
Non-Alcoholic Beer Influences Glucose and Lipid Metabolism and Changes Body Composition in Healthy, Young, Male Adults

 , , ,
, , ,
Abstract
1. Introduction
2. Material and Methods
2.1. Patient Recruitment, Ethical Statement, and Sample Collection
2.2. Assessment of Liver Steatosis via Transient Elastography
2.3. Collection of Fecal Samples and Preparation for Sequencing
2.4. Sequencing Data Analysis
2.5. Statistical Analysis
3. Results
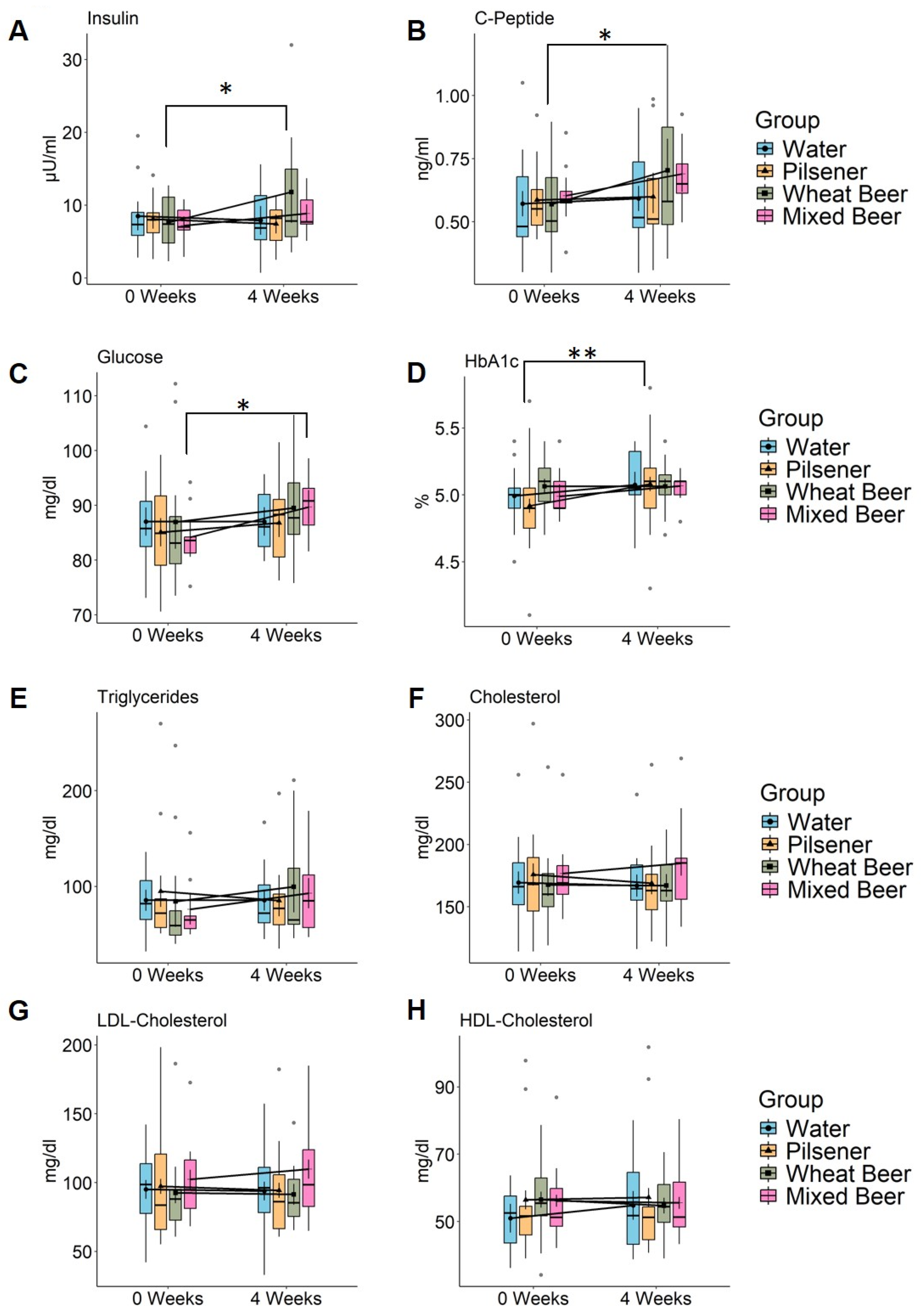
3.1. Impact of Mixed Beer and Wheat Beer on Glucose and Fat Metabolism
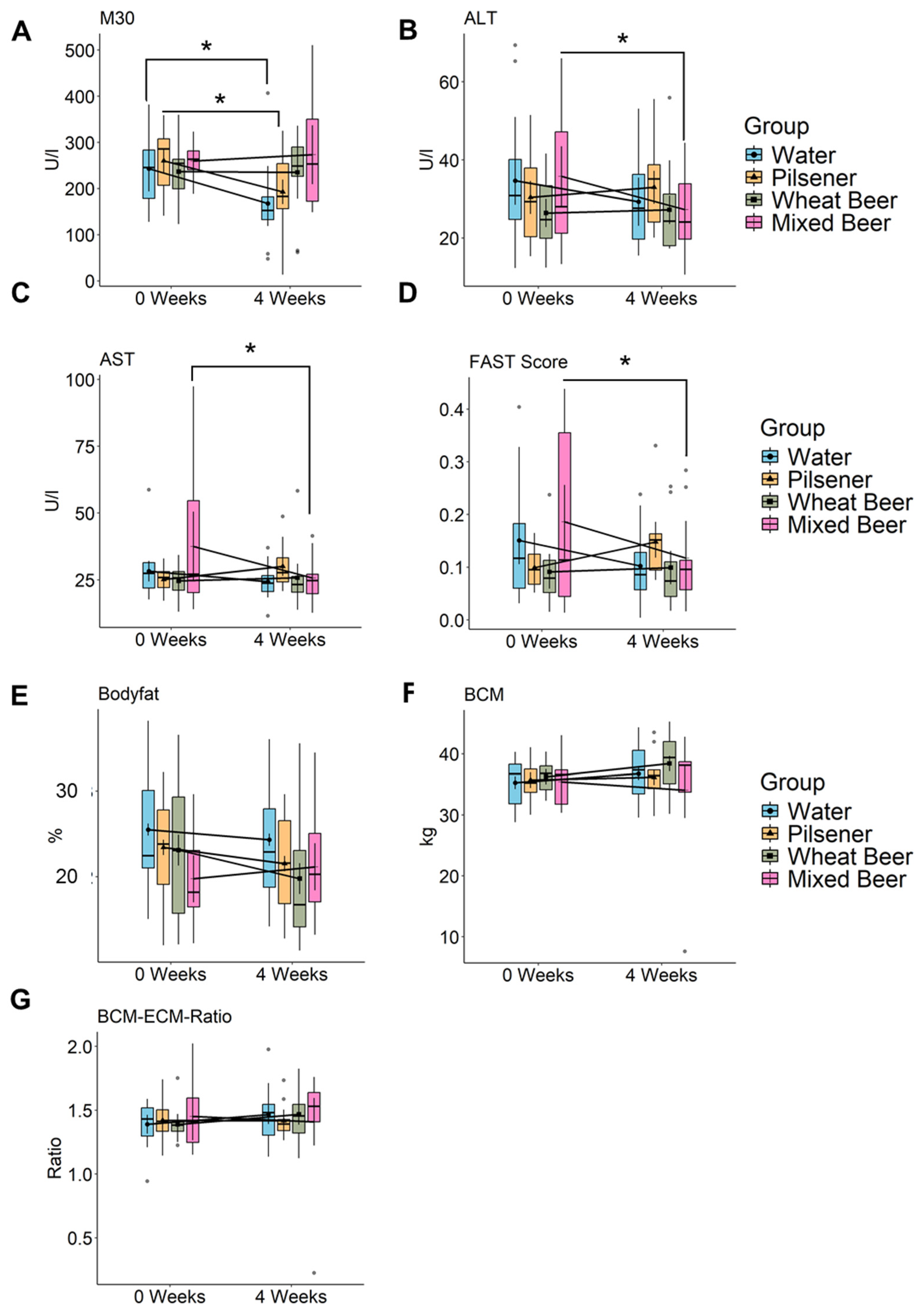
3.2. Impact of Water, Non-Alcoholic Pilsener, and Non-Alcoholic Mixed Beer on Liver Damage
3.3. Liver Steatosis and Stiffness After Abstinence from Drinks Other than Water After 4 Weeks
3.4. Wheat and Mixed Beer Influence Body Composition
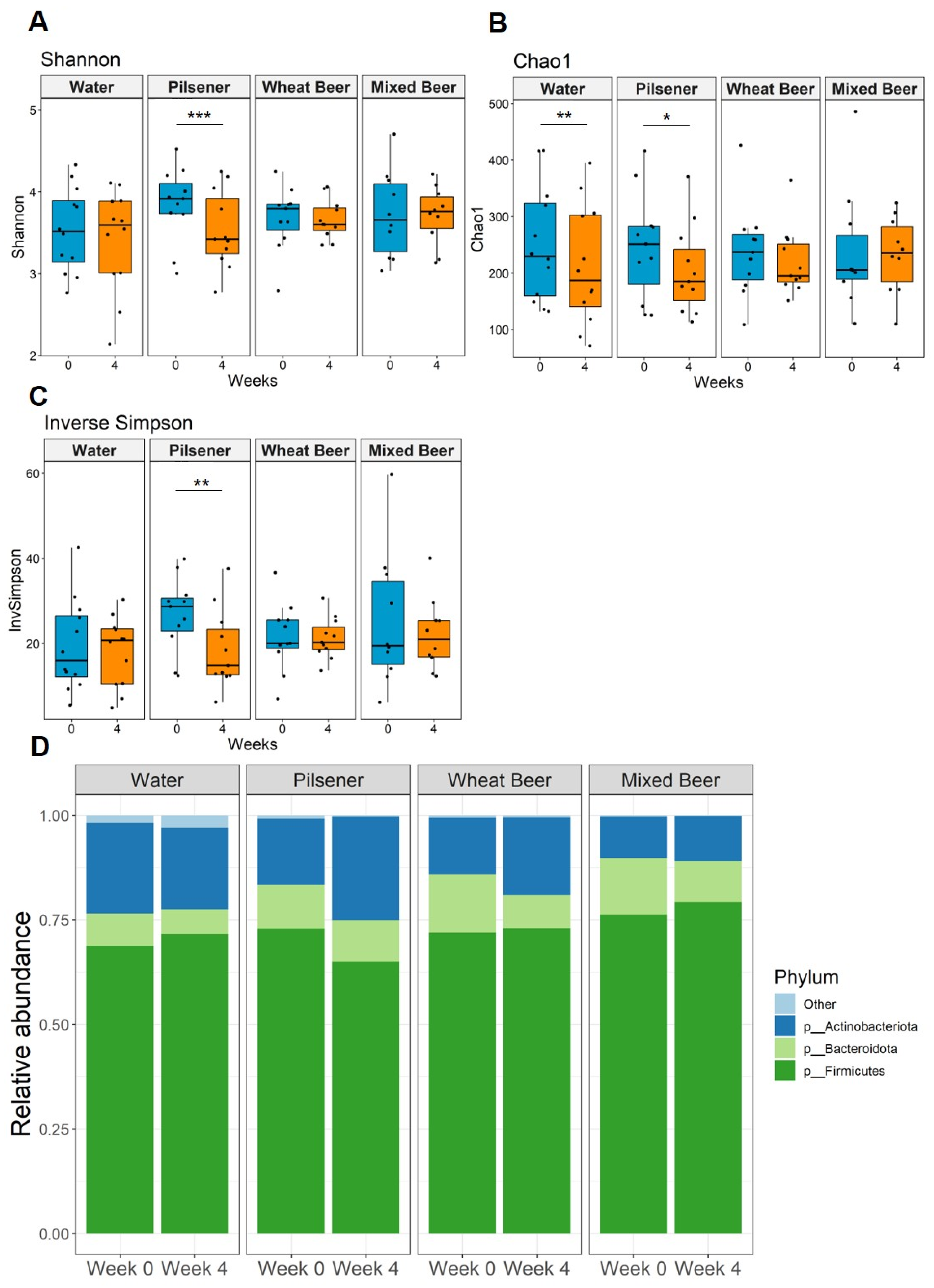
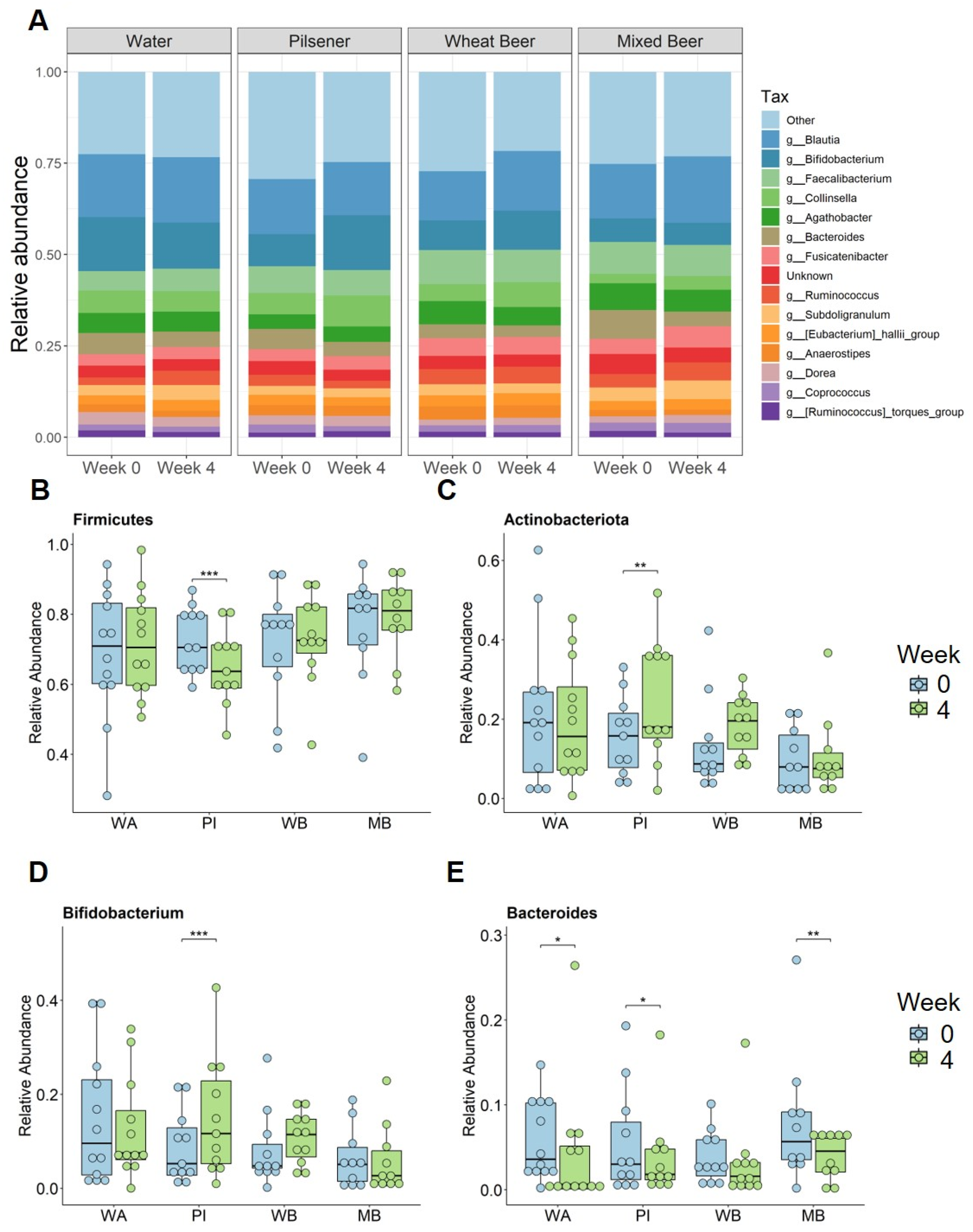
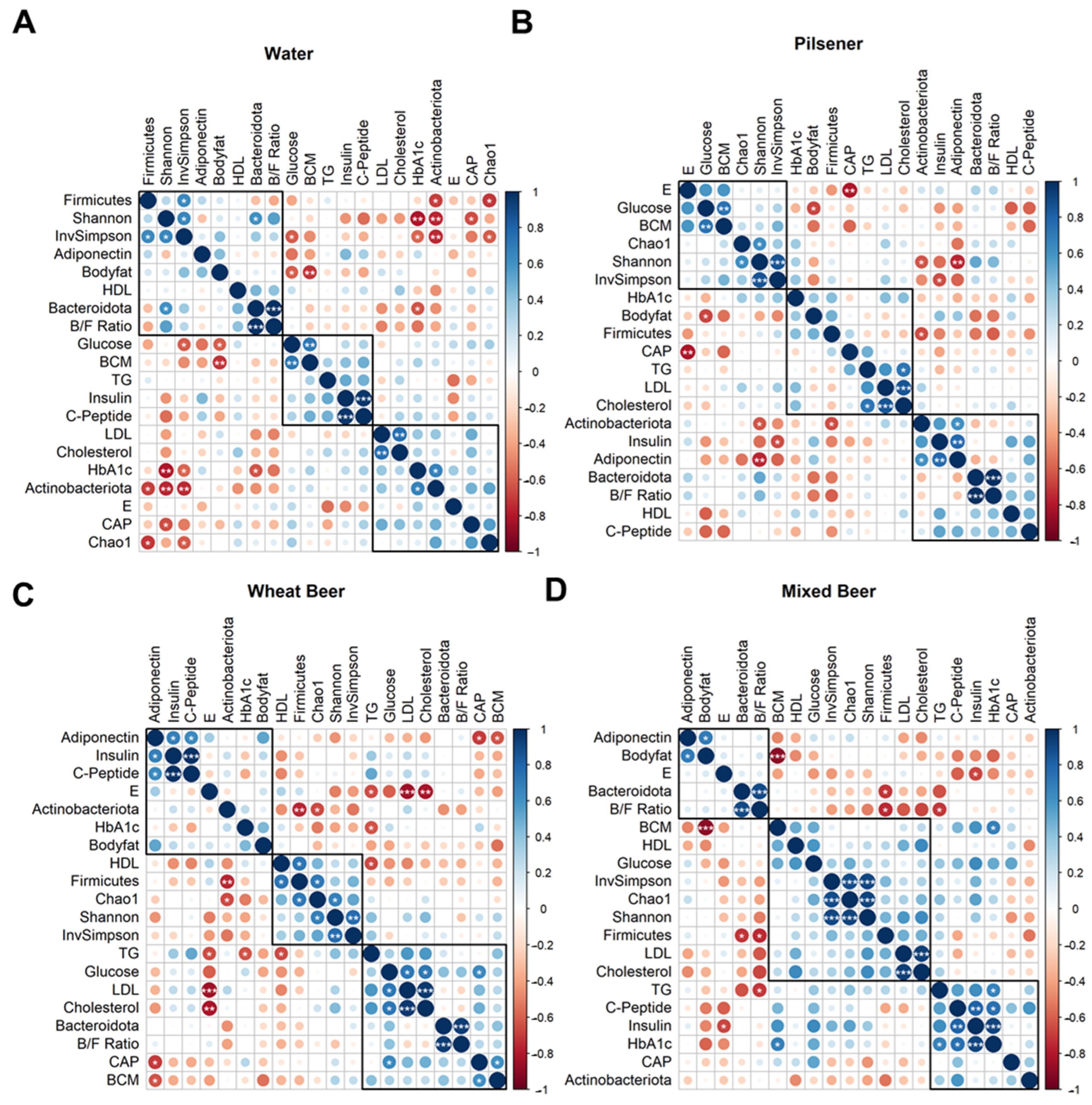
3.5. Pilsener Consumption Decreased Alpha Diversity and Firmicutes Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kokole, D.; Jané Llopis, E.; Anderson, P. Non-alcoholic beer in the European Union and UK: Availability and apparent consumption. Drug Alcohol. Rev. 2022, 41, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Griswold, M.G.; Fullman, N.; Hawley, C.; Arian, N.; Zimsen, S.R.; Tymeson, H.D.; Venkateswaran, V.; Tapp, A.D.; Forouzanfar, M.H.; Salama, J.S.; et al. Alcohol use and burden for 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2018, 392, 1015–1035. [Google Scholar] [CrossRef] [PubMed]
- Bryazka, D.; Reitsma, M.B.; Griswold, M.G.; Abate, K.H.; Abbafati, C.; Abbasi-Kangevari, M.; Abbasi-Kangevari, Z.; Abdoli, A.; Abdollahi, M.; Abdullah, A.Y.M.; et al. Population-level risks of alcohol consumption by amount, geography, age, sex, and year: A systematic analysis for the Global Burden of Disease Study 2020. Lancet 2022, 400, 185–235. [Google Scholar] [CrossRef] [PubMed]
- Baik, I.; Shin, C. Prospective study of alcohol consumption and metabolic syndrome. Am. J. Clin. Nutr. 2008, 87, 1455–1463. [Google Scholar] [CrossRef]
- Naudin, S.; Li, K.; Jaouen, T.; Assi, N.; Kyrø, C.; Tjønneland, A.; Overvad, K.; Boutron-Ruault, M.; Rebours, V.; Védié, A.; et al. Lifetime and baseline alcohol intakes and risk of pancreatic cancer in the European Prospective Investigation into Cancer and Nutrition study. Int. J. Cancer 2018, 143, 801–812. [Google Scholar] [CrossRef]
- Larsen, B.A.; Klinedinst, B.S.; Le, S.T.; Pappas, C.; Wolf, T.; Meier, N.F.; Lim, Y.L.; Willette, A.A. Beer, wine, and spirits differentially influence body composition in older white adults–a United Kingdom Biobank study. Obes. Sci. Pract. 2022, 8, 641–656. [Google Scholar] [CrossRef]
- Shield, K.D.; Parry, C.; Rehm, J. Chronic Diseases and Conditions Related to Alcohol Use 155 Chronic Diseases and Conditions Related to Alcohol Use. Alcohol Res. Curr. Rev. 2014, 35, 155. [Google Scholar]
- Anderson, B.O.; Berdzuli, N.; Ilbawi, A.; Kestel, D.; Kluge, H.P.; Krech, R.; Mikkelsen, B.; Neufeld, M.; Poznyak, V.; Rekve, D.; et al. Comment Health and cancer risks associated with low levels of alcohol consumption. Lancet Public. Health 2022, 8, e6–e7. [Google Scholar] [CrossRef]
- Hernández-Quiroz, F.; Nirmalkar, K.; Villalobos-Flores, L.E.; Murugesan, S.; Cruz-Narváez, Y.; Rico-Arzate, E.; Hoyo-Vadillo, C.; Chavez-Carbajal, A.; Pizano-Zárate, M.L.; García-Mena, J. Influence of moderate beer consumption on human gut microbiota and its impact on fasting glucose and β-cell function. Alcohol 2020, 85, 77–94. [Google Scholar] [CrossRef]
- Trius-Soler, M.; Marhuenda-Muñoz, M.; Laveriano-Santos, E.P.; Martínez-Huélamo, M.; Sasot, G.; Storniolo, C.E.; Estruch, R.; Lamuela-Raventós, R.M.; Tresserra-Rimbau, A. Moderate consumption of beer (with and without ethanol) and menopausal symptoms: Results from a parallel clinical trial in postmenopausal women. Nutrients 2021, 13, 2278. [Google Scholar] [CrossRef]
- Martínez-montoro, J.I.; Quesada-molina, M.; Gutiérrez-repiso, C.; Ruiz-limón, P.; Subiri-verdugo, A.; Tinahones, F.J.; Moreno-indias, I. Effect of Moderate Consumption of Different Phenolic-Content Beers on the Human Gut Microbiota Composition: A Randomized Crossover Trial. Antioxidants 2022, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- González-Zancada, N.; Redondo-Useros, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A.; Nova, E. Association of moderate beer consumption with the gut microbiota and SCFA of healthy adults. Molecules 2020, 25, 4772. [Google Scholar] [CrossRef] [PubMed]
- Martínez Alvarez, J.R.; Bellés, V.V.; López-Jaén, A.B.; Marín, A.V.; Codoñer-Franch, P. Effects of alcohol-free beer on lipid profile and parameters of oxidative stress and inflammation in elderly women. Nutrition 2009, 25, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Padro, T.; Muñoz-García, N.; Vilahur, G.; Chagas, P.; Deyà, A.; Antonijoan, R.M.; Badimon, L. Moderate beer intake and cardiovascular health in overweight individuals. Nutrients 2018, 10, 1237. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Lahti, L.; Shetty, S. Microbiome R Package: Tools for Microbiome Analysis in R. (2012–2019). Available online: https://github.com/microbiome/microbiome (accessed on 4 May 2025).
- Sowa, J.P.; Heider, D.; Bechmann, L.P.; Gerken, G.; Hoffmann, D.; Canbay, A. Novel algorithm for non-invasive assessment of fibrosis in NAFLD. PLoS ONE 2013, 8, e62439. [Google Scholar] [CrossRef]
- Diab, D.L.; Yerian, L.; Schauer, P.; Kashyap, S.R.; Lopez, R.; Hazen, S.L.; Feldstein, A.E. Cytokeratin 18 fragment levels as a noninvasive biomarker for nonalcoholic steatohepatitis in bariatric surgery patients. Clin. Gastroenterol. Hepatol. 2008, 6, 1249–1254. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Wieckowska, A.; Lopez, A.R.; Liu, Y.C.; Zein, N.N.; McCullough, A.J. Cytokeratin-18 fragment levels as noninvasive biomarkers for nonalcoholic steatohepatitis: A multicenter validation study. Hepatology 2009, 50, 1072–1078. [Google Scholar] [CrossRef]
- Lemmer, P.; Manka, P.; Best, J.; Kahraman, A.; Kälsch, J.; Vilchez-Vargas, R.; Link, A.; Chiang, H.; Gerken, G.; Canbay, A.; et al. Effects of Moderate Alcohol Consumption in Non-Alcoholic Fatty Liver Disease. J. Clin. Med. 2022, 11, 890. [Google Scholar] [CrossRef]
- Ma, J.; Fox, C.S.; Jacques, P.F.; Speliotes, E.K.; Hoffmann, U.; Smith, C.E.; Saltzman, E.; McKeown, N.M. Sugar-sweetened beverage, diet soda, and fatty liver disease in the Framingham Heart Study cohorts. J. Hepatol. 2015, 63, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Aeberli, I.; Gerber, P.A.; Hochuli, M.; Kohler, S.; Haile, S.R.; Gouni-Berthold, I.; Berthold, H.K.; Spinas, G.A.; Berneis, K. Low to moderate sugar-sweetened beverage consumption impairs glucose and lipid metabolism and promotes inflammation in healthy young men: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 94, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Manka, P.; Olliges, V.; Bechmann, L.P.; Schlattjan, M.; Jochum, C.; Treckmann, J.W.; Saner, F.H.; Gerken, G.; Syn, W.K.; Canbay, A. Low levels of blood lipids are associated with etiology and lethal outcome in acute liver failure. PLoS ONE 2014, 9, e102351. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, A.; Forte, M.; Nardini, M. Characterization of Phenolics Content and Antioxidant Activity of Different Beer Types. J. Agric. Food Chem. 2010, 58, 10677–10683. [Google Scholar] [CrossRef]
- Di Domenico, M.; Feola, A.; Ambrosio, P.; Pinto, F.; Galasso, G.; Zarrelli, A.; Di Fabio, G.; Porcelli, M.; Scacco, S.; Inchingolo, F.; et al. Antioxidant Effect of Beer Polyphenols and Their Bioavailability in Dental-Derived Stem Cells (D-dSCs) and Human Intestinal Epithelial Lines (Caco-2) Cells. Stem Cells Int. 2020, 2020, 8835813. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Cheung, F.; Hong, M.; Feng, Y. The Potential and Action Mechanism of Polyphenols in the Treatment of Liver Diseases. Oxid. Med. Cell Longev. 2018, 2018, 8394818. [Google Scholar] [CrossRef]
- Beulens, J.W.J.; van den Berg, R.; Kok, F.J.; Helander, A.; Vermunt, S.H.F.; Hendriks, H.F.J. Moderate alcohol consumption and lipoprotein-associated phospholipase A2 activity. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 539–544. [Google Scholar] [CrossRef]
- Degrace, P.; Moindrot, B.; Mohamed, I.; Gresti, J.; Clouet, P. Moderate consumption of beer reduces liver triglycerides and aortic cholesterol deposit in LDLr-/- apoB100/100 mice. Atherosclerosis 2006, 189, 328–335. [Google Scholar] [CrossRef]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Condines, X.; Magraner, E.; Roth, I.; Valderas-Martínez, P.; Arranz, S.; Casas, R.; Martínez-Huélamo, M.; Vallverdú-Queralt, A.; Quifer-Rada, P.; et al. The non-alcoholic fraction of beer increases stromal cell derived factor 1 and the number of circulating endothelial progenitor cells in high cardiovascular risk subjects: A randomized clinical trial. Atherosclerosis 2014, 233, 518–524. [Google Scholar] [CrossRef]
- De Gaetano, G.; Costanzo, S.; Di Castelnuovo, A.; Badimon, L.; Bejko, D.; Alkerwi, A.; Chiva-Blanch, G.; Estruch, R.; La Vecchia, C.; Panico, S.; et al. Effects of moderate beer consumption on health and disease: A consensus document. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 443–467. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Yoshida, H. Beneficial effects of adiponectin on glucose and lipid metabolism and atherosclerotic progression: Mechanisms and perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: Adiponectin. Nat. Metab. 2019, 1, 334–339. [Google Scholar] [CrossRef]
- Leven, A.S.; Gieseler, R.K.; Schlattjan, M.; Schreiter, T.; Niedergethmann, M.; Baars, T.; Baba, H.A.; Özçürümez, M.K.; Sowa, J.P.; Canbay, A. Association of cell death mechanisms and fibrosis in visceral white adipose tissue with pathological alterations in the liver of morbidly obese patients with NAFLD. Adipocyte 2021, 10, 558–573. [Google Scholar] [CrossRef]
- Wree, A.; Schlattjan, M.; Bechmann, L.P.; Claudel, T.; Sowa, J.-P.; Stojakovic, T.; Scharnagl, H.; Köfeler, H.; Baba, H.A.; Gerken, G.; et al. Adipocyte cell size, free fatty acids and apolipoproteins are associated with non-alcoholic liver injury progression in severely obese patients. Metabolism 2014, 63, 1542–1552. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Rajakaruna, S.; Pastoriza, S.; Paliy, O.; Ángel Rufián-Henares, J. Bioactivity of food melanoidins is mediated by gut microbiota. Food Chem. 2020, 316, 126309. [Google Scholar] [CrossRef]
- Porras, D.; Nistal, E.; Martínez-Flórez, S.; Pisonero-Vaquero, S.; Olcoz, J.L.; Jover, R.; González-Gallego, J.; García-Mediavilla, M.V.; Sánchez-Campos, S. Protective effect of quercetin on high-fat diet-induced non-alcoholic fatty liver disease in mice is mediated by modulating intestinal microbiota imbalance and related gut-liver axis activation. Free Radic. Biol. Med. 2017, 102, 188–202. [Google Scholar] [CrossRef]
- Teixeira, C.; Prykhodko, O.; Alminger, M.; Fåk Hållenius, F.; Nyman, M. Barley Products of Different Fiber Composition Selectively Change Microbiota Composition in Rats. Mol. Nutr. Food Res. 2018, 62, e1701023. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Overall n = 44 | Water n = 12 | Pilsener n = 11 | Wheat Beer n = 11 | Mixed Beer n = 10 | p-Value |
|---|---|---|---|---|---|---|
| Body Characteristics | ||||||
| Height (cm) n = 44 | 183.0 (177.5, 186.5) | 183.0 (177.0, 185.5) | 183.0 (174.0, 190.0) | 185.0 (180.0, 186.0) | 179.0 (178.0, 187.0) | 0.8 |
| Weight (kg) n = 44 | 80 (73, 88) | 81 (77, 89) | 79 (72, 90) | 80 (74, 94) | 74 (68, 83) | 0.4 |
| BMI (kg/m2) n = 44 | 24.5 (22.1, 26.0) | 24.8 (24.1, 26.2) | 24.8 (21.6, 27.1) | 23.8 (21.4, 29.1) | 22.8 (21.4, 25.9) | 0.5 |
| Waist Circumference (cm) n = 44 | 83 (78, 88) | 85 (80, 86) | 86 (76, 96) | 85 (75, 99) | 79 (76, 83) | 0.3 |
| Hip Circumference (cm) n = 44 | 94.5 (91.5, 98.0) | 97.0 (93.0, 100.5) | 95.0 (91.0, 97.0) | 94.0 (90.0, 98.0) | 92.0 (91.0, 96.0) | 0.4 |
| Body fat (kg) n = 44 | 18 (13, 24) | 19 (17, 26) | 18 (13, 26) | 18 (11, 32) | 14 (11, 20) | 0.3 |
| Body Cell Mass (BCM) (kg) n = 44 | 36.3 (32.8, 38.0) | 36.7 (31.4, 38.5) | 35.2 (33.3, 37.7) | 36.8 (33.2, 38.2) | 36.6 (31.8, 37.5) | >0.9 |
| Extracellular Mass (ECM) (kg) n = 44 | 25.7 (24.0, 27.6) | 26.1 (23.6, 27.1) | 26.0 (22.4, 28.2) | 26.5 (25.6, 28.5) | 24.9 (23.9, 25.7) | 0.5 |
| LSM (kPa) n = 44 | 5.05 (3.2, 7.1) | 5.3 (3.7, 7.1) | 5.05 (3.5, 5.2) | 4.49 (3.3, 6.3) | 5.34 (3.2, 5.2) | 0.3 |
| Liver steatosis CAP (db/m2) n = 44 | 218.8 (152, 354) | 226.6 (181, 354) | 216.9 (162, 291) | 218.2 (152, 287) | 213.2 (172, 318) | >0.9 |
| Laboratory parameters | ||||||
| Bilirubin (mg/dL) n = 43 | 0.76 (0.60, 0.95) | 0.88 (0.75, 1.24) | 0.70 (0.47, 1.10) | 0.74 (0.60, 0.78) | 0.73 (0.57, 0.90) | 0.2 |
| Creatinine (mg/dL) n = 43 | 0.88 (0.82, 0.98) | 0.94 (0.86, 1.00) | 0.86 (0.77, 0.91) | 0.91 (0.85, 0.99) | 0.85 (0.79, 0.92) | 0.2 |
| AST (U/L) n = 43 | 27 (21, 30) | 27 (21, 32) | 26 (22, 29) | 26 (21, 29) | 27 (20, 55) | 0.9 |
| ALT (U/L) n = 43 | 28 (19, 42) | 31 (23, 44) | 29 (19, 44) | 25 (16, 34) | 28 (21, 47) | 0.8 |
| AP (U/L) n = 43 | 74 (64, 90) | 78 (63, 93) | 77 (73, 94) | 65 (59, 88) | 69 (67, 87) | 0.5 |
| GGT (U/L) n = 43 | 17 (14, 23) | 20 (15, 23) | 20 (14, 28) | 15 (12, 17) | 18 (16, 20) | 0.4 |
| LDH (U/L) n = 43 | 166 (157, 177) | 161 (154, 180) | 172 (166, 175) | 162 (154, 172) | 171 (164, 191) | 0.3 |
| Lipase (U/L) n = 43 | 28 (23, 39) | 28 (21, 42) | 27 (23, 38) | 27 (22, 31) | 30 (28, 39) | 0.8 |
| Albumin (g/dL) n = 43 | 48.10 (44.90, 49.60) | 46.90 (44.10, 49.65) | 48.80 (44.30, 50.50) | 48.10 (44.50, 49.20) | 48.10 (45.80, 49.20) | 0.8 |
| M30 (U/L) n = 43 | 255 (198, 286) | 245 (177, 286) | 286 (198, 316) | 254 (197, 268) | 264 (241, 281) | 0.6 |
| Adiponectin (µg/L) n = 43 | 5.1 (4.2, 9.1) | 4.8 (3.6, 8.4) | 5.1 (3.5, 7.5) | 7.5 (4.8, 10.1) | 5.1 (4.8, 6.5) | 0.5 |
| Glucose metabolism | ||||||
| Fasting glucose (mg/dL) n = 43 | 84 (81, 90) | 86 (82, 91) | 85 (76, 94) | 83 (78, 88) | 84 (81, 84) | 0.9 |
| C-Peptide (nmol/L) n = 43 | 0.56 (0.45, 0.67) | 0.48 (0.44, 0.69) | 0.55 (0.45, 0.65) | 0.50 (0.43, 0.70) | 0.59 (0.57, 0.62) | 0.9 |
| HbA1c (%) n = 43 | 5.00 (4.80, 5.20) | 5.00 (4.90, 5.10) | 4.90 (4.70, 5.10) | 5.10 (4.90, 5.20) | 4.90 (4.90, 5.10) | 0.5 |
| Insulin (µU/mL) n = 43 | 7.4 (5.4, 10.2) | 7.4 (5.7, 9.5) | 8.0 (5.6, 9.1) | 7.4 (4.3, 11.9) | 7.0 (6.6, 9.3) | >0.9 |
| Fat metabolism | ||||||
| Cholesterol (mg/dL) n = 43 | 167 (148, 183) | 166 (149, 191) | 168 (145, 200) | 160 (148, 183) | 169 (160, 183) | >0.9 |
| Triglycerides (mg/dL) n = 43 | 69 (56, 106) | 83 (66, 116) | 72 (56, 98) | 59 (48, 76) | 65 (56, 69) | 0.3 |
| HDL-Cholesterol (mg/dL) n = 43 | 53 (47, 61) | 53 (42, 59) | 52 (45, 55) | 55 (49, 64) | 51 (49, 60) | 0.6 |
| LDL-Cholesterol (mg/dL) n = 43 | 91 (77, 113) | 99 (77, 116) | 84 (65, 129) | 88 (69, 98) | 95 (81, 116) | 0.7 |
| Wheat Beer | Pilsener | Mixed Beer | ||||
|---|---|---|---|---|---|---|
| 100 mL | 660 mL | 100 mL | 660 mL | 100 mL | 660 mL | |
| Calories (kJ) | 107.0 | 706.2 | 111.0 | 732.6 | 114.0 | 752.4 |
| Fat (g) | 0.1 | 0.66 | 0.0 | 0.0 | 0.0 | 0.0 |
| Carbohydrates (g) | 5.3 | 34.98 | 5.7 | 37.62 | 6.3 | 41.58 |
| Sugar (g) | 3.6 | 23.76 | 2.8 | 18.48 | 4.9 | 32.34 |
| Protein (g) | 0.4 | 2.64 | 0.0 | 0.0 | 0.0 | 0.0 |
| Alcohol (%) | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 |
| Polyphenols (mg) | 30 | 198 | n/a | n/a | n/a | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kreimeyer, H.; Sydor, S.; Buchholz, L.; Toskal, C.; Özcürümez, M.; Schnabl, B.; Syn, W.-K.; Sowa, J.-P.; Manka, P.; Canbay, A. Non-Alcoholic Beer Influences Glucose and Lipid Metabolism and Changes Body Composition in Healthy, Young, Male Adults. Nutrients 2025, 17, 1625. https://doi.org/10.3390/nu17101625
Kreimeyer H, Sydor S, Buchholz L, Toskal C, Özcürümez M, Schnabl B, Syn W-K, Sowa J-P, Manka P, Canbay A. Non-Alcoholic Beer Influences Glucose and Lipid Metabolism and Changes Body Composition in Healthy, Young, Male Adults. Nutrients. 2025; 17(10):1625. https://doi.org/10.3390/nu17101625
Chicago/Turabian StyleKreimeyer, Henriette, Svenja Sydor, Lara Buchholz, Cagatay Toskal, Mustafa Özcürümez, Bernd Schnabl, Wing-Kin Syn, Jan-Peter Sowa, Paul Manka, and Ali Canbay. 2025. "Non-Alcoholic Beer Influences Glucose and Lipid Metabolism and Changes Body Composition in Healthy, Young, Male Adults" Nutrients 17, no. 10: 1625. https://doi.org/10.3390/nu17101625
APA StyleKreimeyer, H., Sydor, S., Buchholz, L., Toskal, C., Özcürümez, M., Schnabl, B., Syn, W.-K., Sowa, J.-P., Manka, P., & Canbay, A. (2025). Non-Alcoholic Beer Influences Glucose and Lipid Metabolism and Changes Body Composition in Healthy, Young, Male Adults. Nutrients, 17(10), 1625. https://doi.org/10.3390/nu17101625