
Antioxidant and Anti-Inflammatory Activities of Latilactobacillus curvatus and L. sakei Isolated from Green Tripe

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
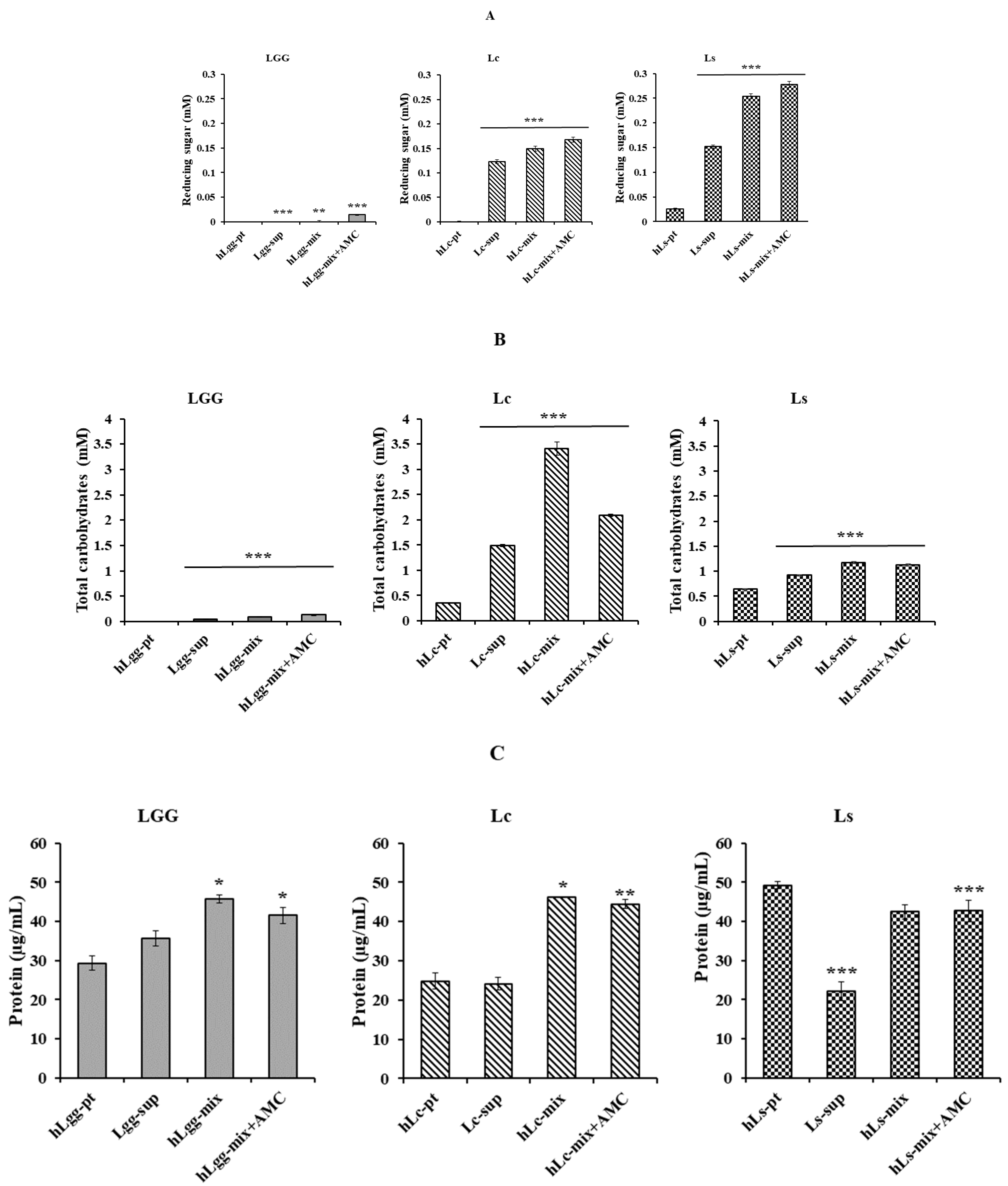
2.2. Biochemical Characterization: Determination of Reducing Sugar, Total Carbohydrate, and Protein Content
2.2.1. Preparation of Active Molecular Chitosan
2.2.2. Quantification of Reducing Sugar Content
2.2.3. Quantitative Analysis of Total Carbohydrates and Protein
2.3. Isolation of GRET-Derived Beneficial Bacteria and Separation of Intracellular Metabolites Through Heat Treatment
2.4. Antioxidant Activity
2.4.1. Ferric Reducing Antioxidant Power (FRAP) Assay
2.4.2. ABTS Radical Scavenging Activity
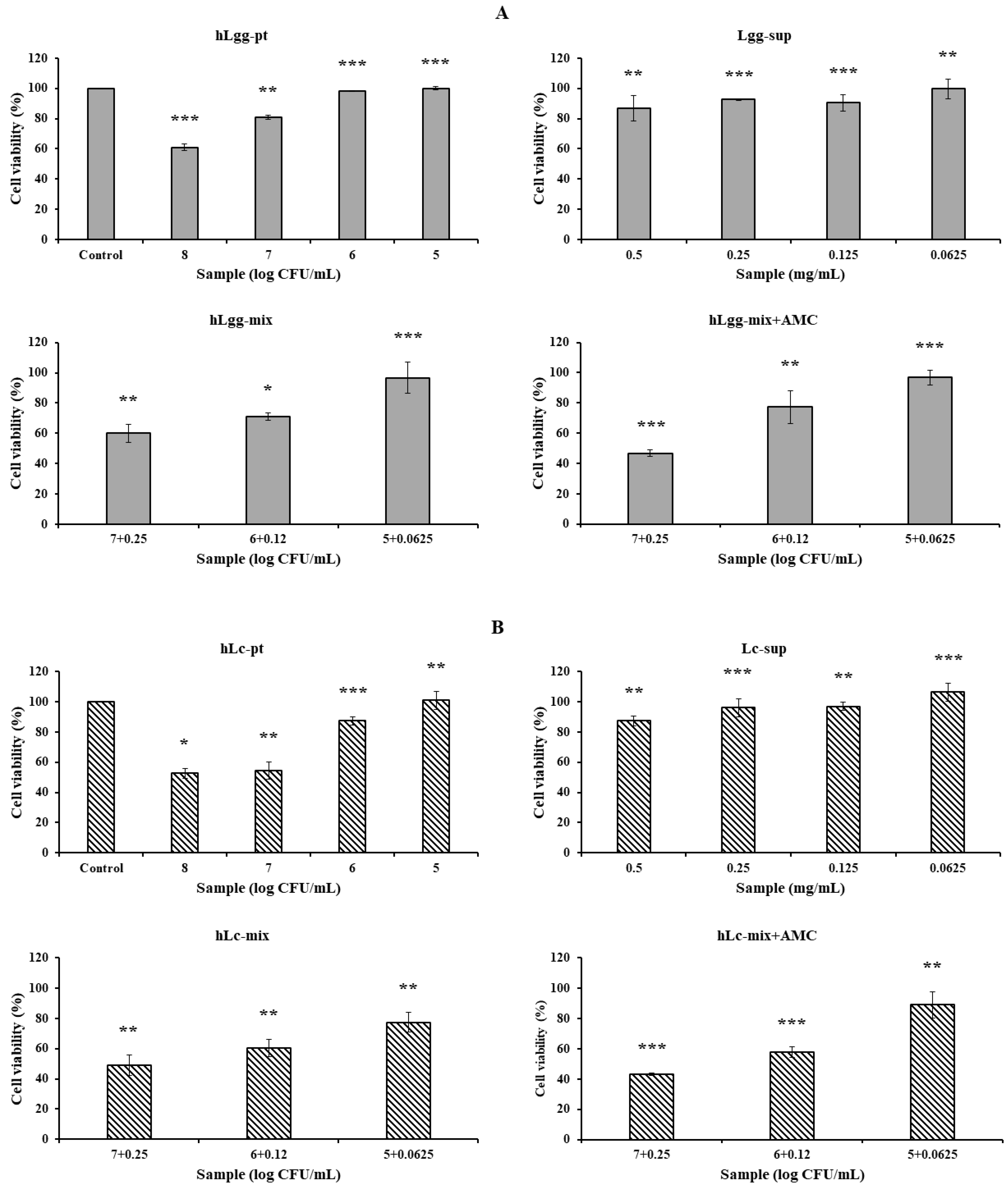
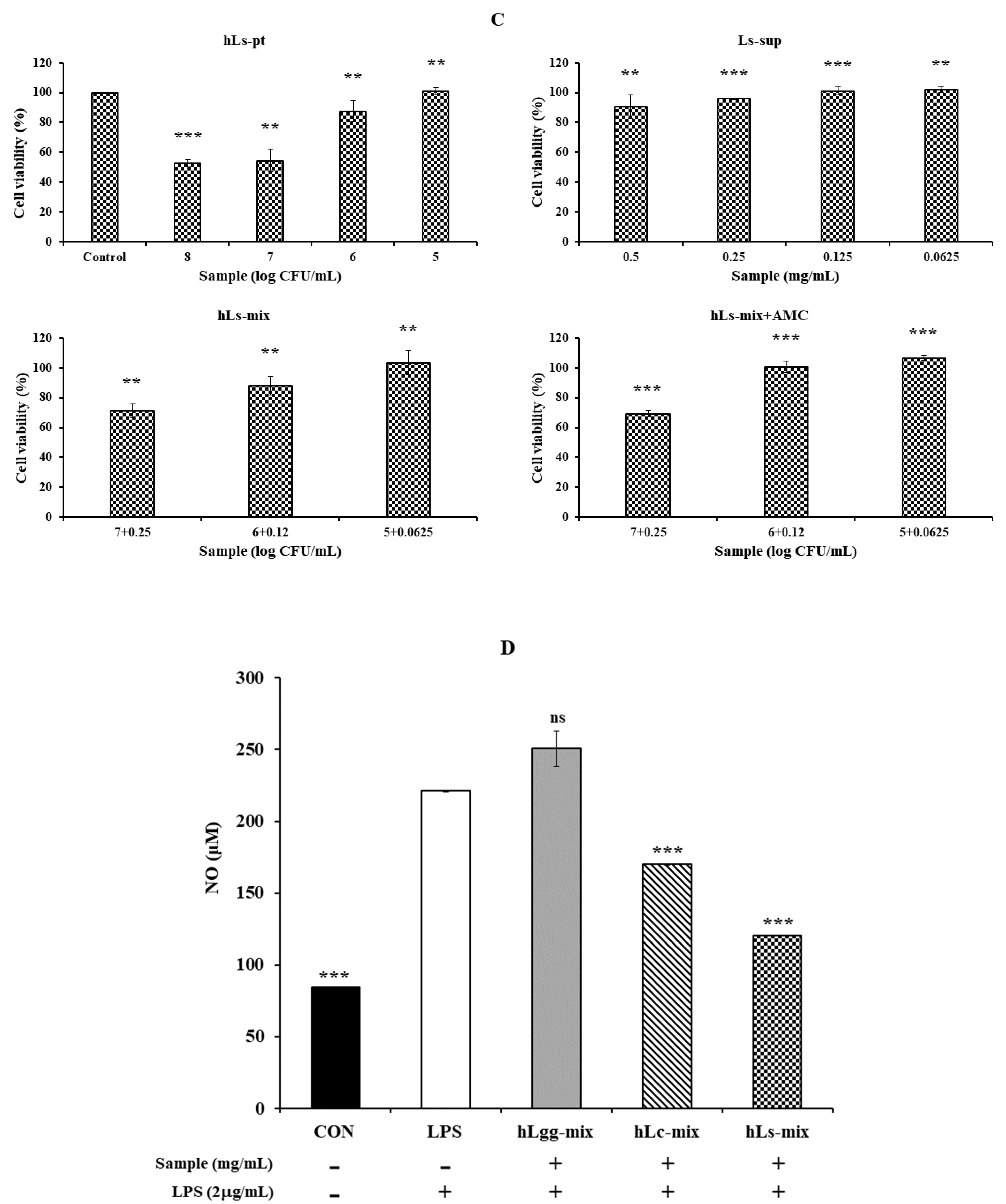
2.5. Cell Viability
2.6. Quantitative Analysis of NO Production
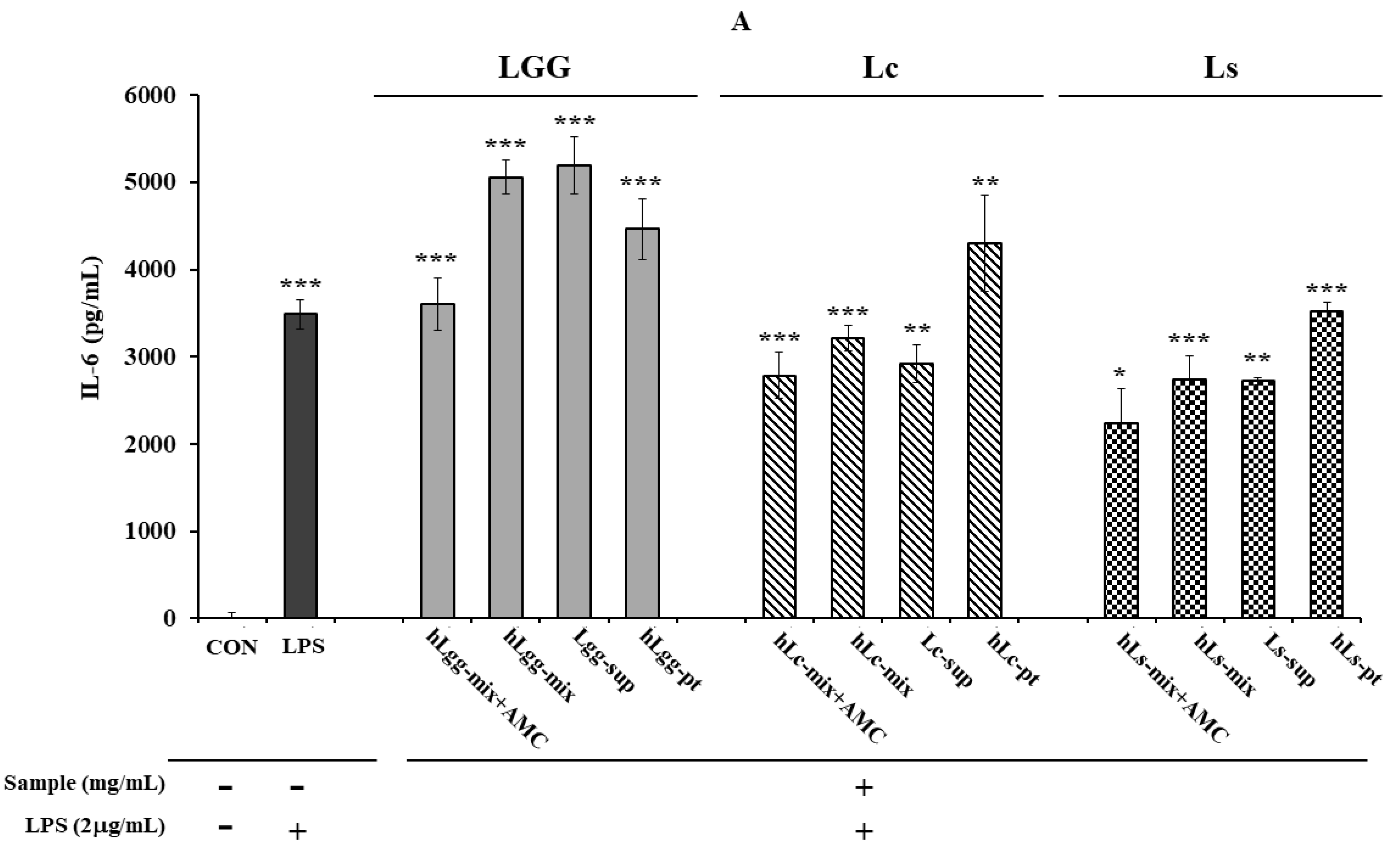
2.7. Quantitative Analysis of TNF-α and IL-6
2.8. Statistical Analysis
3. Results
3.1. Isolation and Identification of Lactic Acid-Producing Bacteria from GRET
3.2. Biochemical Characteristics of Bacterial Strains L. curvatus and L. sakei Isolated from GRET
3.3. Effects of Lc and Ls on Cell Viability
3.4. Effects of Lc and Ls on NO and Inflammatory Cytokine Expression
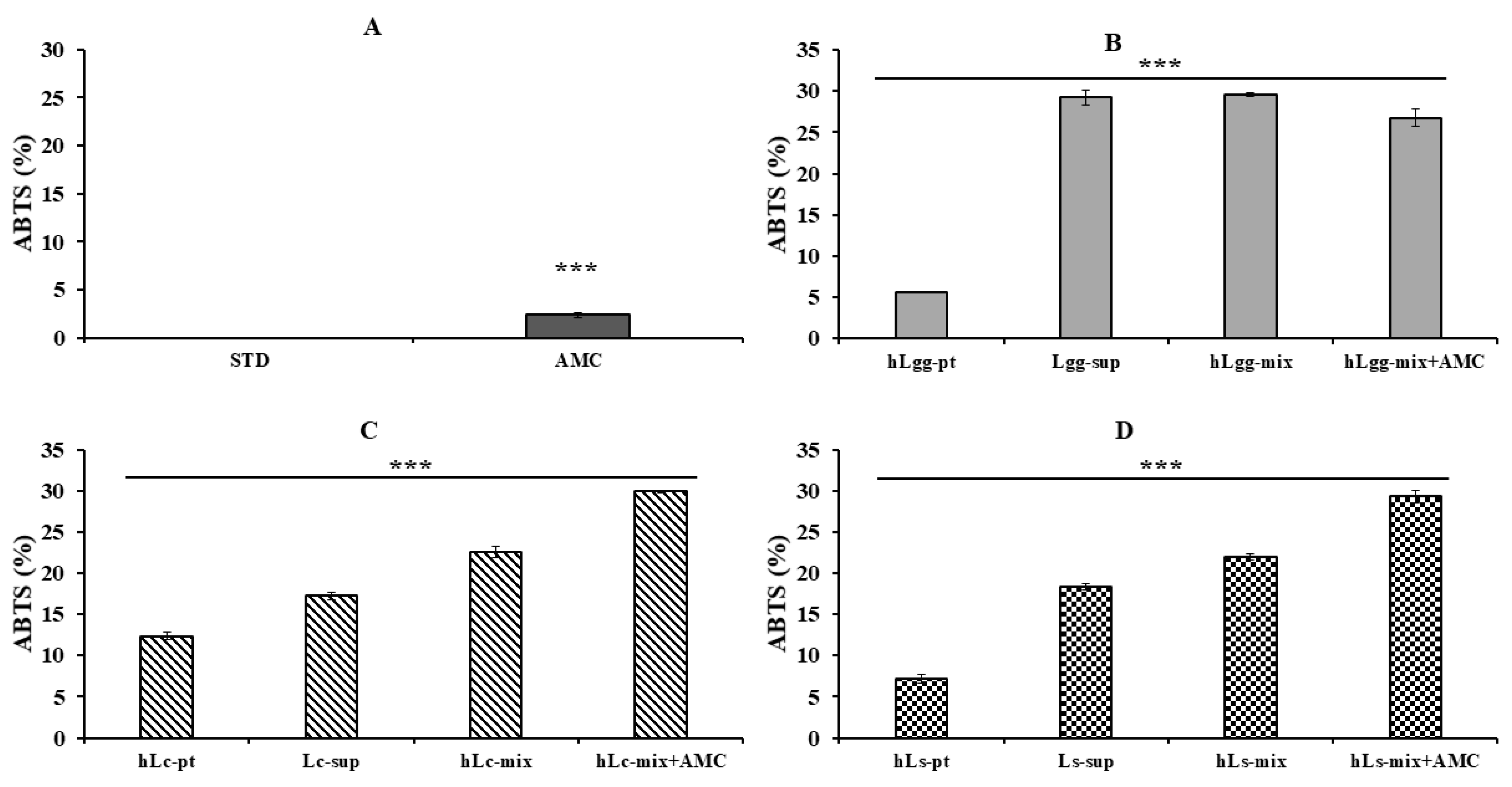
3.5. Antioxidant Effects of Lc and Ls
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Safari, R.; Hoseinifar, S.H.; Nejadmoghadam, S.; Khalili, M. Apple cider vinegar boosted immunomodulatory and health promoting effects of Lactobacillus casei in common carp (Cyprinus carpio). Fish Shellfish Immunol. 2017, 67, 441–448. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef]
- Ma, Y.H.; Sheng, Y.D.; Zhang, D.; Liu, J.T.; Tian, Y.; Li, H.; Li, X.F.; Li, N.; Sun, P.; Siddiqui, S.A.; et al. Acanthopanax senticosus cultures fermented by Lactobacillus rhamnosus enhanced immune response through improvement of antioxidant activity and inflammation in crucian carp (Carassius auratus). Microb. Pathog. 2024, 190, 106614. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, S.J.; Mehrabani, D.; Saberifiroozi, M.; Fattahi, M.R.; Moradi, F.; Najafi, M. The effect of yogurt fortified with Lactobacillus acidophilus and Bifidobacterium sp. probiotic in patients with lactose intolerance. Food Sci. Nutr. 2021, 9, 1704–1711. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front. Microbiol. 2018, 9, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Blackwood, B.P.; Yuan, C.Y.; Wood, D.R.; Nicolas, J.D.; Grothaus, J.S.; Hunter, C.J. Probiotic Lactobacillus species strengthen intestinal barrier function and tight junction integrity in experimental necrotizing enterocolitis. J. Probiotics Health 2017, 5, 159. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Chiba, E.; Tomosada, Y.; Salva, S.; Marranzino, G.; Kitazawa, H.; Alvarez, S. Orally administered Lactobacillus rhamnosus modulates the respiratory immune response triggered by the viral pathogen-associated molecular pattern poly(I:C). BMC Immunol. 2012, 13, 53. [Google Scholar] [CrossRef]
- Rastogi, S.; Singh, A. Gut microbiome and human health: Exploring how the probiotic genus Lactobacillus modulate immune responses. Front. Pharmacol. 2022, 13, 1042189. [Google Scholar] [CrossRef]
- Song, M.; Yun, B.; Moon, J.-H.; Park, D.-J.; Lim, K.; Oh, S. Characterization of selected Lactobacillus strains for use as probiotics. Korean J. Food Sci. Anim. Resour. 2015, 35, 551–556. [Google Scholar] [CrossRef]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef]
- López-López, A.; Camelo-Castillo, A.; Ferrer, M.D.; Simon-Soro, Á.; Mira, A. Health-associated niche inhabitants as oral probiotics: The case of Streptococcus dentisani. Front. Microbiol. 2017, 8, 379. [Google Scholar] [CrossRef]
- Capurso, L. Thirty years of Lactobacillus rhamnosus GG: A review. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef]
- Pagnini, C.; Corleto, V.D.; Martorelli, M.; Lanini, C.; D’Ambra, G.; Di Giulio, E.; Delle Fave, G. Mucosal adhesion and anti-inflammatory effects of Lactobacillus rhamnosus GG in the human colonic mucosa: A proof-of-concept study. World J. Gastroenterol. 2018, 24, 4652–4662. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef] [PubMed]
- Vougiouklaki, D.; Tsironi, T.; Tsantes, A.G.; Tsakali, E.; Van Impe, J.F.; Houhoula, D. Probiotic properties and antioxidant activity in vitro of lactic acid bacteria. Microorganisms 2023, 11, 1264. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.K.Y.; Tai, E.K.K.; Koo, M.W.L.; Wong, H.P.S.; Wu, W.K.K.; Yu, L.; So, W.H.L.; Woo, P.C.Y.; Cho, C.H. Enhancement of gastric mucosal integrity by Lactobacillus rhamnosus GG. Life Sci. 2007, 80, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.; Baird, C.H.; Rush, A.A.; Barnes, C.; Wischmeyer, P.E. Lactobacillus rhamnosus GG treatment improves intestinal permeability and modulates inflammatory response and homeostasis of spleen and colon in experimental model of Pseudomonas aeruginosa pneumonia. Clin. Nutr. 2017, 36, 1549–1557. [Google Scholar] [CrossRef]
- Elfaki, M.O.; Abdelatti, K.A. Rumen content as animal feed: A review. J. Vet. Med. Anim. Prod. 2016, 7, 80–88. [Google Scholar]
- Agbabiaka, L.; Madubuike, F.; Amadi, S. Studies on nutrients and anti-nutrients of rumen digesta from three most domesticated ruminants in Nigeria. Pak. J. Nutr. 2012, 11, 580–582. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Ossa, F. The rumen microbiome: Composition, abundance, diversity, and new investigative tools. Prof. Anim. Sci. 2014, 30, 1–12. [Google Scholar] [CrossRef]
- Jami, E.; Mizrahi, I. Composition and similarity of bovine rumen microbiota across individual animals. PLoS ONE 2012, 7, e33306. [Google Scholar] [CrossRef]
- Jose, N.M.; Bunt, C.R.; Hussain, M.A. Comparison of microbiological and probiotic characteristics of lactobacilli isolates from dairy food products and animal rumen contents. Microorganisms 2015, 3, 198–212. [Google Scholar] [CrossRef]
- Broadbent, J.R.; Larsen, R.L.; Deibel, V.; Steele, J.L. Physiological and transcriptional response of Lactobacillus casei ATCC 334 to acid stress. J. Bacteriol. 2010, 192, 2445–2458. [Google Scholar] [CrossRef]
- Fuochi, V.; Petronio, G.P.; Lissandrello, E.; Furneri, P.M. Evaluation of resistance to low pH and bile salts of human Lactobacillus spp. isolates. Int. J. Immunopathol. Pharmacol. 2015, 28, 426–433. [Google Scholar] [CrossRef]
- Sabino, Y.N.V.; Santana, M.F.; Oyama, L.B.; Santos, F.G.; Moreira, A.J.S.; Huws, S.A.; Mantovani, H.C. Characterization of antibiotic resistance genes in the species of the rumen microbiota. Nat. Commun. 2019, 10, 5252. [Google Scholar] [CrossRef]
- Hu, G.; Jiang, H.; Zong, Y.; Datsomor, O.; Kou, L.; An, Y.; Zhao, J.; Miao, L. Characterization of lactic acid-producing bacteria isolated from rumen: Growth, acid and bile salt tolerance, and antimicrobial function. Fermentation 2022, 8, 385. [Google Scholar] [CrossRef]
- Ladha, G.; Jeevaratnam, K. A novel antibacterial compound produced by Lactobacillus plantarum LJR13 isolated from rumen liquor of goat effectively controls multi-drug resistant human pathogens. Microbiol. Res. 2020, 241, 126563. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.S.; Lee, N.K.; Paik, H.D. Heat-killed Latilactobacillus sakei CNSC001WB and Lactobacillus pentosus WB693 have an anti-inflammatory effect on LPS-stimulated RAW 264.7 cells. Probiotics Antimicrob. Proteins 2024, 16, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Shin, J.S.; Lee, S.G.; Rhee, Y.K.; Cho, C.W.; Hong, H.D.; Lee, K.T. Lactobacillus sakei K040706 evokes immunostimulatory effects on macrophages through TLR 2-mediated activation. Int. Immunopharmacol. 2015, 28, 88–96. [Google Scholar] [CrossRef]
- Hyun, J.H.; Woo, I.K.; Kim, K.T.; Park, Y.S.; Kang, D.K.; Lee, N.K.; Paik, H.D. Heat-treated paraprobiotic Latilactobacillus sakei KU15041 and Latilactobacillus curvatus KU15003 show an antioxidant and immunostimulatory effect. J. Microbiol. Biotechnol. 2023, 34, 358–366. [Google Scholar] [CrossRef]
- Zagorec, M.; Champomier-Vergès, M.C. Lactobacillus sakei: A starter for sausage fermentation, a protective culture for meat products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef]
- Janßen, D.; Eisenbach, L.; Ehrmann, M.A.; Vogel, R.F. Assertiveness of Lactobacillus sakei and Lactobacillus curvatus in a fermented sausage model. Int. J. Food Microbiol. 2018, 285, 188–197. [Google Scholar] [CrossRef]
- Zommiti, M.; Connil, N.; Hamida, B.J.; Ferchichi, M. Probiotic characteristics of Lactobacillus curvatus DN317, a strain isolated from chicken ceca. Probiotics Antimicrob. Proteins 2017, 9, 415–424. [Google Scholar] [CrossRef]
- Castilho, N.P.A.; Colombo, M.; Oliveira LLde Todorov, S.D.; Nero, L.A. Lactobacillus curvatus UFV-NPAC1 and other lactic acid bacteria isolated from calabresa, a fermented meat product, present high bacteriocinogenic activity against Listeria monocytogenes. BMC Microbiol. 2019, 19, 63. [Google Scholar] [CrossRef]
- Luan, C.; Jiang, N.; Zhou, X.; Zhang, C.; Zhao, Y.; Li, Z.; Li, C. Antibacterial and anti-biofilm activities of probiotic Lactobacillus curvatus BSF206 and Pediococcus pentosaceus AC1-2 against Streptococcus mutans. Microb. Pathog. 2022, 164, 105446. [Google Scholar] [CrossRef]
- Zhu, J.; Tan, Z.; Zhang, Z.; Shi, X. Characterization on structure and bioactivities of an exopolysaccharide from Lactobacillus curvatus SJTUF 62116. Int. J. Biol. Macromol. 2022, 210, 504–517. [Google Scholar] [CrossRef]
- Melian, C.; Ploper, D.; Chehín, R.; Vignolo, G.; Castellano, P. Impairment of Listeria monocytogenes biofilm developed on industrial surfaces by Latilactobacillus curvatus CRL1579 bacteriocin. Food Microbiol. 2024, 121, 104491. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Choi, S.H.; Ju, S.H.; Park, J.K. Antifungal effect of chitosan hydrolysates against ginseng infectious fungi Fusarium foetens. J. Chitin Chitosan 2024, 29, 184–194. [Google Scholar] [CrossRef]
- Park, J.K.; Jeong, D.Y.; Shin, D.C.; Lee, J.H.; Kim, H.E.; Kim, G.H.; Lee, Y.H. Effects of chitosan as a growth regulator of the mealworm Tenebrio molitor. J. Chitin Chitosan 2023, 28, 1–10. [Google Scholar] [CrossRef]
- Na, Y.S.; Kim, W.J.; Kim, S.M.; Park, J.K.; Lee, S.M.; Kim, S.O.; Synytsya, A.; Park, Y.I. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar] [CrossRef]
- Kim, K.W. Isolation and characterization of a bacterial isolate for polyhydroxyalkanoates production. J. Chitin Chitosan 2014, 19, 87–92. [Google Scholar]
- Yoo, A.Y.; Park, J.K. Isolation and characterization of a serine protease-producing marine bacterium Marinomonas arctica PT-1. Bioprocess Biosyst. Eng. 2016, 39, 307–314. [Google Scholar] [CrossRef]
- Avelelas, F.; Horta, A.; Pinto, L.F.; Cotrim Marques, S.; Marques Nunes, P.; Pedrosa, R.; Leandro, S.M. Antifungal and antioxidant properties of chitosan polymers obtained from nontraditional Polybius henslowii sources. Mar. Drugs 2019, 17, 239. [Google Scholar] [CrossRef] [PubMed]
- Xing, R.; Yu, H.; Liu, S.; Zhang, W.; Zhang, Q.; Li, Z.; Li, P. Antioxidant activity of differently regioselective chitosan sulfates in vitro. Bioorg. Med. Chem. 2005, 13, 1387–1392. [Google Scholar] [CrossRef]
- Azuma, K.; Osaki, T.; Minami, S.; Okamoto, Y. Anticancer and anti-inflammatory properties of chitin and chitosan oligosaccharides. J. Funct. Biomater. 2015, 6, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Facchin, B.M.; Dos Reis, G.O.; Vieira, G.N.; Mohr, E.T.B.; da Rosa, J.S.; Kretzer, I.F.; Demarchi, I.G.; Dalmarco, E.M. Inflammatory biomarkers on an LPS-induced RAW 264.7 cell model: A systematic review and meta-analysis. Inflamm. Res. 2022, 71, 741–758. [Google Scholar] [CrossRef]
- Noh, H.J.; Park, J.M.; Kwon, Y.J.; Kim, K.; Park, S.Y.; Kim, I.; Lim, J.H.; Kim, B.K.; Kim, B.Y. Immunostimulatory effect of heat-killed probiotics on RAW264.7 macrophages. J. Microbiol. Biotechnol. 2022, 32, 638–644. [Google Scholar] [CrossRef]
- Ivec, M.; Botić, T.; Koren, S.; Jakobsen, M.; Weingartl, H.; Cencič, A. Interactions of macrophages with probiotic bacteria lead to increased antiviral response against vesicular stomatitis virus. Antivir. Res. 2007, 75, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, C.; Li, Y.; Li, Z.; Li, C.; Luan, C. Antioxidant effects and probiotic properties of Latilactobacillus sakei MS103 isolated from sweet pickled garlic. Foods 2023, 12, 4276. [Google Scholar] [CrossRef]
- Duz, M.; Dogan, Y.N.; Dogan, I. Antioxidant activity of Lactobacillus plantarum, Lactobacillus sakei and Lactobacillus curvatus strains isolated from fermented Turkish Sucuk. An. Acad. Bras. Ciênc. 2020, 92, e20200105. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.H.; Choi, S.H.; Lee, Y.H.; Park, J.K. Antioxidant and Anti-Inflammatory Activities of Latilactobacillus curvatus and L. sakei Isolated from Green Tripe. Nutrients 2025, 17, 2464. https://doi.org/10.3390/nu17152464
Lee GH, Choi SH, Lee YH, Park JK. Antioxidant and Anti-Inflammatory Activities of Latilactobacillus curvatus and L. sakei Isolated from Green Tripe. Nutrients. 2025; 17(15):2464. https://doi.org/10.3390/nu17152464
Chicago/Turabian StyleLee, Ga Hun, Sung Hyun Choi, Yong Hyun Lee, and Jae Kweon Park. 2025. "Antioxidant and Anti-Inflammatory Activities of Latilactobacillus curvatus and L. sakei Isolated from Green Tripe" Nutrients 17, no. 15: 2464. https://doi.org/10.3390/nu17152464
APA StyleLee, G. H., Choi, S. H., Lee, Y. H., & Park, J. K. (2025). Antioxidant and Anti-Inflammatory Activities of Latilactobacillus curvatus and L. sakei Isolated from Green Tripe. Nutrients, 17(15), 2464. https://doi.org/10.3390/nu17152464