
Oral Carnosine Supplementation Preserves Vascular Function of Sprague Dawley Rats on a High-Salt Diet via Restored Antioxidative Defence

 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Animals and Carnosine Supplementation Protocol
2.3. Blood Pressure Measurement and Blood Sampling
2.4. Assessment of Vasorelaxation of Aortic Rings
2.5. Aortic Tissue mRNA Expression Analyses
2.6. Advanced Oxidation Protein Products (AOPPs) and Ferric Reducing Antioxidant Power (FRAP) Assays
2.7. Superoxide Dismutase (SOD) Enzyme Activity Assay
2.8. Statistical Analysis
3. Results
3.1. Body Mass and Blood Pressure of Studied Groups
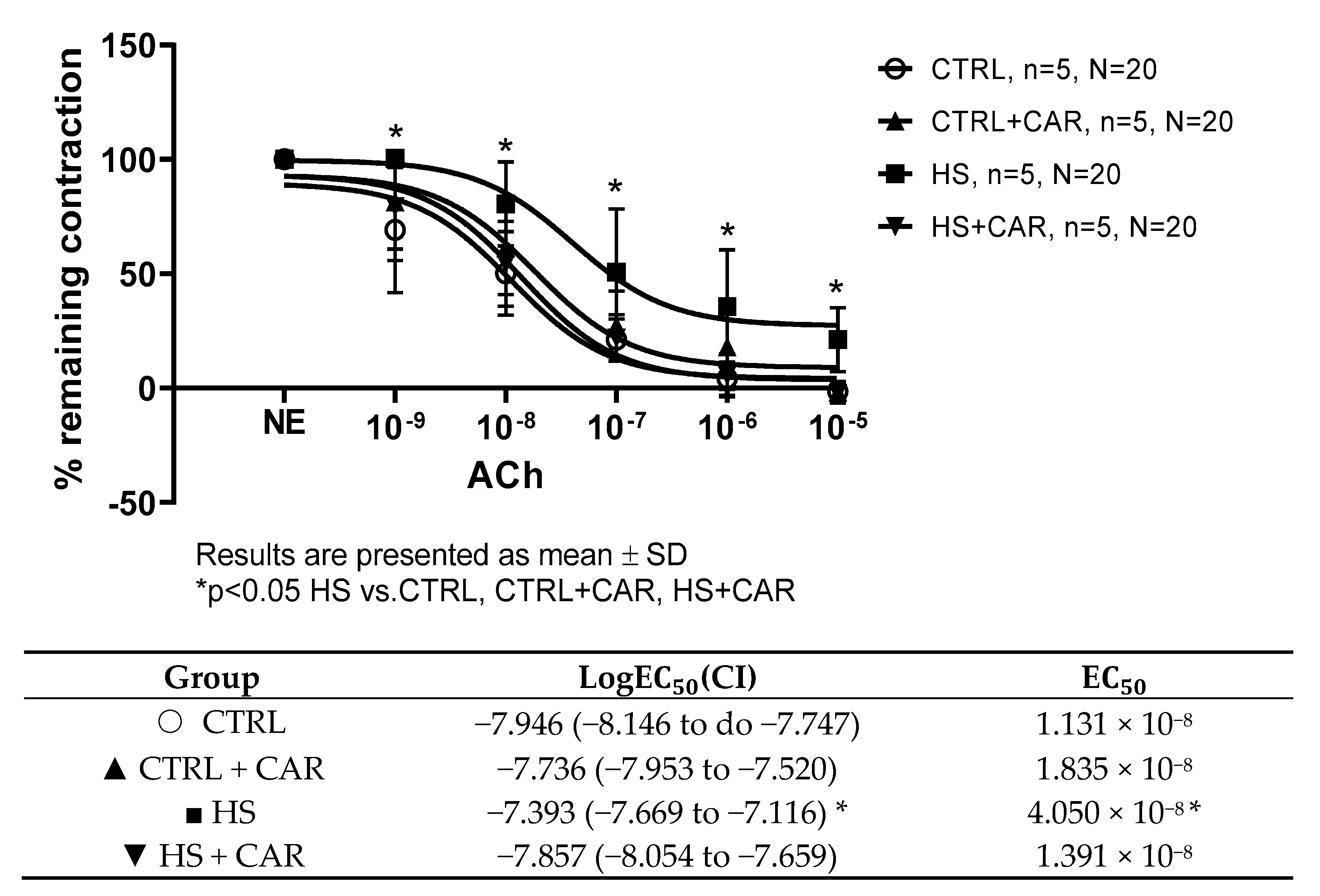
3.2. Effect of Carnosine Supplementation on Acetylcholine-Induced Relaxation (AChIR)
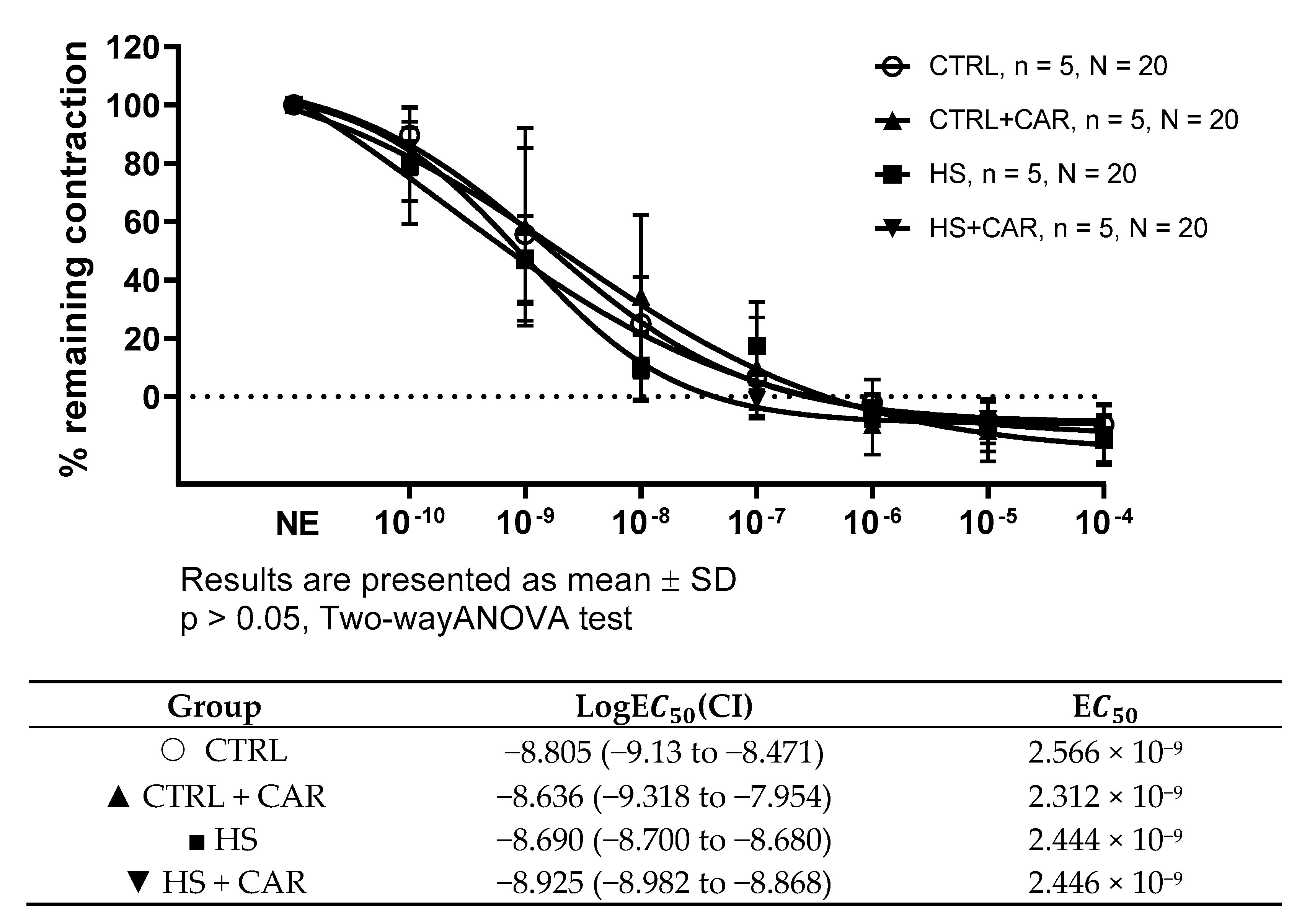
3.3. Effect of Carnosine Supplementation on Endothelium-Independent Relaxation Assessed by Sodium Nitroprusside (SNP)
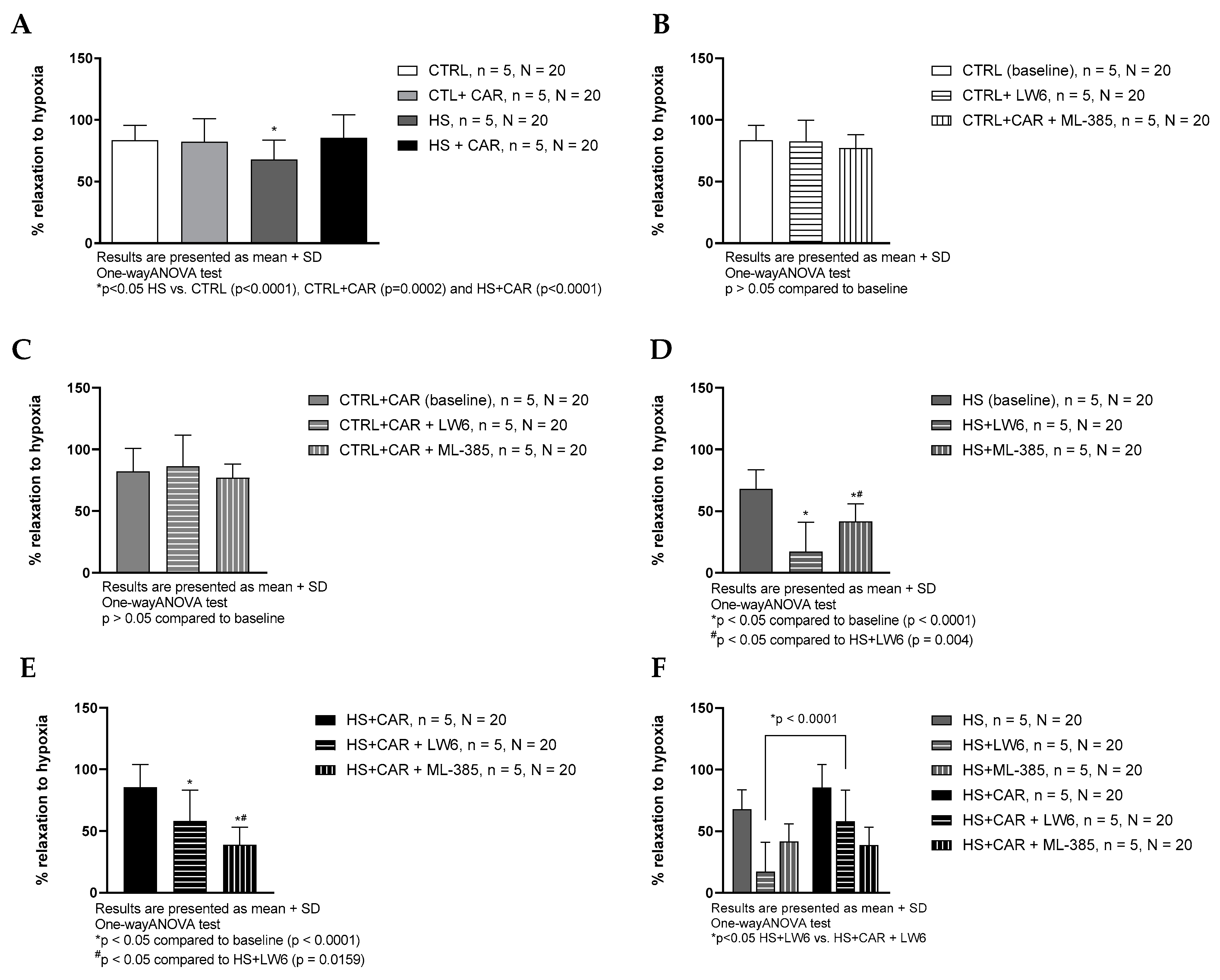
3.4. Effect of Carnosine Supplementation on Hypoxia-Induced Relaxation (HIR) of Aortic Rings
3.5. Effect of Carnosine Supplementation on Oxidative Stress, Antioxidative Capacity, and Superoxide Dismutase Activity
3.6. Effect of Carnosine Supplementation on Gene Expression in Aortic Tissue
3.6.1. Antioxidant Enzymes’ mRNA Expression in Aortic Tissue
3.6.2. Effect of Carnosine Supplementation on mRNA Expression of Enzymes Involved in Vascular Reactivity—Cyclooxygenases 1 and 2 (COX1 and 2) and Nitric Oxide Synthases (Inducible—iNOS, Endothelial—eNOS)
3.6.3. Effect of Carnosine Supplementation on mRNA Expression of HIF-1α, NRF2, and NQO1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boegehold, M.A. The Effect of High Salt Intake on Endothelial Function: Reduced Vascular Nitric Oxide in the Absence of Hypertension. J. Vasc. Res. 2013, 50, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Cosic, A.; Jukic, I.; Stupin, A.; Mihalj, M.; Mihaljevic, Z.; Novak, S.; Vukovic, R.; Drenjancevic, I. Attenuated Flow-induced Dilatation of Middle Cerebral Arteries Is Related to Increased Vascular Oxidative Stress in Rats on a Short-term High Salt Diet. J. Physiol. 2016, 594, 4917–4931. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, K.M.; Clifton, P.M.; Keogh, J.B. Endothelial Function Is Impaired after a High-Salt Meal in Healthy Subjects. Am. J. Clin. Nutr. 2011, 93, 500–505. [Google Scholar] [CrossRef]
- Liu, Y.; Fredricks, K.T.; Roman, R.J.; Lombard, J.H. Response of Resistance Arteries to Reduced PO2 and Vasodilators during Hypertension and Elevated Salt Intake. Am. J. Physiol. Heart Circ. Physiol. 1997, 273, H869–H877. [Google Scholar] [CrossRef]
- Matic, A.; Jukic, I.; Stupin, A.; Baric, L.; Mihaljevic, Z.; Unfirer, S.; Tartaro Bujak, I.; Mihaljevic, B.; Lombard, J.H.; Drenjancevic, I. High Salt Intake Shifts the Mechanisms of Flow-Induced Dilation in the Middle Cerebral Arteries of Sprague-Dawley Rats. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H718–H730. [Google Scholar] [CrossRef]
- Wu, G. Important Roles of Dietary Taurine, Creatine, Carnosine, Anserine and 4-Hydroxyproline in Human Nutrition and Health. Amino Acids 2020, 52, 329–360. [Google Scholar] [CrossRef]
- Bingül, İ.; Yılmaz, Z.; Aydın, A.F.; Çoban, J.; Doğru-Abbasoğlu, S.; Uysal, M. Antiglycation and Antioxidant Efficiency of Carnosine in the Plasma and Liver of Aged Rats. Geriatr. Gerontol. Int. 2017, 17, 2610–2614. [Google Scholar] [CrossRef]
- Jukić, I.; Kolobarić, N.; Stupin, A.; Matić, A.; Kozina, N.; Mihaljević, Z.; Mihalj, M.; Šušnjara, P.; Stupin, M.; Ćurić, Ž.B.; et al. Carnosine, Small but Mighty—Prospect of Use as Functional Ingredient for Functional Food Formulation. Antioxidants 2021, 10, 1037. [Google Scholar] [CrossRef]
- Caruso, G.; Privitera, A.; Antunes, B.M.; Lazzarino, G.; Lunte, S.M.; Aldini, G.; Caraci, F. The Therapeutic Potential of Carnosine as an Antidote against Drug-Induced Cardiotoxicity and Neurotoxicity: Focus on Nrf2 Pathway. Molecules 2022, 27, 4452. [Google Scholar] [CrossRef]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The Therapeutic Potential of Carnosine/Anserine Supplementation against Cognitive Decline: A Systematic Review with Meta-Analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef]
- Palin, M.-F.; Lapointe, J.; Gariépy, C.; Beaudry, D.; Kalbe, C. 57 Carnosine Prevents Oxidative Damage in Myoblast Cells Derived from Porcine Skeletal Muscle. J. Anim. Sci. 2019, 97, 59. [Google Scholar] [CrossRef]
- Ou, H.; Huang, Z.; Mo, Z.; Xiao, J. The Characteristics and Roles of Advanced Oxidation Protein Products in Atherosclerosis. Cardiovasc. Toxicol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Matic, A.; Jukic, I.; Mihaljevic, Z.; Kolobaric, N.; Stupin, A.; Kozina, N.; Bujak, I.T.; Kibel, A.; Lombard, J.H.; Drenjancevic, I. Low-Dose Angiotensin II Supplementation Restores Flow-Induced Dilation Mechanisms in Cerebral Arteries of Sprague-Dawley Rats on a High Salt Diet. J. Hypertens. 2022, 40, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Solana-Manrique, C.; Sanz, F.J.; Martínez-Carrión, G.; Paricio, N. Antioxidant and Neuroprotective Effects of Carnosine: Therapeutic Implications in Neurodegenerative Diseases. Antioxidants 2022, 11, 848. [Google Scholar] [CrossRef]
- Cesak, O.; Vostalova, J.; Vidlar, A.; Bastlova, P.; Student, V. Carnosine and Beta-Alanine Supplementation in Human Medicine: Narrative Review and Critical Assessment. Nutrients 2023, 15, 1770. [Google Scholar] [CrossRef]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’Amato, A.; Vistoli, G.; Carini, M. Understanding the Antioxidant and Carbonyl Sequestering Activity of Carnosine: Direct and Indirect Mechanisms. Free Radic. Res. 2021, 55, 321–330. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Li, X.; Gu, X.; Li, X.; Yan, J.; Mao, X.; Yu, Q.; Du, Y.; Kurihara, H.; Yan, C.; Li, W. Supplementation with Carnosine, a Food-derived Bioactive Dipeptide, Alleviates Dexamethasone-induced Oxidative Stress and Bone Impairment via the NRF2 Signaling Pathway. J. Sci. Food Agric. 2024, 105, 1091–1104. [Google Scholar] [CrossRef]
- Gong, X.; Gutala, R.; Jaiswal, A.K. Quinone Oxidoreductases and Vitamin K Metabolism. Vitam. Horm. 2008, 78, 85–101. [Google Scholar]
- Kohar, I.; Baca, M.; Suarna, C.; Stocker, R.; Southwell-Keely, P.T. Is α-Tocopherol a Reservoir for α-Tocopheryl Hydroquinone? Free Radic. Biol. Med. 1995, 19, 197–207. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct Evidence That Sulfhydryl Groups of Keap1 Are the Sensors Regulating Induction of Phase 2 Enzymes That Protect against Carcinogens and Oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed]
- Bondi, C.D.; Rush, B.M.; Hartman, H.L.; Wang, J.; Al-Bataineh, M.M.; Hughey, R.P.; Tan, R.J. Suppression of NRF2 Activity by HIF-1α Promotes Fibrosis after Ischemic Acute Kidney Injury. Antioxidants 2022, 11, 1810. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Stachurska, A.; Florczyk, U.; Rudnicka, D.; Jazwa, A.; Wegrzyn, J.; Kozakowska, M.; Stalinska, K.; Poellinger, L.; Levonen, A.; et al. HIF-1 Induction Attenuates Nrf2-Dependent IL-8 Expression in Human Endothelial Cells. Antioxid. Redox Signal 2009, 11, 1501–1517. [Google Scholar] [CrossRef] [PubMed]
- Lenda, D.M.; Sauls, B.A.; Boegehold, M.A. Reactive Oxygen Species May Contribute to Reduced Endothelium-Dependent Dilation in Rats Fed High Salt. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H7–H14. [Google Scholar] [CrossRef]
- Agrawal, A.; Rathor, R.; Kumar, R.; Singh, S.N.; Kumar, B.; Suryakumar, G. Endogenous Dipeptide-carnosine Supplementation Ameliorates Hypobaric Hypoxia-induced Skeletal Muscle Loss via Attenuating Endoplasmic Reticulum Stress Response and Maintaining Proteostasis. IUBMB Life 2022, 74, 101–116. [Google Scholar] [CrossRef]
- Drenjancevic, I.; Kibel, A.; Kibel, D.; Seric, V.; Cosic, A. Blood Pressure, Acid-Base and Blood Gas Status and Indicators of Oxidative Stress in Healthy Male Rats Exposed to Acute Hyperbaric Oxygenation. Undersea Hyperb. Med. 2013, 40, 319–328. [Google Scholar]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Rašić, L.; Jukić, I.; Drenjančević, I. Acute Hyperbaric Oxygenation, Contrary to Intermittent Hyperbaric Oxygenation, Adversely Affects Vasorelaxation in Healthy Sprague-Dawley Rats Due to Increased Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 7406027. [Google Scholar] [CrossRef]
- Kibel, A.; Novak, S.; Cosic, A.; Mihaljevic, Z.; Falck, J.R.; Drenjancevic, I. Hyperbaric Oxygenation Modulates Vascular Reactivity to Angiotensin-(1-7) in Diabetic Rats: Potential Role of Epoxyeicosatrienoic Acids. Diabetes Vasc. Dis. Res. 2015, 12, 33–45. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Frkanec, R.; Tavčar, B.; Kelava, V.; Bujak, I.T.; Kolobarić, N.; Kibel, A.; Drenjančević, I. Arachidonic Acid Metabolites of Cyp450 Enzymes and Hif-1α Modulate Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats under Acute and Intermittent Hyperbaric Oxygenation. Int. J. Mol. Sci. 2020, 21, 6353. [Google Scholar] [CrossRef]
- Kibel, A.; Cavka, A.; Cosic, A.; Falck, J.R.; Drenjancevic, I. Effects of Hyperbaric Oxygenation on Vascular Reactivity to Angiotensin II and Angiotensin-(1-7) in Rats. Undersea Hyperb. Med. 2012, 39, 1053–1066. [Google Scholar]
- Unfirer, S.; Mihalj, M.; Novak, S.; Kibel, A.; Cavka, A.; Mihaljevic, Z.; Gros, M.; Brizic, I.; Budimir, D.; Cosic, A.; et al. Hyperbaric Oxygenation Affects the Mechanisms of Acetylcholine-Induced Relaxation in Diabetic Rats. Undersea Hyperb. Med. 2016, 43, 787. [Google Scholar] [PubMed]
- Jukic, I.; Mihaljevic, Z.; Matic, A.; Mihalj, M.; Kozina, N.; Selthofer-Relatic, K.; Mihaljevic, D.; Koller, A.; Bujak, I.T.; Drenjancevic, I. Angiotensin II Type 1 Receptor Is Involved in Flow-Induced Vasomotor Responses of Isolated Middle Cerebral Arteries: Role of Oxidative Stress. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1609–H1624. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Flohé, L.; Ötting, F. [10] Superoxide Dismutase Assays. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 93–104. [Google Scholar]
- Olivera, S.; Graham, D. Sex differences in preclinical models of hypertension. J. Hum. Hypertens. 2023, 37, 619–625. [Google Scholar] [CrossRef]
- Zhu, J.; Mori, T.; Huang, T.; Lombard, J.H. Effect of High-Salt Diet on NO Release and Superoxide Production in Rat Aorta. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H575–H583. [Google Scholar] [CrossRef]
- Drenjancevic-Peric, I.; Lombard, J.H. Reduced Angiotensin II and Oxidative Stress Contribute to Impaired Vasodilation in Dahl Salt-Sensitive Rats on Low-Salt Diet. Hypertension 2005, 45, 687–691. [Google Scholar] [CrossRef]
- Zhu, J.; Drenjancevic-Peric, I.; McEwen, S.; Friesema, J.; Schulta, D.; Yu, M.; Roman, R.J.; Lombard, J.H. Role of Superoxide and Angiotensin II Suppression in Salt-Induced Changes in Endothelial Ca2+ Signaling and NO Production in Rat Aorta. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H929–H938. [Google Scholar] [CrossRef]
- Fredricks, K.T.; Liu, Y.; Rusch, N.J.; Lombard, J.H. Role of Endothelium and Arterial K+ Channels in Mediating Hypoxic Dilation of Middle Cerebral Arteries. Am. J. Physiol. Heart Circ. Physiol. 1994, 267, H580–H586. [Google Scholar] [CrossRef]
- Matrougui, K.; Loufrani, L.; Lévy, B.I.; Henrion, D. High NaCl Intake Decreases Both Flow-Induced Dilation and Pressure-Induced Myogenic Tone in Resistance Arteries from Normotensive Rats: Involvement of Cyclooxygenase-2. Pharmacol. Toxicol. 2001, 89, 183–187. [Google Scholar] [CrossRef]
- Raffai, G.; Wang, J.; Roman, R.J.; Anjaiah, S.; Weinberg, B.; Falck, J.R.; Lombard, J.H. Modulation by Cytochrome P450-4A ω-Hydroxylase Enzymes of Adrenergic Vasoconstriction and Response to Reduced PO2 in Mesenteric Resistance Arteries of Dahl Salt-Sensitive Rats. Microcirculation 2010, 17, 525–535. [Google Scholar] [CrossRef]
- Budzeń, S.; Rymaszewska, J. The Biological Role of Carnosine and Its Possible Applications in Medicine. Adv. Clin. Exp. Med. 2013, 22, 739–744. [Google Scholar] [PubMed]
- Prokopieva, V.D.; Yarygina, E.G.; Bokhan, N.A.; Ivanova, S.A. Use of Carnosine for Oxidative Stress Reduction in Different Pathologies. Oxid. Med. Cell. Longev. 2016, 2016, 2939087. [Google Scholar] [CrossRef] [PubMed]
- Evran, B.; Karpuzoğlu, H.; Develi, S.; Kalaz, E.B.; Soluk-Tekkeşin, M.; Olgaç, V.; Doğru-Abbasoğlu, S.; Uysal, M. Effects of Carnosine on Prooxidant–Antioxidant Status in Heart Tissue, Plasma and Erythrocytes of Rats with Isoproterenol-Induced Myocardial Infarction. Pharmacol. Rep. 2014, 66, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Kilis-Pstrusinska, K. Carnosine and Kidney Diseases: What We Currently Know? Curr. Med. Chem. 2020, 27, 1764–1781. [Google Scholar] [CrossRef]
- Selmeci, L. Advanced Oxidation Protein Products (AOPP): Novel Uremic Toxins, or Components of the Non-Enzymatic Antioxidant System of the Plasma Proteome? Free Radic. Res. 2011, 45, 1115–1123. [Google Scholar] [CrossRef]
- Liu, Z.; Yao, X.; Jiang, W.; Li, W.; Zhu, S.; Liao, C.; Zou, L.; Ding, R.; Chen, J. Advanced Oxidation Protein Products Induce Microglia-Mediated Neuroinflammation via MAPKs-NF-ΚB Signaling Pathway and Pyroptosis after Secondary Spinal Cord Injury. J. Neuroinflamm. 2020, 17, 90. [Google Scholar] [CrossRef]
- Gryszczyńska, B.; Formanowicz, D.; Budzyń, M.; Wanic-Kossowska, M.; Pawliczak, E.; Formanowicz, P.; Majewski, W.; Strzyżewski, K.W.; Kasprzak, M.P.; Iskra, M. Advanced Oxidation Protein Products and Carbonylated Proteins as Biomarkers of Oxidative Stress in Selected Atherosclerosis-Mediated Diseases. Biomed. Res. Int. 2017, 2017, 4975264. [Google Scholar] [CrossRef]
- Ozdogan, K.; Taskin, E.; Dursun, N. Protective Effect of Carnosine on Adriamycin-Induced Oxidative Heart Damage in Rats. Anadolu Kardiyol. Dergisi/Anatol. J. Cardiol. 2011, 11, 3–10. [Google Scholar] [CrossRef]
- Yan, S.; Wu, S.; Yin, M.; Chen, H.; Chen, H. Protective Effects from Carnosine and Histidine on Acetaminophen-Induced Liver Injury. J. Food Sci. 2009, 74, H259–H265. [Google Scholar] [CrossRef]
- Stefani, G.P.; Capalonga, L.; da Silva, L.R.; Heck, T.G.; Frizzo, M.N.; Sulzbacher, L.M.; Sulzbacher, M.M.; de Batista, D.; Vedovatto, S.; Bertoni, A.P.S.; et al. Effects of aerobic and resistance exercise training associated with carnosine precursor supplementation on maximal strength and in rats with heart failure. Life Sci. 2021, 282, 119816. [Google Scholar] [CrossRef]
- Suzuki, S.; Oguro, A.; Osada-Oka, M.; Funae, Y.; Imaoka, S. Epoxyeicosatrienoic Acids and/or Their Metabolites Promote Hypoxic Response of Cells. J. Pharmacol. Sci. 2008, 108, 79–88. [Google Scholar] [CrossRef]
- Manalo, D.J.; Rowan, A.; Lavoie, T.; Natarajan, L.; Kelly, B.D.; Ye, S.Q.; Garcia, J.G.N.; Semenza, G.L. Transcriptional Regulation of Vascular Endothelial Cell Responses to Hypoxia by HIF-1. Blood 2005, 105, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ming, J.; Li, T.; Yang, G.; Xu, J.; Chen, W.; Liu, L. Regulatory Effects of Hypoxia-Inducible Factor 1α on Vascular Reactivity and Its Mechanisms Following Hemorrhagic Shock in Rats. Shock. 2008, 30, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.; Claffey, K.P. Hypoxia-mediated Regulation of Gene Expression in Mammalian Cells. Int. J. Exp. Pathol. 1998, 79, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. HIF-1 and Mechanisms of Hypoxia Sensing. Curr. Opin. Cell Biol. 2001, 13, 167–171. [Google Scholar] [CrossRef]
- Schroedl, C.; McClintock, D.S.; Budinger, G.R.S.; Chandel, N.S. Hypoxic but Not Anoxic Stabilization of HIF-1α Requires Mitochondrial Reactive Oxygen Species. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 283, L922–L931. [Google Scholar] [CrossRef]
- Kolobarić, N.; Kozina, N.; Mihaljević, Z.; Drenjančević, I. Angiotensin II Exposure In Vitro Reduces High Salt-Induced Reactive Oxygen Species Production and Modulates Cell Adhesion Molecules’ Expression in Human Aortic Endothelial Cell Line. Biomedicines 2024, 12, 2741. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Gene | Nucleotide Sequence |
|---|---|
| SOD1 | forward: 5′-CTTCTGTCGTCTCCTTGCTTT-3′ |
| reverse: 5′-AGTGGTACAGCCTTGTGTATTG-3′ | |
| SOD2 | forward 5′-GCGACCTACGTGAACAATCT-3′ |
| reverse 5′-AGCGACCTTGCTCCTTATTG-3′ | |
| SOD3 | forward 5′-TGTTCTGCAACCTGCTACTG-3′ |
| reverse 5′-CATCCAGATCTCCAGGTCTTTG-3′ | |
| GPx1 | forward 5′-TCCAGTATGTGTGCTGCTCG-3′ |
| reverse 5′-GTGTCCGAACTGATTGCACG-3′ | |
| GPx4 | forward 5′-ATACGCCGAGTGTGGTTTAC-3′ |
| reverse 5′-GGCTGCAAACTCCTTGATTTC-3′ | |
| CAT | forward 5′-CTCAGGTGCGGACATTCTATAC-3′ |
| reverse 5′-CATTCTTAGGCTTCTGGGAGTT-3′ | |
| COX1 | forward 5′-TCCTGTTGCGAGCCCAGTT-3′ |
| reverse 5′-GCCAGTGATAGAGGTGGTTGAAT-3′ | |
| COX2 | forward 5′-GAAAGAAATGGCTGCAGAGTTGA-3′ |
| reverse 5′-GCAGGGCGGGATACAGTTC-3′ | |
| HIF-1 alpha | forward 5′-GCCCAGTGAGAAAGGGGA-3′ |
| reverse 5′-CGGCTGGTTACTGCTGGT-3′ | |
| NQO1 | forward 5′-AGTGGAAACCCACGAAGCCTACAA-3′ |
| reverse 5′-TGAACCAGTACAGCGGGAACTGAA-3′ | |
| NRF2 | forward 5′-AAGCCATTCACTCTCTGAACTTCT-3′ |
| reverse 5′-CTGGGACTTGTGTTTAGTGAAATG-3′ | |
| eNOS | forward 5′- CGAACAGCAGGAGCTAGAGG-3′ |
| reverse 5′- GAGGTGGATCTCTCCTGGT-3′ | |
| iNOS | forward 5′-TGGTGAGGGGACTGGACTTT-3′ |
| reverse 5′-CCAACTCTGCTGTTCTCCGT-3′ | |
| β-actin | forward 5′-TCTGTGTGGATTGGTGGCTCTA-3′ |
| reverse 5′-CTGCTTGCTGATCCACATCTG-3′ |
| Experimental Group | Body Mass [g] | SBP [mmHg] | DBP [mmHg] | MAP [mmHg] | |
|---|---|---|---|---|---|
| CTRL | Male (N = 7) | 348.11 ± 22.89 | 129 ± 4 | 92 ± 5 | 104 ± 4 |
| Female (N = 9) | 247.00 ± 16.61 * | 117 ± 10 * | 80 ± 6 | 93 ± 7 * | |
| Mean | 297.56 ± 19.75 | 123 ± 7 | 86 ± 6 | 99 ± 6 | |
| CTRL + CAR | Male (N = 9) | 360.56 ± 27.72 | 131 ± 6 | 95 ± 6 | 107 ± 5 |
| Female (N = 7) | 241.00 ± 12.04 * | 120 ± 3 * | 79 ± 8 | 93 ± 6 * | |
| Mean | 300.78 ± 19.88 | 126 ± 3 | 87 ± 7 | 100 ± 5 | |
| HS | Male (N = 11) | 349.10 ± 21.43 | 131 ± 9 | 90 ± 8 | 104 ± 8 |
| Female (N = 5) | 247.33 ± 25.33 * | 119 ± 2 * | 81 ± 10 | 94 ± 7 * | |
| Mean | 298.22 ± 23.38 | 125 ± 6 | 86 ± 9 | 99 ± 7 | |
| HS + CAR | Male (N = 10) | 356.50 ± 37.52 | 129 ± 5 | 88 ± 12 | 101 ± 9 |
| Female (N = 6) | 234.70 ± 12.07 * | 120 ± 2 * | 84 ± 6 | 96 ± 4 | |
| Mean | 295.60 ± 24.79 | 125 ± 4 | 86 ± 9 | 99 ± 6 |
| CTRL | CTRL + CAR | HS | HS + CAR | |
|---|---|---|---|---|
| AOPPs [nmol/mL] | 15.91 ± 7.04 ‡ | 14.97 ± 5.67 ‡ | 44.26 ± 12.96 *† | 11.43 ± 3.09 ‡ |
| FRAP [mM Trolox] | 0.08 ± 0.02 ‡ | 0.11 ± 0.01 ‡ | 0.03 ± 0.02 *† | 0.07 ± 0.03 ‡ |
| SOD [U/mgP] | 12.39 ± 1.31 | 12.81 ± 0.76 | 11.49 ± 1.06 | 17.72 ± 2.21 *†‡ |
| CTRL | CTRL + CAR | HS | HS + CAR | |
|---|---|---|---|---|
| SOD1 | 0.064 ± 0.03 | 0.181 ± 0.07 * | 0.074 ± 0.03 † | 0.262 ± 0.09 *‡ |
| SOD2 | 0.062 ± 0.02 | 0.162 ± 0.08 * | 0.085 ± 0.06 † | 0.226 ± 0.11 *‡ |
| SOD3 | 3.140 ± 2.27 | 2.351 ± 1.32 | 2.174 ± 1.78 | 1.499 ± 0.64 |
| CAT | 1.355 ± 0.28 | 1.593 ± 0.34 | 1.730 ± 0.49 | 1.966 ± 1.17 |
| GPx1 | 0.415 ± 0.38 | 0.713 ± 0.13 | 0.621 ± 0.20 | 1.063 ± 0.44 *‡ |
| GPx4 | 0.762 ± 0.61 | 0.974 ± 0.25 | 0.681 ± 0.16 | 0.981 ± 0.26 ‡ |
| CTRL | CTRL + CAR | HS | HS + CAR | |
|---|---|---|---|---|
| COX1 | 4.107 ± 3.00 | 3.997 ± 2.47 | 4.185 ± 2.62 | 4.322 ± 1.61 |
| COX2 | 2.468 ± 0.77 | 2.828 ± 0.48 | 1.954 ± 0.86 | 2.208 ± 0.93 |
| iNOS | 0.273 ± 0.28 | 0.569 ± 0.19 | 0.291 ± 0.16 | 0.352 ± 0.20 |
| eNOS | 0.119 ± 0.16 | 0.278 ± 0.33 | 0.113 ± 0.12 | 0.582 ± 0.27 *‡ |
| CTRL | CTRL + CAR | HS | HS + CAR | |
|---|---|---|---|---|
| HIF-1α | 1.412 ± 0.75 | 1.576 ± 0.44 | 1.082 ± 0.33 | 1.324 ± 0.57 |
| NRF2 | 0.046 ± 0.06 | 0.498 ± 0.37 * | 0.149 ± 0.08 | 0.536 ± 0.22 *‡ |
| NQO1 | 0.275 ± 0.15 | 0.925 ± 0.43 * | 0.571 ± 0.29 | 2.064 ± 1.53 *‡ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drenjančević, I.; Stupin, A.; Jukić, I.; Kolobarić, N.; Šušnjara, P.; Kozina, N.; Kovač, L.; Mihaljević, Z. Oral Carnosine Supplementation Preserves Vascular Function of Sprague Dawley Rats on a High-Salt Diet via Restored Antioxidative Defence. Nutrients 2025, 17, 36. https://doi.org/10.3390/nu17010036
Drenjančević I, Stupin A, Jukić I, Kolobarić N, Šušnjara P, Kozina N, Kovač L, Mihaljević Z. Oral Carnosine Supplementation Preserves Vascular Function of Sprague Dawley Rats on a High-Salt Diet via Restored Antioxidative Defence. Nutrients. 2025; 17(1):36. https://doi.org/10.3390/nu17010036
Chicago/Turabian StyleDrenjančević, Ines, Ana Stupin, Ivana Jukić, Nikolina Kolobarić, Petar Šušnjara, Nataša Kozina, Lora Kovač, and Zrinka Mihaljević. 2025. "Oral Carnosine Supplementation Preserves Vascular Function of Sprague Dawley Rats on a High-Salt Diet via Restored Antioxidative Defence" Nutrients 17, no. 1: 36. https://doi.org/10.3390/nu17010036
APA StyleDrenjančević, I., Stupin, A., Jukić, I., Kolobarić, N., Šušnjara, P., Kozina, N., Kovač, L., & Mihaljević, Z. (2025). Oral Carnosine Supplementation Preserves Vascular Function of Sprague Dawley Rats on a High-Salt Diet via Restored Antioxidative Defence. Nutrients, 17(1), 36. https://doi.org/10.3390/nu17010036