosteric regulation of human 5, 10-methylenetetrahydrofolate reductase. Nat. Commun. 2024, 15, 3248. [Google Scholar] [CrossRef]
- Büchler, L.R.; Blomgren, L.K.M.; Bürer, C.; Zanotelli, V.R.T.; Froese, D.S. Evidence for interaction of 5,10-methylenetetrahydrofolate reductase (MTHFR) with methylenetetrahydrofolate dehydrogenase (MTHFD1) and general control nonderepressible 1 (GCN1). Biochimie 2025, 230, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Thakur, J.; Suyal, S.; Oniel, R.; Chakraborty, R.; Pradhan, S.; Sharma, M.; Sengupta, S.; Laxman, S.; Masakapalli, S.K.; et al. Allosteric inhibition of MTHFR prevents futile SAM cycling and maintains nucleotide pools in one-carbon metabolism. J. Biol. Chem. 2020, 295, 16037–16057. [Google Scholar] [CrossRef]
- Reagan, A.M.; Christensen, K.E.; Graham, L.C.; Bedwell, A.A.; Eldridge, K.; Speedy, R.; Figueiredo, L.L.; Persohn, S.C.; Bottiglieri, T.; Nho, K.; et al. The 677C > T variant in methylenetetrahydrofolate reductase causes morphological and functional cerebrovascular deficits in mice. J. Cereb. Blood Flow. Metab. 2022, 42, 2333–2350. [Google Scholar] [CrossRef]
- Leclerc, D.; Christensen, K.E.; Reagan, A.M.; Keser, V.; Luan, Y.; Malysheva, O.V.; Wasek, B.; Bottiglieri, T.; Caudill, M.A.; Howell, G.R.; et al. Folate Deficiency and/or the Genetic Variant Mthfr(677C >T) Can Drive Hepatic Fibrosis or Steatosis in Mice, in a Sex-Specific Manner. Mol. Nutr. Food Res. 2024, 68, e2300355. [Google Scholar] [CrossRef]
- Bieri, J.G.; Stoewsand, G.S.; Briggs, G.M.; Phillips, R.W.; Woodard, J.C.; Knapka, J.J. Report of the American Institute of Nutrition ad hoc Committee on Standards for Nutritional Studies. J. Nutr. 1977, 107, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of the Mouse. In Nutrient Requirements of Laboratory Animals: Fourth Revised Edition, 1995; National Academy Press: Washington, DC, USA, 1995; pp. 80–102. [Google Scholar]
- Munezero, E.; Behan, N.A.; Diaz, S.G.; Neumann, E.M.; MacFarlane, A.J. Poor Reporting Quality in Basic Nutrition Research: A Case Study Based on a Scoping Review of Recent Folate Research in Mouse Models (2009–2021). Adv. Nutr. 2022, 13, 2666–2678. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.E.; Malysheva, O.V.; Carlin, S.; Matias, F.; MacFarlane, A.J.; Jacobs, R.L.; Caudill, M.A.; Rozen, R. Mild Choline Deficiency and MTHFD1 Synthetase Deficiency Interact to Increase Incidence of Developmental Delays and Defects in Mice. Nutrients 2021, 14, 127. [Google Scholar] [CrossRef]
- McFarlane, L.; Truong, V.; Palmer, J.S.; Wilhelm, D. Novel PCR assay for determining the genetic sex of mice. Sex. Dev. 2013, 7, 207–211. [Google Scholar] [CrossRef]
- Tunster, S.J. Genetic sex determination of mice by simplex PCR. Biol. Sex. Differ. 2017, 8, 31. [Google Scholar] [CrossRef]
- Arning, E.; Bottiglieri, T. Quantitation of 5-Methyltetrahydrofolate in Cerebrospinal Fluid Using Liquid Chromatography-Electrospray Tandem Mass Spectrometry. Methods Mol. Biol. 2016, 1378, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Ducros, V.; Belva-Besnet, H.; Casetta, B.; Favier, A. A robust liquid chromatography tandem mass spectrometry method for total plasma homocysteine determination in clinical practice. Clin. Chem. Lab. Med. 2006, 44, 987–990. [Google Scholar] [CrossRef]
- Christensen, K.E.; Mikael, L.G.; Leung, K.Y.; Levesque, N.; Deng, L.; Wu, Q.; Malysheva, O.V.; Best, A.; Caudill, M.A.; Greene, N.D.; et al. High folic acid consumption leads to pseudo-MTHFR deficiency, altered lipid metabolism, and liver injury in mice. Am. J. Clin. Nutr. 2015, 101, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lenth, R.V. emmeans: Estimated Marginal Means, aka Least-Squares Means, R package version 1.11.1; R Foundation for Statistical Computing: Vienna, Austria, 2025. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 14 May 2025).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2025; Available online: https://www.R-project.org/ (accessed on 14 May 2025).
- RStudio Team. RStudio: Integrated Development for R.; RStudio, PBC.: Boston, MA, USA, 2020; Available online: https://www.rstudio.com/ (accessed on 14 May 2025).
- Pickell, L.; Li, D.; Brown, K.; Mikael, L.G.; Wang, X.L.; Wu, Q.; Luo, L.; Jerome-Majewska, L.; Rozen, R. Methylenetetrahydrofolate reductase deficiency and low dietary folate increase embryonic delay and placental abnormalities in mice. Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 531–541. [Google Scholar] [CrossRef]
- Kubo, Y.; Fukuoka, H.; Kawabata, T.; Shoji, K.; Mori, C.; Sakurai, K.; Nishikawa, M.; Ohkubo, T.; Oshida, K.; Yanagisawa, N.; et al. Distribution of 5-Methyltetrahydrofolate and Folic Acid Levels in Maternal and Cord Blood Serum: Longitudinal Evaluation of Japanese Pregnant Women. Nutrients 2020, 12, 1633. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Weir, D.G. The methyl folate trap. A physiological response in man to prevent methyl group deficiency in kwashiorkor (methionine deficiency) and an explanation for folic-acid induced exacerbation of subacute combined degeneration in pernicious anaemia. Lancet 1981, 2, 337–340. [Google Scholar] [CrossRef]
- Wu, H.; Zhu, P.; Geng, X.; Liu, Z.; Cui, L.; Gao, Z.; Jiang, B.; Yang, L. Genetic polymorphism of MTHFR C677T with preterm birth and low birth weight susceptibility: A meta-analysis. Arch. Gynecol. Obstet. 2017, 295, 1105–1118. [Google Scholar] [CrossRef]
- Greene, N.D.; Copp, A.J. Mouse models of neural tube defects: Investigating preventive mechanisms. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 135c, 31–41. [Google Scholar] [CrossRef]
- Heid, M.K.; Bills, N.D.; Hinrichs, S.H.; Clifford, A.J. Folate deficiency alone does not produce neural tube defects in mice. J. Nutr. 1992, 122, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Burgoon, J.M.; Selhub, J.; Nadeau, M.; Sadler, T.W. Investigation of the effects of folate deficiency on embryonic development through the establishment of a folate deficient mouse model. Teratology 2002, 65, 219–227. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Oktayani, P.P.I.; Lee, S.D.; Huang, L.C. Choline in pregnant women: A systematic review and meta-analysis. Nutr. Rev. 2025, 83, e273–e289. [Google Scholar] [CrossRef]
- McNulty, H.; Pentieva, K.; Ward, M. Causes and Clinical Sequelae of Riboflavin Deficiency. Annu. Rev. Nutr. 2023, 43, 101–122. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christensen, K.E.; Faquette, M.-L.; Keser, V.; Reagan, A.M.; Gebert, A.T.; Bottiglieri, T.; Howell, G.R.; Rozen, R. Low Dietary Folate Increases Developmental Delays in the Litters of Mthfr677TT Mice. Nutrients 2025, 17, 2536. https://doi.org/10.3390/nu17152536
Christensen KE, Faquette M-L, Keser V, Reagan AM, Gebert AT, Bottiglieri T, Howell GR, Rozen R. Low Dietary Folate Increases Developmental Delays in the Litters of Mthfr677TT Mice. Nutrients. 2025; 17(15):2536. https://doi.org/10.3390/nu17152536
Chicago/Turabian StyleChristensen, Karen E., Marie-Lou Faquette, Vafa Keser, Alaina M. Reagan, Aaron T. Gebert, Teodoro Bottiglieri, Gareth R. Howell, and Rima Rozen. 2025. "Low Dietary Folate Increases Developmental Delays in the Litters of Mthfr677TT Mice" Nutrients 17, no. 15: 2536. https://doi.org/10.3390/nu17152536
APA StyleChristensen, K. E., Faquette, M.-L., Keser, V., Reagan, A. M., Gebert, A. T., Bottiglieri, T., Howell, G. R., & Rozen, R. (2025). Low Dietary Folate Increases Developmental Delays in the Litters of Mthfr677TT Mice. Nutrients, 17(15), 2536. https://doi.org/10.3390/nu17152536